
Characterization of a Near Full-Length Hepatitis E Virus Genome of Subtype 3c Generated from Naturally Infected South African Backyard Pigs


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Study Design
2.3. Sample Collection and Processing
2.4. Metagenomic Sequencing
2.5. Genome Assembly and Annotation
2.6. Phylogenetic Analysis and ‘HEVnet Typing Tool’ Analysis
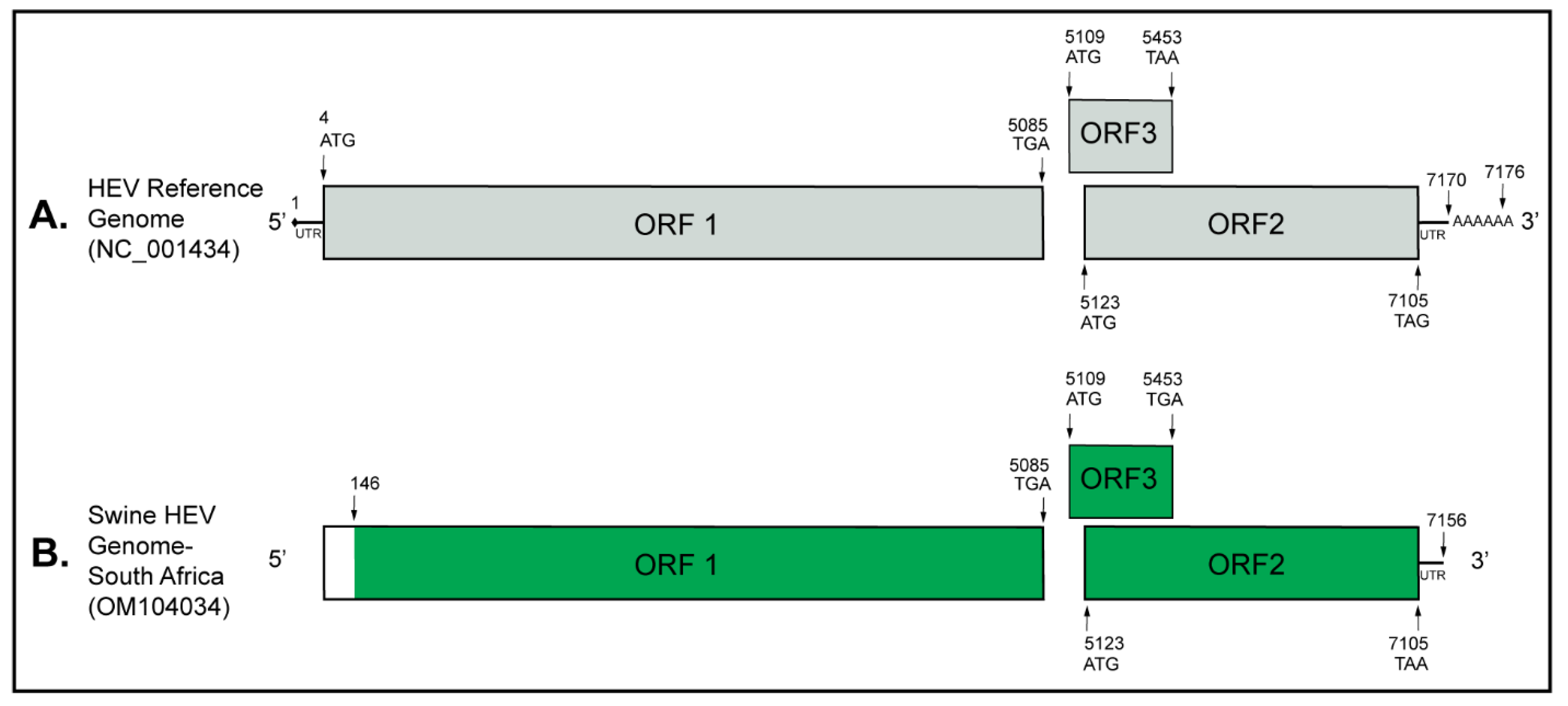
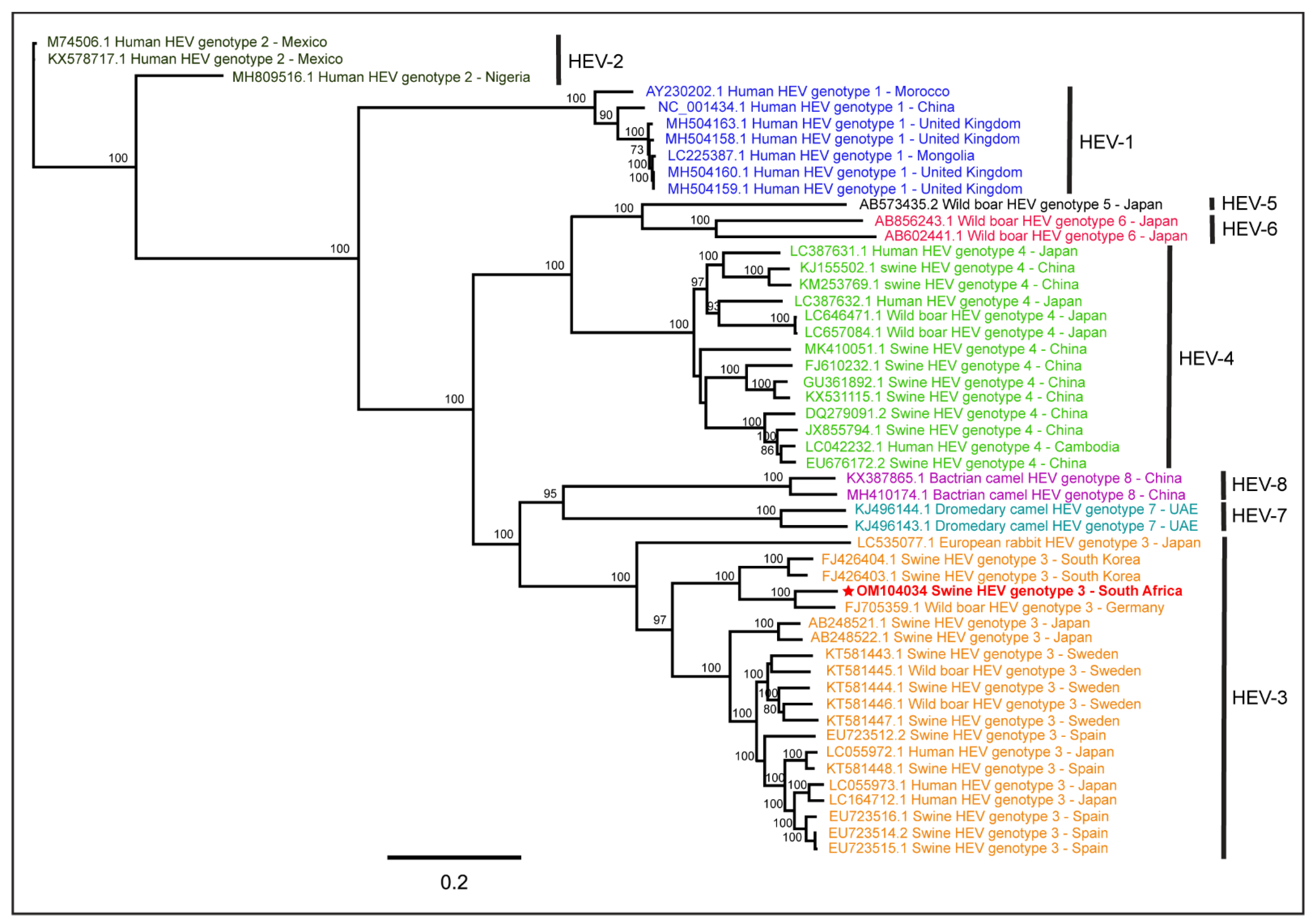
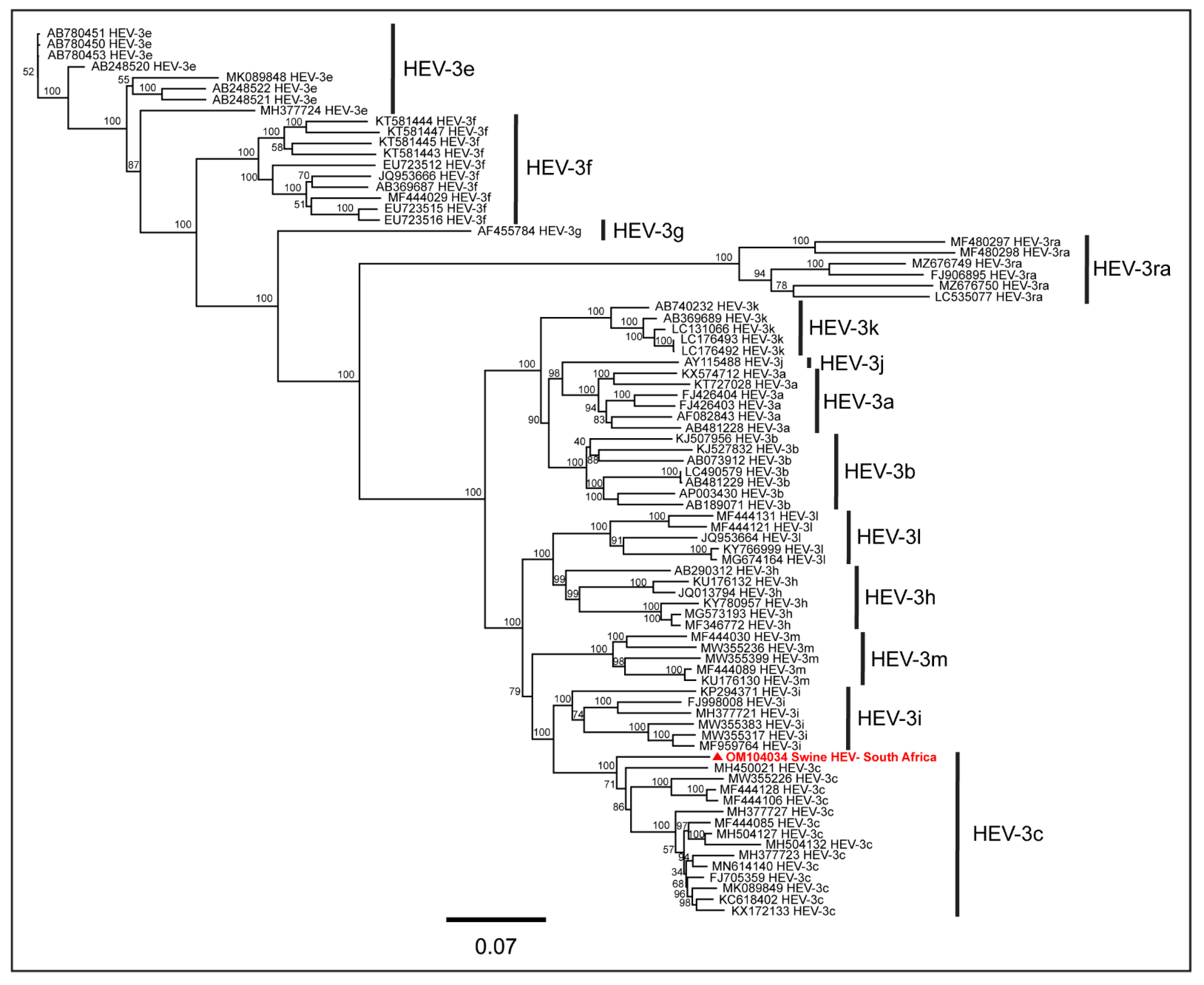
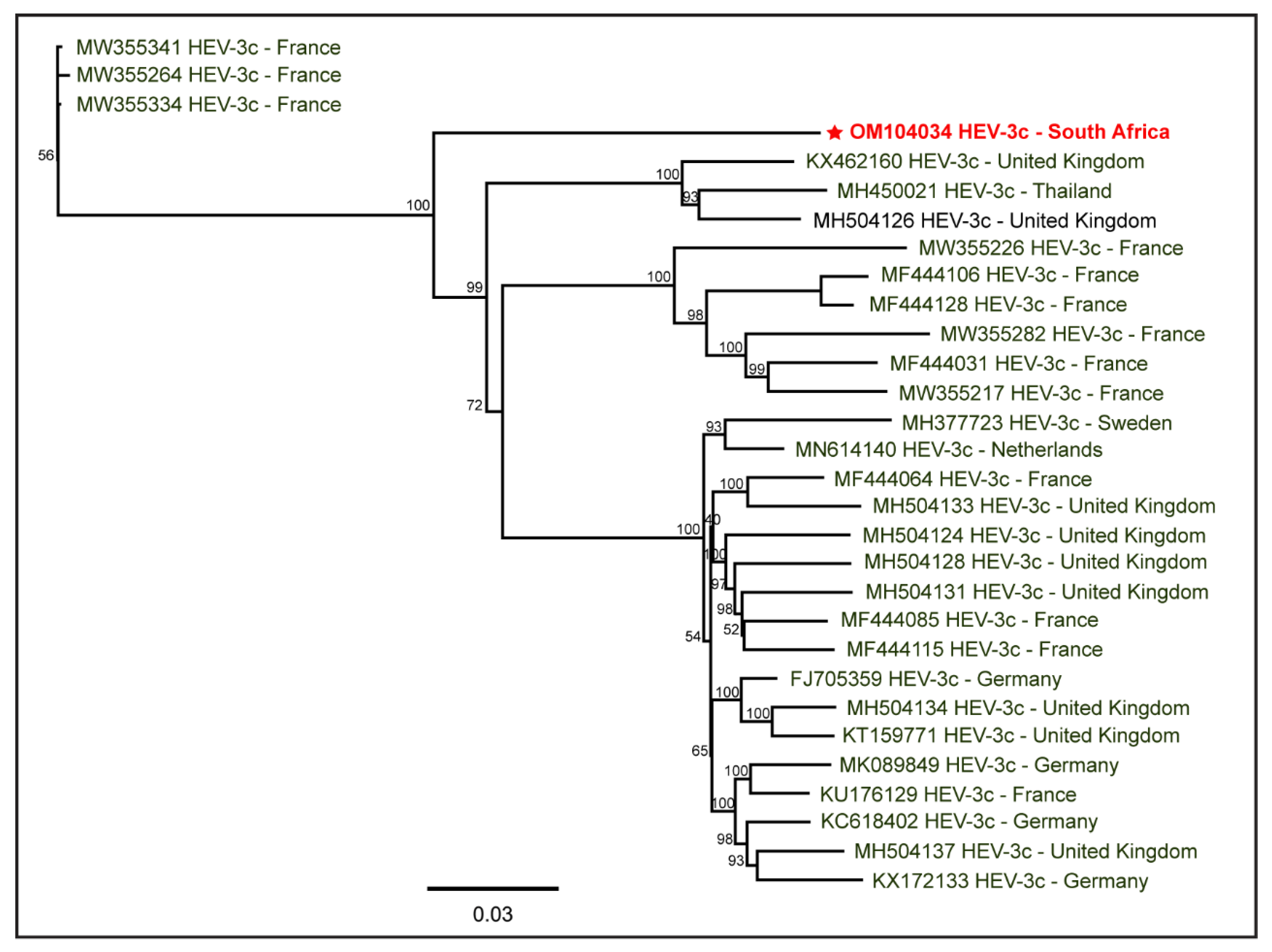
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Henritzi, D.; Petric, P.P.; Lewis, N.S.; Graaf, A.; Pessia, A.; Starick, E.; Breithaupt, A.; Strebelow, G.; Luttermann, C.; Parker, L.M.K.; et al. Surveillance of European Domestic Pig Populations Identifies an Emerging Reservoir of Potentially Zoonotic Swine Influenza A Viruses. Cell Host Microbe 2020, 28, 614–627.e616. [Google Scholar] [CrossRef]
- Chauhan, R.P.; Gordon, M.L. A Systematic Review Analyzing the Prevalence and Circulation of Influenza Viruses in Swine Population Worldwide. Pathogens 2020, 9, 355. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, R.P.; Gordon, M.L. Deciphering transmission dynamics and spillover of avian influenza viruses from avian species to swine populations globally. Virus Genes 2021, 57, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, R.P.; Gordon, M.L. A systematic review of influenza A virus prevalence and transmission dynamics in backyard swine populations globally. Porc. Health Manag. 2022, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, E.; Salogni, C.; Martella, V.; Alborali, G.L.; Scaburri, A.; Boniotti, M.B. Assessing the Epidemiology of Rotavirus A, B, C and H in Diarrheic Pigs of Different Ages in Northern Italy. Pathogens 2022, 11, 467. [Google Scholar] [CrossRef]
- Chauhan, R.P.; Gordon, M.L. Review of genome sequencing technologies in molecular characterization of influenza A viruses in swine. J. Veter. Diagn. Investig. 2022, 34, 177–189. [Google Scholar] [CrossRef]
- Chauhan, R.P.; Gordon, M.L. An overview of influenza A virus genes, protein functions, and replication cycle highlighting important updates. Virus Genes 2022, 58, 255–269. [Google Scholar] [CrossRef]
- Pallerla, S.R.; Harms, D.; Johne, R.; Todt, D.; Steinmann, E.; Schemmerer, M.; Wenzel, J.J.; Hofmann, J.; Shih, J.W.K.; Wedemeyer, H.; et al. Hepatitis E Virus Infection: Circulation, Molecular Epidemiology, and Impact on Global Health. Pathogens 2020, 9, 856. [Google Scholar] [CrossRef]
- Meester, M.; Tobias, T.J.; Bouwknegt, M.; Kusters, N.E.; Stegeman, J.A.; van der Poel, W.H.M. Infection dynamics and persistence of hepatitis E virus on pig farms—A review. Porc. Health Manag. 2021, 7, 16. [Google Scholar] [CrossRef]
- Pavio, N.; Doceul, V.; Bagdassarian, E.; Johne, R. Recent knowledge on hepatitis E virus in Suidae reservoirs and transmission routes to human. Veter. Res. 2017, 48, 78. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Ge, S.; Zhang, J.; Guo, Q.; Ng, M.H.; Wang, F.; Xia, N.; Jiang, Q. Swine as a Principal Reservoir of Hepatitis E Virus That Infects Humans in Eastern China. J. Infect. Dis. 2006, 193, 1643–1649. [Google Scholar] [CrossRef] [PubMed]
- Doceul, V.; Bagdassarian, E.; Demange, A.; Pavio, N. Zoonotic Hepatitis E Virus: Classification, Animal Reservoirs and Transmission Routes. Viruses 2016, 8, 270. [Google Scholar] [CrossRef] [PubMed]
- Corman, V.M.; Nagy, P.; Ostermann, S.; Arloth, J.; Liljander, A.; Barua, R.; Das Gupta, A.; Hakimuddin, F.; Juhasz, J.; Wernery, U.; et al. Hepatitis E Virus Genotype 7 RNA and Antibody Kinetics in Naturally Infected Dromedary Calves, United Arab Emirates. Emerg. Infect. Dis. 2020, 26, 2214–2217. [Google Scholar] [CrossRef] [PubMed]
- Dalton, H.R.; Izopet, J. Transmission and Epidemiology of Hepatitis E Virus Genotype 3 and 4 Infections. Cold Spring Harb. Perspect. Med. 2018, 8, a032144. [Google Scholar] [CrossRef]
- Ivanova, A.; Tefanova, V.; Reshetnjak, I.; Kuznetsova, T.; Geller, J.; Lundkvist, Å.; Janson, M.; Neare, K.; Velström, K.; Jokelainen, P.; et al. Hepatitis E Virus in Domestic Pigs, Wild Boars, Pig Farm Workers, and Hunters in Estonia. Food Environ. Virol. 2015, 7, 403–412. [Google Scholar] [CrossRef]
- Meng, X.J.; Purcell, R.H.; Halbur, P.G.; Lehman, J.R.; Webb, D.M.; Tsareva, T.S.; Haynes, J.S.; Thacker, B.J.; Emerson, S.U. A novel virus in swine is closely related to the human hepatitis E virus. Proc. Natl. Acad. Sci. USA 1997, 94, 9860–9865. [Google Scholar] [CrossRef]
- Sooryanarain, H.; Meng, X.-J. Swine hepatitis E virus: Cross-species infection, pork safety and chronic infection. Virus Res. 2020, 284, 197985. [Google Scholar] [CrossRef]
- Bouwknegt, M.; Frankena, K.; Rutjes, S.A.; Wellenberg, G.J.; de Roda Husman, A.M.; van der Poel, W.H.; de Jong, M.C. Estimation of hepatitis E virus transmission among pigs due to contact-exposure. Vet. Res. 2008, 39, 40. [Google Scholar] [CrossRef]
- Bouwknegt, M.; Rutjes, S.A.; Reusken, C.B.; Stockhofe-Zurwieden, N.; Frankena, K.; de Jong, M.C.; de Roda Husman, A.M.; Poel, W.H. The course of hepatitis E virus infection in pigs after contact-infection and intravenous inoculation. BMC Vet. Res. 2009, 5, 7. [Google Scholar] [CrossRef]
- Casas, M.; Pina, S.; de Deus, N.; Peralta, B.; Martín, M.; Segalés, J. Pigs orally inoculated with swine hepatitis E virus are able to infect contact sentinels. Vet. Microbiol. 2009, 138, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Treagus, S.; Wright, C.; Baker-Austin, C.; Longdon, B.; Lowther, J. The Foodborne Transmission of Hepatitis E Virus to Humans. Food Environ. Virol. 2021, 13, 127–145. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Liu, J.; Li, Y.; Su, J.; Ma, Z.; Bramer, W.M.; Cao, W.; de Man, R.A.; Peppelenbosch, M.P.; Pan, Q. The global epidemiology of hepatitis E virus infection: A systematic review and meta-analysis. Liver Int. 2020, 40, 1516–1528. [Google Scholar] [CrossRef] [PubMed]
- Micas, F.; Suin, V.; Péron, J.M.; Scholtes, C.; Tuaillon, E.; Vanwolleghem, T.; Bocket, L.; Lhomme, S.; Dimeglio, C.; Izopet, J.; et al. Analyses of Clinical and Biological Data for French and Belgian Immunocompetent Patients Infected With Hepatitis E Virus Genotypes 4 and 3. Front. Microbiol. 2021, 12, 645020. [Google Scholar] [CrossRef]
- Owada, Y.; Oshiro, Y.; Inagaki, Y.; Harada, H.; Fujiyama, N.; Kawagishi, N.; Yagisawa, T.; Usui, J.; Akutsu, N.; Itabashi, Y.; et al. A Nationwide Survey of Hepatitis E Virus Infection and Chronic Hepatitis in Heart and Kidney Transplant Recipients in Japan. Transplantation 2020, 104, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.J.; Cheung, R.; Gish, R.G.; Chitnis, A.S. Prevalence of hepatitis E infection among adults with concurrent chronic liver disease. J. Viral. Hepat. 2021, 28, 1643–1655. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.J.; Zhou, N.; Dong, W.; Li, G.X.; Cong, W.; Zhu, X.Q. Seroprevalence and risk factors of hepatitis E virus infection in cancer patients in eastern China. Int. J. Infect. Dis. 2018, 71, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Ghandili, S.; Lindhauer, C.; Pischke, S.; Zur Wiesch, J.S.; Von Kroge, P.H.; Polywka, S.; Bokemeyer, C.; Fiedler, W.; Kröger, N.; Ayuk, F.; et al. Clinical features of hepatitis E infections in patients with hematological disorders. Haematologica 2022. [Google Scholar] [CrossRef]
- Fu, H.; Li, L.; Zhu, Y.; Wang, L.; Geng, J.; Chang, Y.; Xue, C.; Du, G.; Li, Y.; Zhuang, H. Hepatitis E virus infection among animals and humans in Xinjiang, China: Possibility of swine to human transmission of sporadic hepatitis E in an endemic area. Am. J. Trop. Med. Hyg. 2010, 82, 961–966. [Google Scholar] [CrossRef]
- Salines, M.; Andraud, M.; Rose, N. From the epidemiology of hepatitis E virus (HEV) within the swine reservoir to public health risk mitigation strategies: A comprehensive review. Veter. Res. 2017, 48, 31. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.S.; Silveira, S.; Caron, V.S.; Mósena, A.C.S.; Weber, M.N.; Cibulski, S.P.; Medeiros, A.A.R.; Silva, G.S.; Corbellini, L.G.; Klein, R.; et al. Backyard pigs are a reservoir of zoonotic hepatitis E virus in southern Brazil. Trans. R Soc. Trop. Med. Hyg. 2018, 112, 14–21. [Google Scholar] [CrossRef]
- Passos-Castilho, A.M.; Granato, C.F.H. High frequency of hepatitis E virus infection in swine from South Brazil and close similarity to human HEV isolates. Braz. J. Microbiol. 2017, 48, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Ji, Y.; Li, Y.; Ma, Z.; Pan, Q. Estimating the global prevalence of hepatitis E virus in swine and pork products. One Health 2022, 14, 100362. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, J.J.; Preiss, J.; Schemmerer, M.; Huber, B.; Plentz, A.; Jilg, W. Detection of hepatitis E virus (HEV) from porcine livers in Southeastern Germany and high sequence homology to human HEV isolates. J. Clin. Virol. 2011, 52, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Feagins, A.R.; Opriessnig, T.; Guenette, D.K.; Halbur, P.G.; Meng, X.J. Detection and characterization of infectious Hepatitis E virus from commercial pig livers sold in local grocery stores in the USA. J. Gen. Virol. 2007, 88, 912–917. [Google Scholar] [CrossRef] [PubMed]
- Yazaki, Y.; Mizuo, H.; Takahashi, M.; Nishizawa, T.; Sasaki, N.; Gotanda, Y.; Okamoto, H. Sporadic acute or fulminant hepatitis E in Hokkaido, Japan, may be food-borne, as suggested by the presence of hepatitis E virus in pig liver as food. J. Gen. Virol. 2003, 84, 2351–2357. [Google Scholar] [CrossRef]
- Pavio, N.; Merbah, T.; Thébault, A. Frequent hepatitis E virus contamination in food containing raw pork liver, France. Emerg. Infect. Dis. 2014, 20, 1925–1927. [Google Scholar] [CrossRef]
- Temmam, S.; Besnard, L.; Andriamandimby, S.F.; Foray, C.; Rasamoelina-Andriamanivo, H.; Héraud, J.M.; Cardinale, E.; Dellagi, K.; Pavio, N.; Pascalis, H.; et al. High prevalence of hepatitis E in humans and pigs and evidence of genotype-3 virus in swine, Madagascar. Am. J. Trop. Med. Hyg. 2013, 88, 329–338. [Google Scholar] [CrossRef]
- Okano, H.; Takahashi, M.; Isono, Y.; Tanaka, H.; Nakano, T.; Oya, Y.; Sugimoto, K.; Ito, K.; Ohmori, S.; Maegawa, T.; et al. Characterization of sporadic acute hepatitis E and comparison of hepatitis E virus genomes in acute hepatitis patients and pig liver sold as food in Mie, Japan. Hepatol. Res. 2014, 44, E63–E76. [Google Scholar] [CrossRef]
- Chelli, E.; Suffredini, E.; De Santis, P.; De Medici, D.; Di Bella, S.; D’Amato, S.; Gucciardi, F.; Guercio, A.; Ostanello, F.; Perrone, V.; et al. Hepatitis E Virus Occurrence in Pigs Slaughtered in Italy. Animals 2021, 11, 277. [Google Scholar] [CrossRef]
- Mykytczuk, O.; Harlow, J.; Bidawid, S.; Corneau, N.; Nasheri, N. Prevalence and Molecular Characterization of the Hepatitis E Virus in Retail Pork Products Marketed in Canada. Food Environ. Virol. 2017, 9, 208–218. [Google Scholar] [CrossRef] [Green Version]
- Pallerla, S.R.; Schembecker, S.; Meyer, C.G.; Linh, L.T.K.; Johne, R.; Wedemeyer, H.; Bock, C.T.; Kremsner, P.G.; Velavan, T.P. Hepatitis E virus genome detection in commercial pork livers and pork meat products in Germany. J. Viral Hepat. 2021, 28, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Cao, W.; Situ, J.; Zhao, Y.; Huang, F.; Yu, W. Hepatitis E Virus Detected in Pork Products. Food Environ. Virol. 2018, 10, 391–393. [Google Scholar] [CrossRef] [PubMed]
- Moor, D.; Liniger, M.; Baumgartner, A.; Felleisen, R. Screening of Ready-to-Eat Meat Products for Hepatitis E Virus in Switzerland. Food Environ. Virol. 2018, 10, 263–271. [Google Scholar] [CrossRef]
- Bouwknegt, M.; Lodder-Verschoor, F.; van der Poel, W.H.; Rutjes, S.A.; de Roda Husman, A.M. Hepatitis E virus RNA in commercial porcine livers in The Netherlands. J. Food Prot. 2007, 70, 2889–2895. [Google Scholar] [CrossRef]
- Colson, P.; Borentain, P.; Queyriaux, B.; Kaba, M.; Moal, V.; Gallian, P.; Heyries, L.; Raoult, D.; Gerolami, R. Pig liver sausage as a source of hepatitis E virus transmission to humans. J. Infect. Dis. 2010, 202, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Hoan, N.X.; Huy, P.X.; Sy, B.T.; Meyer, C.G.; Son, T.V.; Binh, M.T.; Giang, D.P.; Tu Anh, D.; Bock, C.T.; Wang, B.; et al. High Hepatitis E virus (HEV) Positivity Among Domestic Pigs and Risk of HEV Infection of Individuals Occupationally Exposed to Pigs and Pork Meat in Hanoi, Vietnam. Open Forum Infect. Dis. 2019, 6, ofz306. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Wang, L.; Wang, X.; Fu, H.; Bu, Q.; Liu, P.; Zhu, Y.; Wang, M.; Sui, Y.; Zhuang, H. Potential risk of zoonotic transmission from young swine to human: Seroepidemiological and genetic characterization of hepatitis E virus in human and various animals in Beijing, China. J. Viral Hepat. 2011, 18, e583–e590. [Google Scholar] [CrossRef] [PubMed]
- Drobeniuc, J.; Favorov, M.O.; Shapiro, C.N.; Bell, B.P.; Mast, E.E.; Dadu, A.; Culver, D.; Iarovoi, P.; Robertson, B.H.; Margolis, H.S. Hepatitis E virus antibody prevalence among persons who work with swine. J. Infect. Dis. 2001, 184, 1594–1597. [Google Scholar] [CrossRef]
- Shirazi, R.; Pozzi, P.; Wax, M.; Bar-Or, I.; Asulin, E.; Lustig, Y.; Mendelson, E.; Ben-Ari, Z.; Schwartz, E.; Mor, O. Hepatitis E in pigs in Israel: Seroprevalence, molecular characterisation and potential impact on humans. Euro Surveill. 2018, 23, 1800067. [Google Scholar] [CrossRef]
- Bansal, M.; Kaur, S.; Deka, D.; Singh, R.; Gill, J.P.S. Seroepidemiology and molecular characterization of hepatitis E virus infection in swine and occupationally exposed workers in Punjab, India. Zoonoses Public Health 2017, 64, 662–672. [Google Scholar] [CrossRef]
- Chaussade, H.; Rigaud, E.; Allix, A.; Carpentier, A.; Touzé, A.; Delzescaux, D.; Choutet, P.; Garcia-Bonnet, N.; Coursaget, P. Hepatitis E virus seroprevalence and risk factors for individuals in working contact with animals. J. Clin. Virol. 2013, 58, 504–508. [Google Scholar] [CrossRef] [PubMed]
- Bagulo, H.; Majekodunmi, A.O.; Welburn, S.C.; Bimi, L. Hepatitis E seroprevalence and risk factors in humans and pig in Ghana. BMC Infect. Dis. 2022, 22, 132. [Google Scholar] [CrossRef] [PubMed]
- Modiyinji, A.F.; Sanding, G.M.A.M.; Atsama, M.A.; Monamele, C.G.; Nola, M.; Njouom, R. Serological and molecular investigation of hepatitis E virus in pigs reservoirs from Cameroon reveals elevated seroprevalence and presence of genotype 3. PLoS ONE 2020, 15, e0229073. [Google Scholar] [CrossRef] [PubMed]
- Owolodun, O.A.; Gerber, P.F.; Giménez-Lirola, L.G.; Kwaga, J.K.; Opriessnig, T. First report of hepatitis E virus circulation in domestic pigs in Nigeria. Am. J. Trop. Med. Hyg. 2014, 91, 699–704. [Google Scholar] [CrossRef]
- Adelabu, O.A.; Chuks Iweriebor, B.; Nwodo, U.U.; Obi, L.C.; Okoh, A.I. Incidence and Molecular Characterization of Hepatitis E Virus from Swine in Eastern Cape, South Africa. Adv. Virol. 2017, 2017, 1073253. [Google Scholar] [CrossRef]
- Chauhan, R.P.; San, J.E.; Gordon, M.L. Metagenomic Analysis of RNA Fraction Reveals the Diversity of Swine Oral Virome on South African Backyard Swine Farms in the uMgungundlovu District of KwaZulu-Natal Province. Pathogens 2022, 11, 927. [Google Scholar] [CrossRef]
- Chauhan, R.P.; Gordon, M.L. No evidence of influenza A virus RNA in South African backyard swine in the uMgungundlovu district of the KwaZulu-Natal province. Indian J. Animal Res. 2022. [Google Scholar] [CrossRef]
- Prickett, J.R.; Kim, W.; Simer, R. Oral-fluid samples for surveillance of commercial growing pigs for porcine reproductive and respiratory syndrome virus and porcine circovirus type 2 infections. JSHAP 2008, 16, 86–91. [Google Scholar]
- Henao-Diaz, A.; Giménez-Lirola, L.; Baum, D.H.; Zimmerman, J. Guidelines for oral fluid-based surveillance of viral pathogens in swine. Porc. Health Manag. 2020, 6, 28. [Google Scholar] [CrossRef]
- Vilsker, M.; Moosa, Y.; Nooij, S.; Fonseca, V.; Ghysens, Y.; Dumon, K.; Pauwels, R.; Alcantara, L.C.; Vanden Eynden, E.; Vandamme, A.-M.; et al. Genome Detective: An automated system for virus identification from high-throughput sequencing data. Bioinformatics 2019, 35, 871–873. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Mulder, A.C.; Kroneman, A.; Franz, E.; Vennema, H.; Tulen, A.D.; Takkinen, J.; Hofhuis, A.; Adlhoch, C. HEVnet: A One Health, collaborative, interdisciplinary network and sequence data repository for enhanced hepatitis E virus molecular typing, characterisation and epidemiological investigations. Euro Surveill. 2019, 24, 1800407. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.J.; Norder, H.; Okamoto, H.; van der Poel, W.H.M.; Reuter, G.; et al. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Baha, S.; Behloul, N.; Liu, Z.; Wei, W.; Shi, R.; Meng, J. Comprehensive analysis of genetic and evolutionary features of the hepatitis E virus. BMC Genom. 2019, 20, 790. [Google Scholar] [CrossRef]
- Zhou, J.H.; Li, X.R.; Lan, X.; Han, S.Y.; Wang, Y.N.; Hu, Y.; Pan, Q. The genetic divergences of codon usage shed new lights on transmission of hepatitis E virus from swine to human. Infect. Genet. Evol. 2019, 68, 23–29. [Google Scholar] [CrossRef]
- Pierini, I.; Di Bartolo, I.; Manuali, E.; Pirani, S.; Bazzucchi, M.; Moscati, L.; De Mia, G.M.; Giammarioli, M. Hepatitis E virus (HEV) genotype 3 diversity: Identification of a novel HEV subtype in wild boar in Central Italy. Transbound. Emerg. Dis. 2021, 68, 2121–2129. [Google Scholar] [CrossRef]
- Bouquet, J.; Tessé, S.; Lunazzi, A.; Eloit, M.; Rose, N.; Nicand, E.; Pavio, N. Close similarity between sequences of hepatitis E virus recovered from humans and swine, France, 2008-2009. Emerg. Infect. Dis. 2011, 17, 2018–2025. [Google Scholar] [CrossRef]
- Suin, V.; Klamer, S.E.; Hutse, V.; Wautier, M.; Jacques, M.; Abady, M.; Lamoral, S.; Verburgh, V.; Thomas, I.; Brochier, B.; et al. Epidemiology and genotype 3 subtype dynamics of hepatitis E virus in Belgium, 2010 to 2017. Euro Surveill. 2019, 24, 1800141. [Google Scholar] [CrossRef]
- Porea, D.; Anita, A.; Vata, A.; Teodor, D.; Crivei, L.; Raileanu, C.; Gotu, V.; Ratoi, I.; Cozma, A.; Anita, D.; et al. Common European Origin of Hepatitis E Virus in Human Population From Eastern Romania. Front. Public Health 2020, 8, 578163. [Google Scholar] [CrossRef]
- Priemer, G.; Cierniak, F.; Wolf, C.; Ulrich, R.G.; Groschup, M.H.; Eiden, M. Co-Circulation of Different Hepatitis E Virus Genotype 3 Subtypes in Pigs and Wild Boar in North-East Germany, 2019. Pathogens 2022, 11, 773. [Google Scholar] [CrossRef] [PubMed]
- Rutjes, S.A.; Lodder, W.J.; Lodder-Verschoor, F.; van den Berg, H.H.; Vennema, H.; Duizer, E.; Koopmans, M.; de Roda Husman, A.M. Sources of hepatitis E virus genotype 3 in The Netherlands. Emerg. Infect. Dis. 2009, 15, 381–387. [Google Scholar] [CrossRef]
- De Sabato, L.; Amoroso, M.G.; Ianiro, G.; Esposito, C.; De Grossi, L.; Fusco, G.; Barone, A.; Martini, E.; Ostanello, F.; Di Bartolo, I. Detection of Hepatitis E Virus in Livers and Muscle Tissues of Wild Boars in Italy. Food Environ. Virol. 2020, 12, 1–8. [Google Scholar] [CrossRef] [PubMed]
- van der Poel, W.H.; Verschoor, F.; van der Heide, R.; Herrera, M.I.; Vivo, A.; Kooreman, M.; de Roda Husman, A.M. Hepatitis E virus sequences in swine related to sequences in humans, The Netherlands. Emerg. Infect. Dis. 2001, 7, 970–976. [Google Scholar] [CrossRef]
- de Souza, A.J.S.; Gomes-Gouvêa, M.S.; Soares, M.d.C.P.; Pinho, J.R.R.; Malheiros, A.P.; Carneiro, L.A.; dos Santos, D.R.L.; Pereira, W.L.A. HEV infection in swine from Eastern Brazilian Amazon: Evidence of co-infection by different subtypes. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 477–485. [Google Scholar] [CrossRef]
- Korsman, S.; Bloemberg, J.; Brombacher, M.; Giuricich, A.; Halley-Stott, R.P.; Kaba, M. Hepatitis E in pig-derived food products in Cape Town, South Africa, 2014. S. Afr. Med. J. 2019, 109, 584–586. [Google Scholar] [CrossRef]
- De Sabato, L.; Di Bartolo, I.; Lapa, D.; Capobianchi, M.R.; Garbuglia, A.R. Molecular Characterization of HEV Genotype 3 in Italy at Human/Animal Interface. Front. Microbiol. 2020, 11, 137. [Google Scholar] [CrossRef]
- Madden, R.G.; Wallace, S.; Sonderup, M.; Korsman, S.; Chivese, T.; Gavine, B.; Edem, A.; Govender, R.; English, N.; Kaiyamo, C.; et al. Hepatitis E virus: Western Cape, South Africa. World J. Gastroenterol. 2016, 22, 9853–9859. [Google Scholar] [CrossRef]
- Korsman, S.; Hardie, D.; Kaba, M. Hepatitis E virus in patients with acute hepatitis in Cape Town, South Africa, 2011. S. Afr. Med. J. 2019, 109, 582–583. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chauhan, R.P.; Gordon, M.L. Characterization of a Near Full-Length Hepatitis E Virus Genome of Subtype 3c Generated from Naturally Infected South African Backyard Pigs. Pathogens 2022, 11, 1030. https://doi.org/10.3390/pathogens11091030
Chauhan RP, Gordon ML. Characterization of a Near Full-Length Hepatitis E Virus Genome of Subtype 3c Generated from Naturally Infected South African Backyard Pigs. Pathogens. 2022; 11(9):1030. https://doi.org/10.3390/pathogens11091030
Chicago/Turabian StyleChauhan, Ravendra P., and Michelle L. Gordon. 2022. "Characterization of a Near Full-Length Hepatitis E Virus Genome of Subtype 3c Generated from Naturally Infected South African Backyard Pigs" Pathogens 11, no. 9: 1030. https://doi.org/10.3390/pathogens11091030