
Susceptibility of Different Life Stages of Kudzu Bug Megacopta cribraria (F.) (Hemiptera: Plataspidae) to Two Different Native Strains of Beauveria bassiana
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Kudzu Bug Collections
2.2. B. bassiana Spore Preparation
2.3. Bioassay Procedures
2.4. Statistical Analyses
3. Results
3.1. Nymph and Adult Survival and Sporulation
3.2. Nymphal and Adult Mortality
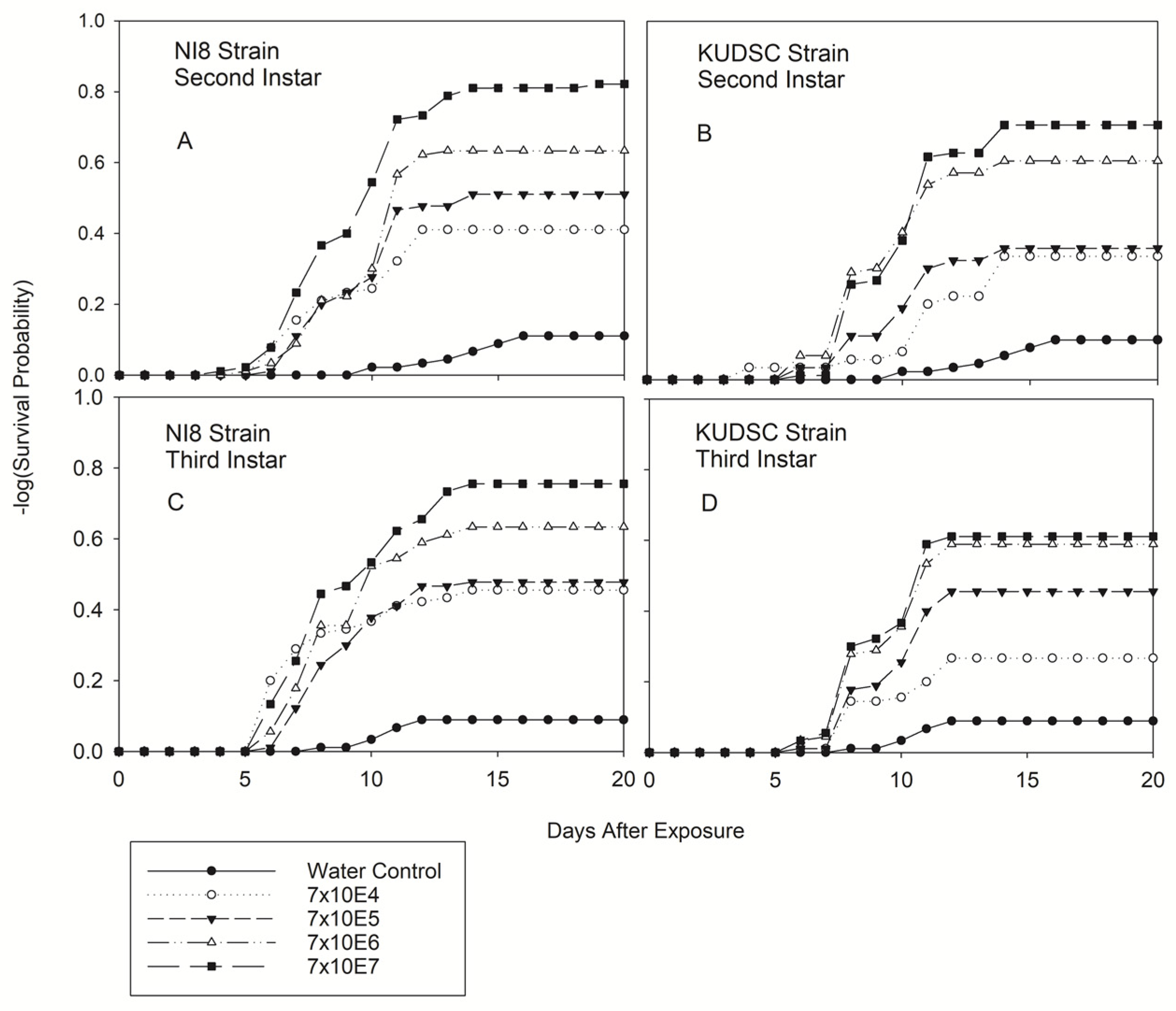
3.2.1. Second and Third Instars of Kudzu Bug
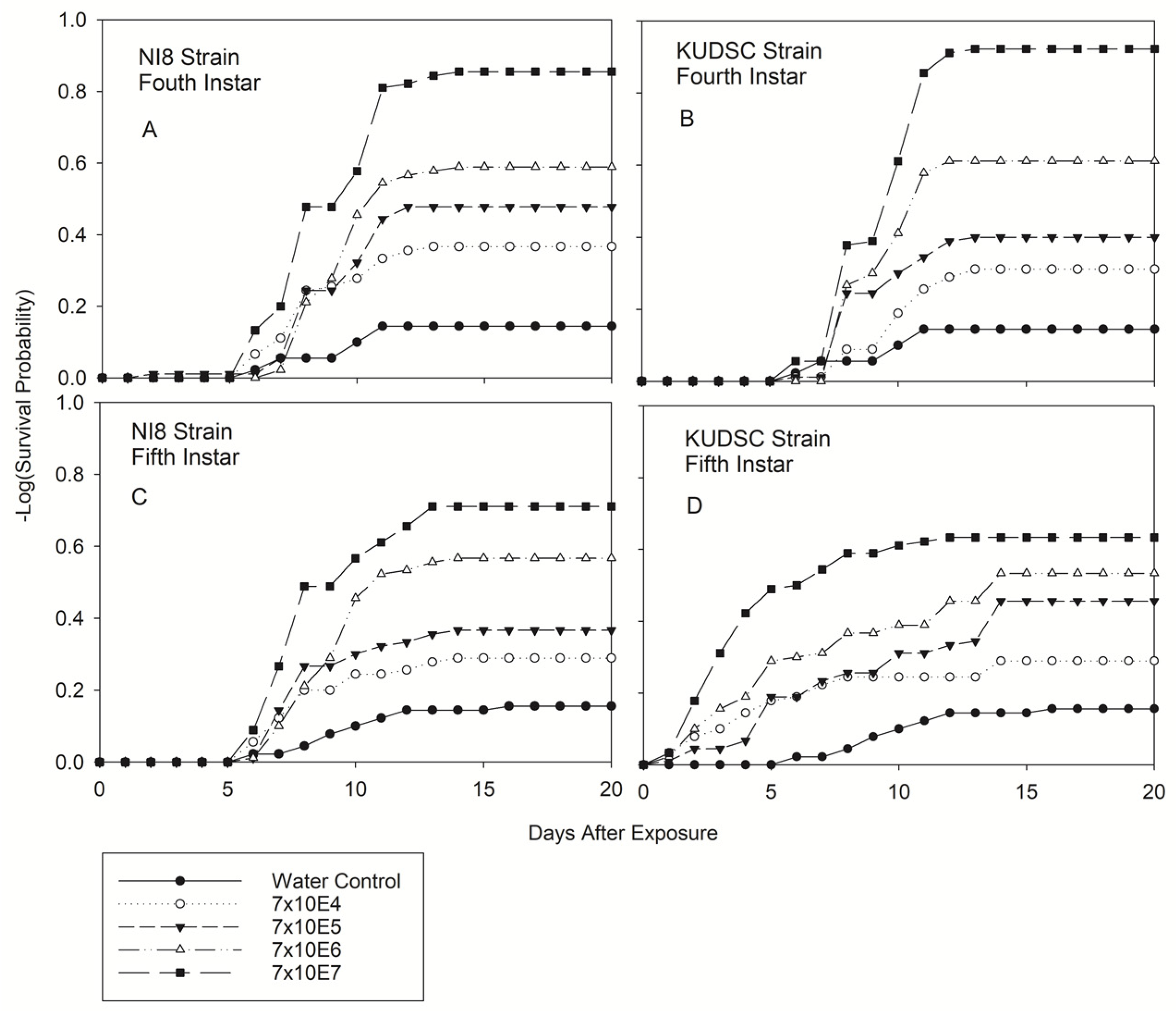
3.2.2. Fourth and Fifth Instars of Kudzu Bug
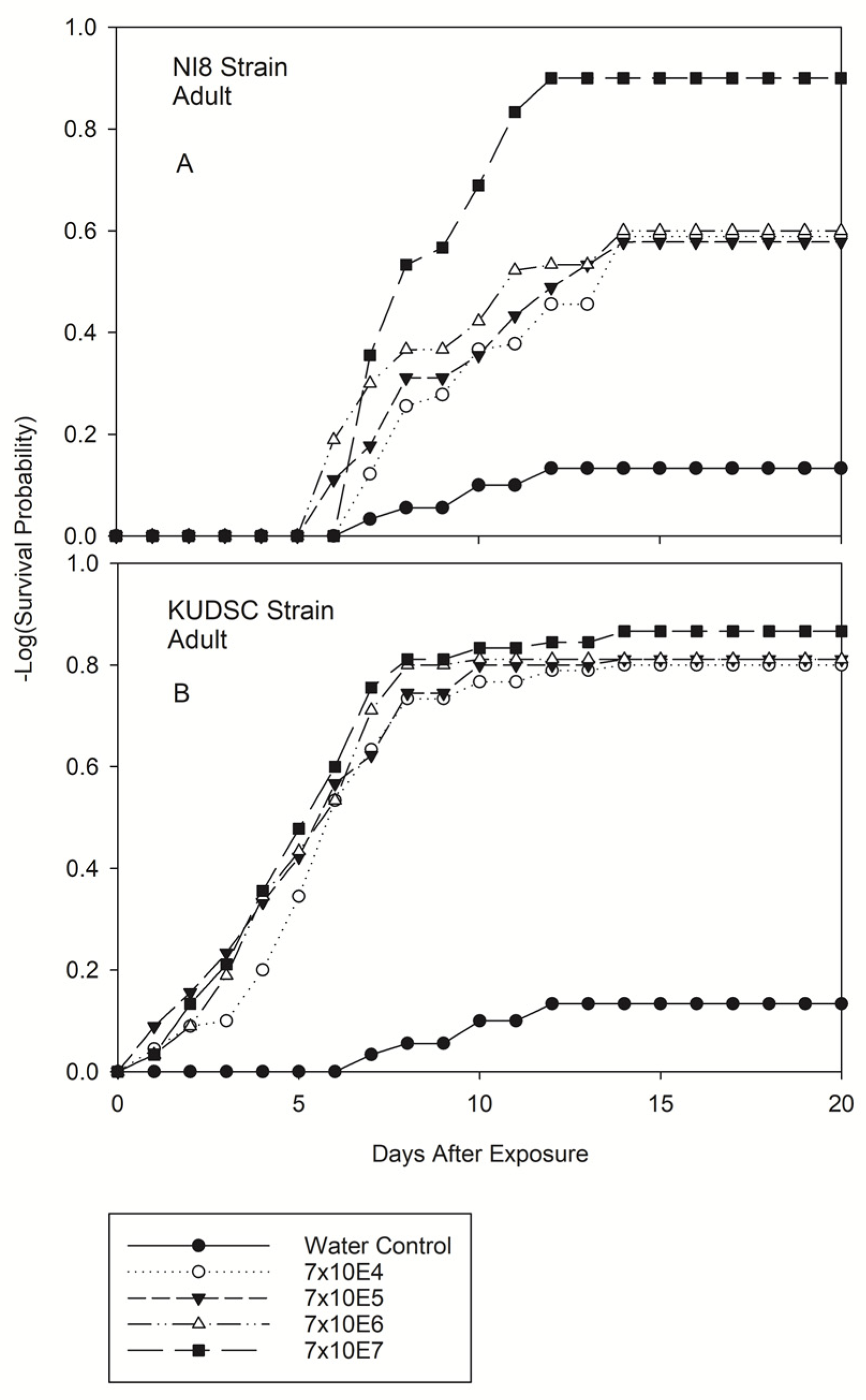
3.2.3. Adults of Kudzu Bug
3.3. Lethal Concentration and Resistance Ratio of Life Stages of Kudzu Bug
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eger, J.E., Jr.; Ames, L.M.; Suiter, D.R.; Jenkins, T.M.; Rider, D.A.; Halbert, S.E. Occurrence of the Old World bug Megacopta cribraria (Fabricius) (Heteroptera: Plataspidae) in Georgia: A serious home invader and potential legume pest. Insecta Mundi 2010, 121, 1–11. [Google Scholar]
- Eger, J.E., Jr.; Pitcher, A.J.; Halbert, S.E.; Penca, C.; Hodges, A.C. First report of Brachyplatys subaeneus (Westwood) (Hemiptera: Heteroptera: Plataspidae) in the United States. Insecta Mundi 2020, 2020, 1–6. [Google Scholar]
- Dhammi, A.; van Krestchmar, J.B.; Ponnusamy, L.; Bacheler, J.S.; Reisig, D.D.; Herbert, A.; Del Pozo-Valdivia, A.I.; Roe, R.M. Biology, Pest Status, Microbiome and Control of Kudzu Bug (Hemiptera: Heteroptera: Plataspidae): A New Invasive Pest in the U.S. Int. J. Mol. Sci. 2016, 17, 1570. [Google Scholar] [CrossRef] [PubMed]
- GBIF. Occurrence Download. Available online: https://doi.org/10.15468/dl.fsrmtb (accessed on 10 May 2021).
- Liang, W.; Tran, L.; Wiggins, G.J.; Grant, J.F.; Stewart, S.D.; Washington-Allen, R. Determining Spread Rate of Kudzu Bug (Hemiptera: Plataspidae) and Its Associations with Environmental Factors in a Heterogeneous Landscape. Environ. Entomol. 2019, 48, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Tran, L.; Washington-Allen, R.; Wiggins, G.; Stewart, S.; Vogt, J.; Grant, J. Predicting the potential invasion of kudzu bug, Megacopta cribraria (Heteroptera: Plataspidae), in North and South America and determining its climatic preference. Biol. Invasions 2018, 20, 2899–2913. [Google Scholar] [CrossRef]
- Grant, J.I.; Lamp, W.O. Cold Tolerance of Megacopta cribraria (Hemiptera: Plataspidae): An Invasive Pest of Soybeans. Environ. Entomol. 2017, 46, 1406–1414. [Google Scholar] [CrossRef]
- Lovejoy, R.T.; Lozier, J.D. Core-marginal dynamics interact with sex and temperature to influence morphology of the rapidly expanding invasive kudzu bug (Megacopta cribraria). Ecol. Entomol. 2021, 46, 790–799. [Google Scholar] [CrossRef]
- Merwin, A.C. Flight capacity increases then declines from the core to the margins of an invasive species’ range. Biol. Lett. 2019, 15, 20190496. [Google Scholar] [CrossRef]
- Lahiri, S.; Reisig, D.D. Ecology and Management of Kudzu Bug (Hemiptera: Plataspidae) in Southeastern Soybeans. J. Int. Pest Manag. 2016, 7, 14. [Google Scholar] [CrossRef]
- Medal, J.; Halbert, S.; Santa Cruz, A.; Smith, T.; Davis, B.J. Greenhouse study to determine the host range of the kudzu bug, Megacopta cribraria (Heteroptera: Plataspidae). Fla. Entomol. 2016, 99, 303–305. [Google Scholar] [CrossRef]
- Yang, L.; Hu, X.P.; Golec, J.R.; Zeng, X. Effects of Legume Species and Plant Growth Stage on Attraction, Fecundity, and Development of the Kudzu Bug (Heteroptera: Plataspidae). J. Econ. Entomol. 2018, 111, 2217–2224. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, S.; Orr, D.; Sorenson, C.E.; Cardoza, Y.J. Overwintering refuge sites for Megacopta cribraria (Hemiptera: Plataspidae). J. Entomol. Sci. 2015, 50, 69–73. [Google Scholar] [CrossRef]
- Del Pozo-Valdivia, A.I.; Reisig, D.D. First-generation Megacopta cribraria (Hemiptera: Plataspidae) can develop on soybeans. J. Econ. Entomol. 2013, 106, 533–535. [Google Scholar] [CrossRef] [PubMed]
- Seiter, N.J.; Greene, J.K.; Reay-Jones, F.P. Reduction of soybean yield components by Megacopta cribraria (Hemiptera: Plataspidae). J. Econ. Entomol. 2013, 106, 1676–1683. [Google Scholar] [CrossRef]
- Grant, J.I.; Lamp, W.O. Degree day requirements for Kudzu Bug (Hemiptera: Plataspidae), a pest of soybeans. J. Econ. Entomol. 2018, 111, 700–706. [Google Scholar] [CrossRef]
- Seiter, N.J.; Benson, E.P.; Reay-Jones, F.P.; Greene, J.K.; Zungoli, P.A. Residual efficacy of insecticides applied to exterior building material surfaces for control of nuisance infestations of Megacopta cribraria (Hemiptera: Plataspidae). J. Econ. Entomol. 2013, 106, 2448–2456. [Google Scholar] [CrossRef]
- Seiter, N.J.; Greene, J.K.; Reay-Jones, F.P.F.; Roberts, P.M.; All, J.N. Insecticidal control of Megacopta cribraria (Hemiptera: Plataspidae) in Soybean. J. Entomol. Sci. 2015, 50, 263–283. [Google Scholar] [CrossRef]
- Ruberson, J.R.; Takasu, K.; David Buntin, G.; Eger, J.E.; Gardner, W.A.; Greene, J.K.; Jenkins, T.M.; Jones, W.A.; Olson, D.M.; Roberts, P.M.; et al. From Asian curiosity to eruptive American pest: Megacopta cribraria (Hemiptera: Plataspidae) and prospects for its biological control. Appl.Entomol. Zool. 2013, 48, 3–13. [Google Scholar] [CrossRef]
- Golec, J.R.; Hu, X.P.; Ray, C.; Woodley, N.E. Strongygaster triangulifera (Diptera: Tachinidae) as a parasitoid of adults of the invasive Megacopta cribraria (Heteroptera: Plataspidae) in Alabama. J. Entomol. Sci. 2013, 48, 352–354. [Google Scholar] [CrossRef]
- Stubbins, F.L.; Agudelo, P.; Reay-Jones, F.P.F.; Greene, J.K. First report of a mermithid nematode infecting the invasive Megacopta cribraria (Hemiptera: Plataspidae) in the United States. J. Invert. Pathol. 2015, 127, 35–37. [Google Scholar] [CrossRef]
- Gardner, W.A.; Blount, J.L.; Golec, J.R.; Jones, W.A.; Hu, X.P.; Talamas, E.J.; Evans, R.M.; Dong, X.; Ray, C.H., Jr.; Buntin, G.D.; et al. Discovery of Paratelenomus saccharalis (Dodd) (Hymenoptera: Platygastridae), an egg parasitoid of Megacopta cribraria F. (Hemiptera: Plataspidae) in its expanded North American Range. J. Entomol. Sci. 2013, 48, 355–359. [Google Scholar] [CrossRef]
- Ademokoya, B.; Balusu, R.; Ray, C.; Mottern, J.; Fadamiro, H. The First Record of Ooencyrtus nezarae (Hymenoptera: Encyrtidae) on Kudzu Bug (Hemiptera: Plataspidae) in North America. J. Insect Sci. 2018, 18, 105. [Google Scholar] [CrossRef] [Green Version]
- Britt, K.; Grant, J.F.; Wiggins, G.J.; Stewart, S.D. Prevalence and Localized Infection of the Entomopathogenic Fungus Beauveria bassiana on Kudzu Bug (Hemiptera: Plataspidae) in Eastern Tennessee. J. Entomol. Sci. 2016, 51, 321–324. [Google Scholar] [CrossRef]
- Portilla, M.; Jones, W.; Perera, O.; Seiter, N.; Greene, J.; Luttrell, R. Estimation of Median Lethal Concentration of Three Isolates of Beauveria bassiana for Control of Megacopta cribraria (Heteroptera: Plataspidae) Bioassayed on Solid Lygus spp. Diet. Insects 2016, 7, 31. [Google Scholar] [CrossRef]
- Seiter, N.J.; Grabke, A.; Greene, J.K.; Kerrigan, J.L.; Reay-Jones, F.P.F. Beauveria bassiana is a pathogen of Megacopta cribraria (Hemiptera: Plataspidae) in South Carolina. J. Entomol. Sci. 2014, 49, 326–330. [Google Scholar] [CrossRef]
- Gardner, W.; Olson, D.M. Population Census of Megacopta cribraria (Hemiptera: Plataspidae) in Kudzu in Georgia, U.S.A., 2013–2016. J. Entomol. Sci. 2016, 51, 325–328. [Google Scholar] [CrossRef]
- Leland, J.E.; Snodgrass, G.L. Prevalence of naturally occurring Beauveria bassiana in Lygus lineolaris (Heteroptera: Miridae) populations from wild host plants of Mississippi. J. Agric. Urban Entomol. 2005, 21, 157–163. [Google Scholar]
- Snodgrass, G.L.; Scott, W.P.; Robbins, J.T.; Hardee, D.D. Area-wide management of the tarnished plant bug by reduction of early-season wild host plant density. Southwest. Entomol. 2000, 23, 59–66. [Google Scholar]
- Parys, K.A.; Portilla, M. Effectiveness of Beauveria bassiana against Piezodorus guildinii (Hemiptera: Pentatomidae), a key pest of soybeans in the neotropics. Biocontrol Sci. Technol. 2020, 30, 451–461. [Google Scholar] [CrossRef]
- Portilla, M.; Abbas, H.K.; Accinelli, C.; Luttrell, R. Laboratory and field investigation on compatibility of Beauveria bassiana (Hypocreales: Clavicipotaceae) spores with a sprayable bioplastic formulation for application in the biocontrol of tarnished plant bug in cotton. J. Econ. Entomol. 2018, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Portilla, M.; Snodgrass, G.; Luttrell, R. A novel bioassay to evaluate the potential of Beauvaria bassiana strain NI8 and the insect growth regulator novaluron against Lygus lineolaris on a non-autoclaved solid artificial diet. J. Insect Sci. 2014, 14, 1–13. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute. SAS 9.4 User’s Guide; SAS Institute: Cary, NC, USA, 2018. [Google Scholar]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Forlani, L.; Pedrini, N.; Girotti, J.R.; Mijailovsky, S.J.; Cardozo, R.M.; Gentile, A.G.; Hernandez-Suarez, C.M.; Rabinovich, J.E.; Juarez, M.P. Biological Control of the Chagas Disease Vector Triatoma infestans with the Entomopathogenic Fungus Beauveria bassiana Combined with an Aggregation Cue: Field, Laboratory and Mathematical Modeling Assessment. PLoS Negl. Trop. Dis. 2015, 9, e0003778. [Google Scholar] [CrossRef]
- Goodell, P.B. Managing Lygus in the Landscape. In Proceedings of the California Chapter of the American Agronomy Society Plant and Soil Conference, Anaheim, CA, USA, 7–8 February 2001. [Google Scholar]
- Shapiro-Ilan, D.I.; Reilly, C.C.; Hotchkiss, M.W. Comparative impact of artificial selection for fungicide resistance on Beauveria bassiana and Metarhizium brunneum. Environ. Entomol. 2011, 40, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Knight, I.A.; Roberts, P.M.; Gardner, W.A.; Oliver, K.M.; Jacobson, A.; Toews, M.D. Effect of Beauveria bassiana (Hypocreales: Clavicipitaceae) and Fungicide Applications on Megacopta cribraria (Hemiptera: Plataspidae) in Soybean. Crop Forage Turfgrass Manag. 2017, 3, 34. [Google Scholar] [CrossRef]
- Niland, E.S.; Lewallen, C.; Carrillo, C.; Perez, L.; Thompson, A.; Gardner, W.A. Quantification and Molecular Identification of Beauveria bassiana (Hypocreales: Clavicipitaceae) in Soils Associated with Megacopta cribraria (Hemiptera: Plastaspidae). J. Entomol. Sci. 2019, 54, 93–98. [Google Scholar] [CrossRef]
- Olson, D.M.; Tillman, G.; Toews, M. Biopesticide and Parasitoid Effects on Megacopta cribraria (Hemiptera: Plataspidae) Life Stage Density and Egg Parasitism in Soybean. J. Entomol. Sci. 2021, 56, 272–277. [Google Scholar] [CrossRef]
- Lahiri, S.; Reisig, D.D.; Dean, L.L.; Reay-Jones, F.P.F.; Greene, J.K.; Carter, T.E.; Mian, R.; Fallen, B.D. Mechanisms of Soybean Host-Plant Resistance Against Megacopta cribraria (Hemiptera: Plataspidae). Environ. Entomol. 2020, 49, 876–885. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatments 1 (Spores/mL) | Kudzu Bug Stages (% Mean) | ||||
|---|---|---|---|---|---|
| 2nd Instar | 3rd Instar | 4th Instar | 5th Instar | Adult | |
| Day 10 | |||||
| Control | 12.21 ± 3.54 de | 10.0 ± 3.23 d | 13.33 ± 3.6 c | 13.33 ± 3.6 d | 14.44 ± 3.72 d |
| 7 × 104-NI8 | 24.4 ± 4.63 bcde | 36.7 ± 5.1 abc | 26.66 ± 4.67 bc | 24.44 ± 4.56 dc | 38.89 ± 5.17 c |
| 7 × 105-NI8 | 27.82 ± 4.74 bcde | 40.0 ± 5.21 ab | 32.22 ± 4.95 bc | 31.11 ± 4.91 dc | 37.78 ± 5.14 c |
| 7 × 106-NI8 | 31.12 ± 4.93 cdb | 53.3 ± 5.35 a | 43.33 ± 5.25 ab | 45.56 ± 5.28 abc | 51.11 ± 5.30 bc |
| 7 × 107-NI8 | 56.70 ± 5.30 a | 55.6 ± 5.2 a | 56.67 ± 5.25 a | 57.78 ± 5.24 a | 68.89 ± 4.91 ab |
| 7 × 104-KUDSC | 7.81 ± 2.80 e | 15.6 ± 3.82 cd | 22.22 ± 4.41 bc | 31.11 ± 4.91 dc | 76.67 ± 4.48 a |
| 7 × 105-KUDSC | 20.00 ± 4.24 cde | 27.8 ± 4.76 bcd | 27.78 ± 4.75 bc | 26.67 ± 4.69 dc | 80.00 ± 4.24 a |
| 7 × 106-KUDSC | 42.23 ± 5.21 ab | 35.6 ± 5.14 abc | 42.22 ± 5.24 ab | 38.89 ± 5.17 bc | 81.11 ± 4.15 a |
| 7 × 107-KUDSC | 40.05 ± 5.28 abc | 53.3 ± 5.3 a | 63.33 ± 5.11 a | 64.44 ± 4.5 a | 86.67 ± 3.6 a |
| Day 20 | |||||
| Control | 21.12 ± 4.39 f | 17.80 ± 4.12 d | 21.11 ± 4.33 d | 16.67 ± 2.53 e | 15.56 ± 3.84 e |
| 7 × 104-NI8 | 41.16 ± 5.25 edf | 46.75 ± 5.3 bc | 35.56 ± 5.07 cd | 28.89 ± 4.8 ed | 62.22 ± 5.14 dc |
| 7 × 105-NI8 | 51.13 ± 5.33 edc | 50.03 ± 5.3 b | 48.89 ± 5.3 bc | 37.78 ± 5.14 edc | 56.67 ± 5.25 d |
| 7 × 106-NI8 | 64.40 ± 5.10 abc | 64.41 ± 4.51 ab | 58.89 ± 5.22 b | 56.67 ± 5.25 abc | 70.00 ± 4.85 bcd |
| 7 × 107-NI8 | 84.41 ± 3.83 a | 76.72 ± 4.8 a | 85.56 ± 3.73 a | 72.22 ± 5.28 a | 90.00 ± 3.18 a |
| 7 × 104-KUDSC | 37.78 ± 5.17 ef | 25.63 ± 4.68 cd | 33.33 ± 4.5 cd | 45.56 ± 5.28 bdc | 80.00 ± 4.23 abc |
| 7 × 105-KUDSC | 37.03 ± 5.12 ef | 47.83 ± 5.33 b | 37.78 ± 5.14 cd | 31.11 ± 4.9 ed | 81.11 ± 4.15 abc |
| 7 × 106-KUDSC | 62.20 ± 5.15 edc | 64.40 ± 5.1 ab | 63.33 ± 5.11 b | 53.33 ± 5.29 abc | 81.11 ± 4.45 abc |
| 7 × 107-KUDSC | 73.33 ± 4.70 ab | 85.65 ± 3.71 a | 95.56 ± 2.18 a | 66.67 ± 4.5 ab | 86.67 ± 3.6 ab |
| Stage | Concentration Response (Spores/mm2) | |||||||
|---|---|---|---|---|---|---|---|---|
| n | Slope ± SE | LC50 (95% CI) | Probit Trend | RR50 (95% CI) * | ||||
| Test for Slope | Test for GoF | |||||||
| X2 | p > X2 | X2 | p > X2 | |||||
| 10 Days | ||||||||
| 2nd Instar | 450 | 0.18 ± 0.07 | 789 (77–3.1 × 1026) | 5.3 | 0.0214 | 1.89 | 0.0415 | 17 (1–309) |
| 3rd Instar | 450 | 0.09 ± 0.03 | 199 (24–7.3 × 104) | 8.51 | 0.0035 | 0.68 | 0.7436 | 3 (0.13–50) |
| 4th Instar | 450 | 0.15 ± 0.04 | 480 (101–1.0 × 104) | 14.18 | 0.0002 | 1.1 | 0.3551 | 8 (0.61–110) |
| 5th Instar | 450 | 0.18 ± 0.04 | 307 (85–2.4 ×103) | 17.29 | 0.0001 | 1.44 | 0.9265 | 6 (0.57–73) |
| Adult | 450 | 0.15 ± 0.04 | 73 (5–4.5 × 103) | 9.3 | 0.0023 | 1.75 | 0.0568 | 1 |
| 20 Days | ||||||||
| 2nd Instar | 450 | 0.22 ± 0.04 | 13 (3–36) | 28.65 | 0.0001 | 1.22 | 0.2680 | 16 (0.46–588) |
| 3rd Instar | 450 | 0.15 ± 0.03 | 11 (2–45) | 18.46 | 0.0001 | 1.48 | 0.2265 | 10 (0.22–429) |
| 4th Instar | 450 | 0.25 ± 0.05 | 20 (6–51) | 28.03 | 0.0001 | 1.23 | 0.2624 | 32 (0.94–1096) |
| 5th Instar | 450 | 0.17 ± 0.03 | 133 (33–879) | 18.97 | 0.0001 | 1.24 | 0.2532 | 183 (5–6494) |
| Adult | 450 | 0.54 ± 0.05 | 1 (0.0005–6) | 10.09 | 0.0015 | 1.93 | 0.0363 | 1 |
| Stage | Concentration Response (Spores/mm2) | |||||||
|---|---|---|---|---|---|---|---|---|
| n | Slope ± SE | LC50 (95% CI) | Probit Trend | RR50 (95% CI) * | ||||
| Test for Slope | Test for GoF | |||||||
| X2 | p > X2 | X2 | p > X2 | |||||
| 10 Days | ||||||||
| 2nd Instar | 450 | 0.21 ± 0.04 | 1051 (286–12,053) | 22.54 | 0.0001 | 1.52 | 0.1249 | 438 (71–2694) |
| 3rd Instar | 450 | 0.19 ± 0.04 | 464 (140–3705) | 19.93 | 0.0001 | 1.04 | 0.4043 | 184 (35–981) |
| 4th Instar | 450 | 0.23 ± 0.05 | 205 (70–864) | 17.42 | 0.0001 | 1.11 | 0.3462 | 67 (15–360) |
| 5th Instar | 450 | 0.22 ± 0.05 | 182 (61–748) | 19.56 | 0.0001 | 0.83 | 0.5931 | 58 (13–271) |
| Adult | 450 | 0.23 ± 0.03 | 3 (1–7) | 47.51 | 0.0001 | 1.31 | 0.2159 | 1 |
| 20 Days | ||||||||
| 2nd Instar | 450 | 0.21 ± 0.04 | 49 (12–164) | 19.66 | 0.0001 | 0.67 | 0.7573 | 25 (1.32–482) |
| 3rd Instar | 450 | 0.30 ± 0.04 | 17 (7–36) | 44.11 | 0.0001 | 0.84 | 0.5889 | 19 (1.26–284) |
| 4th Instar | 450 | 0.52 ± 0.15 | 28 (2–65) | 11.28 | 0.0008 | 1.84 | 0.0479 | 32 (1.74–580) |
| 5th Instar | 450 | 0.21 ± 0.04 | 82 (27–297) | 25.38 | 0.0001 | 1.51 | 0.1267 | 68 (4–1117) |
| Adult | 450 | 0.21 ± 0.06 | 1.14 (0.006–7) | 11.62 | 0.0007 | 3.22 | 0.0004 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glover, J.P.; Portilla, M.; Parys, K.; Allen, C.; George, J.; Reddy, G.V.P. Susceptibility of Different Life Stages of Kudzu Bug Megacopta cribraria (F.) (Hemiptera: Plataspidae) to Two Different Native Strains of Beauveria bassiana. Pathogens 2022, 11, 1028. https://doi.org/10.3390/pathogens11091028
Glover JP, Portilla M, Parys K, Allen C, George J, Reddy GVP. Susceptibility of Different Life Stages of Kudzu Bug Megacopta cribraria (F.) (Hemiptera: Plataspidae) to Two Different Native Strains of Beauveria bassiana. Pathogens. 2022; 11(9):1028. https://doi.org/10.3390/pathogens11091028
Chicago/Turabian StyleGlover, James Paul, Maribel Portilla, Katherine Parys, Clint Allen, Justin George, and Gadi V. P. Reddy. 2022. "Susceptibility of Different Life Stages of Kudzu Bug Megacopta cribraria (F.) (Hemiptera: Plataspidae) to Two Different Native Strains of Beauveria bassiana" Pathogens 11, no. 9: 1028. https://doi.org/10.3390/pathogens11091028