
Differentially Represented Proteins in Response to Infection with Mycobacterium tuberculosis Identified by Quantitative Serum Proteomics in Asian Elephants
 ,
, 
 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Serum Samples from TB-Positive (TB+) and TB-Negative (TB−) Elephants
2.2. Diagnostic of Tuberculosis
2.3. Serum Proteomics
2.4. Quality Control of Proteomics Data
2.5. Data Analysis
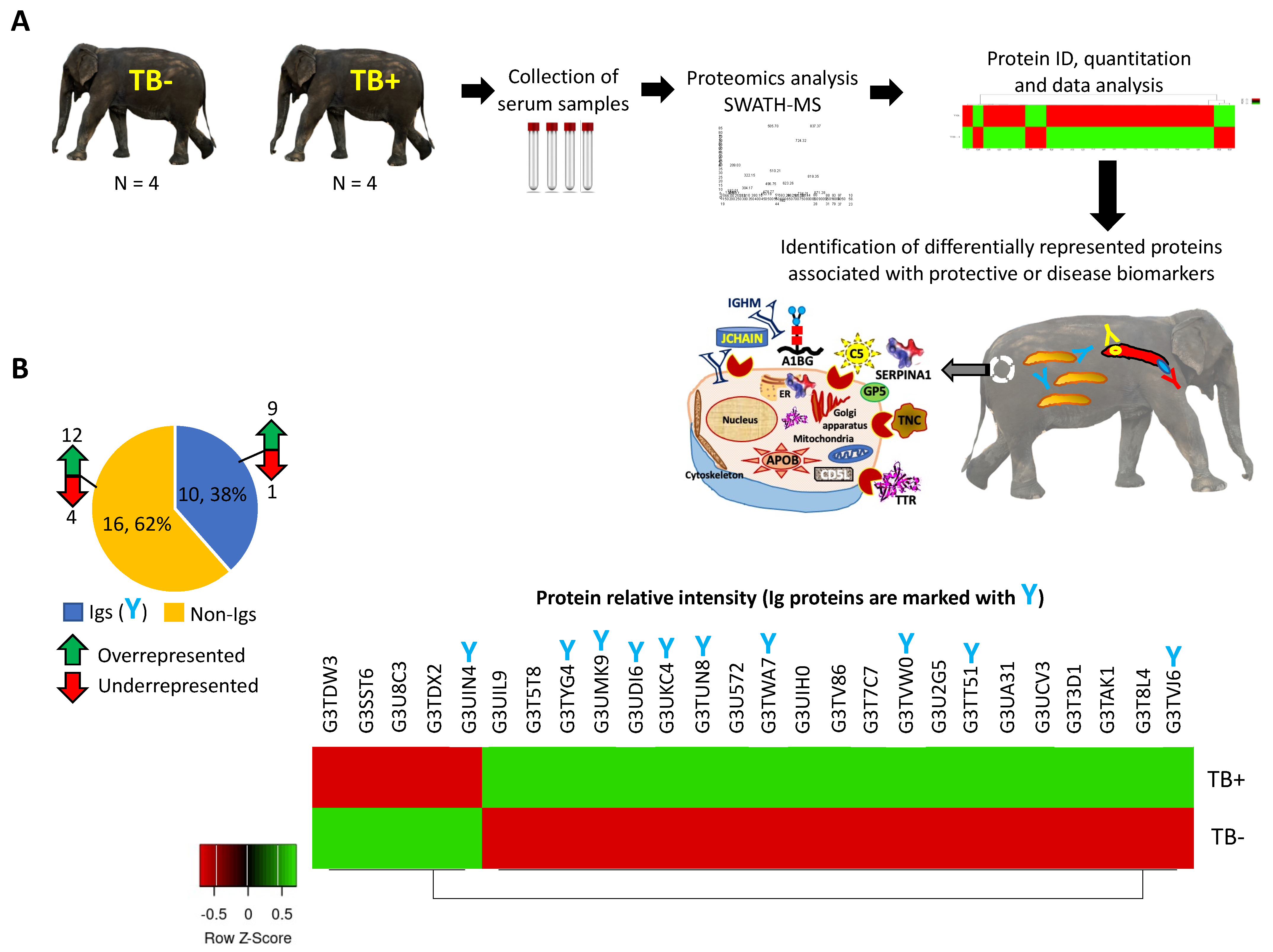
3. Results
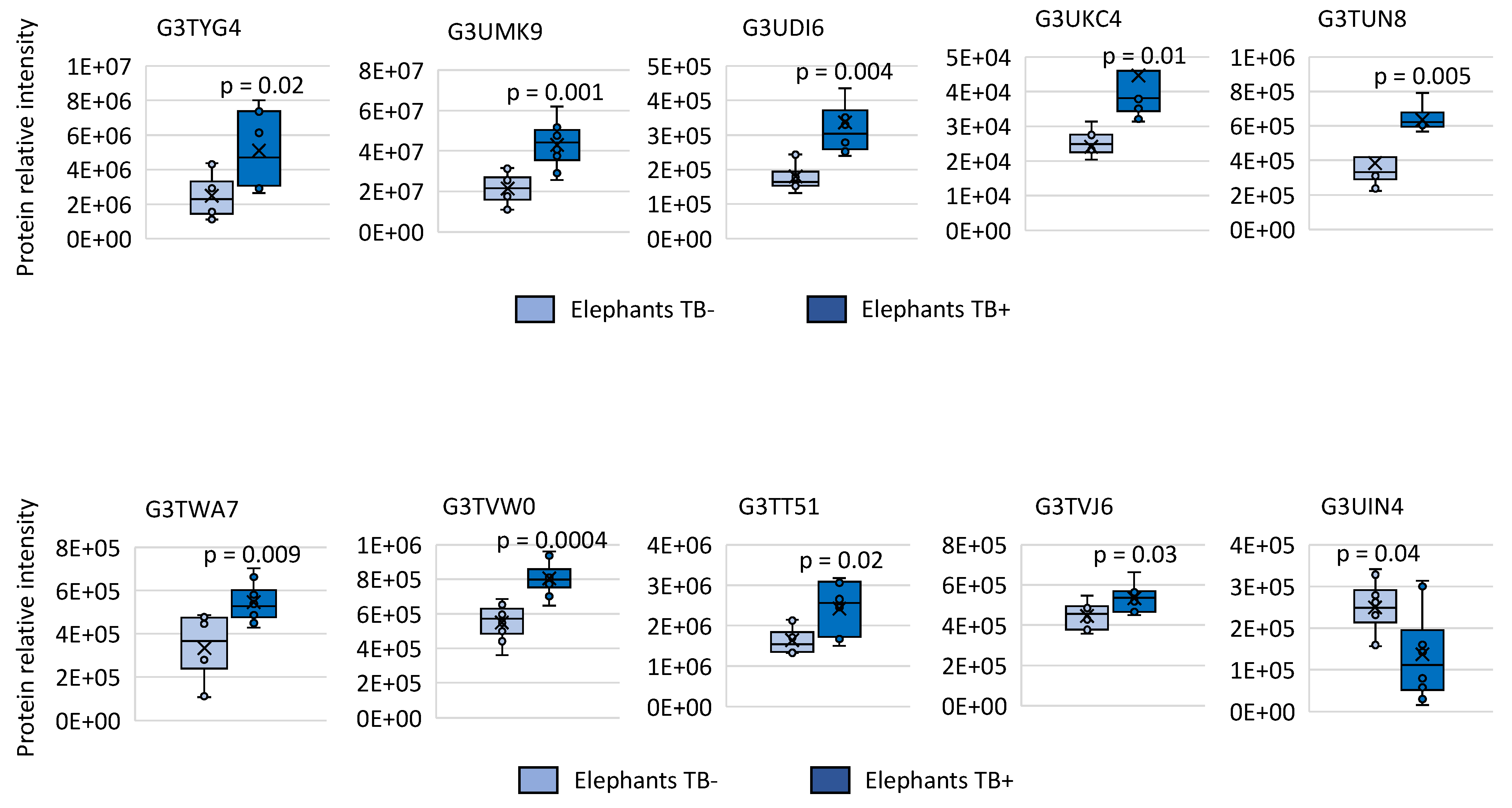
3.1. Identification of Differentially Represented Elephant Serum Proteins in Response to TB by Quantitative Serum Proteomics
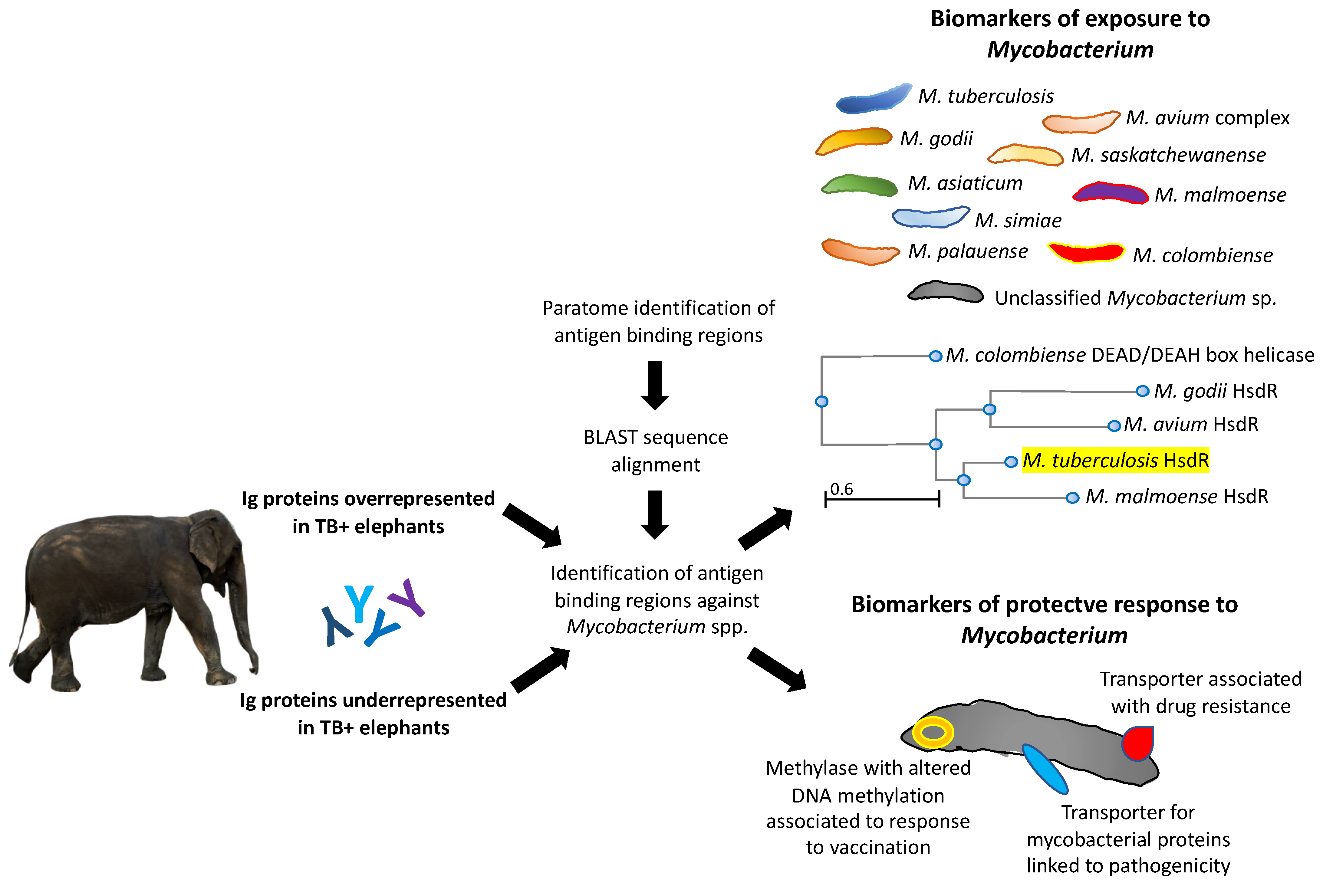
3.2. Characterization of Ig Protein Profiles and Correlation with Mycobacterial Infection or Host Protective Capacity
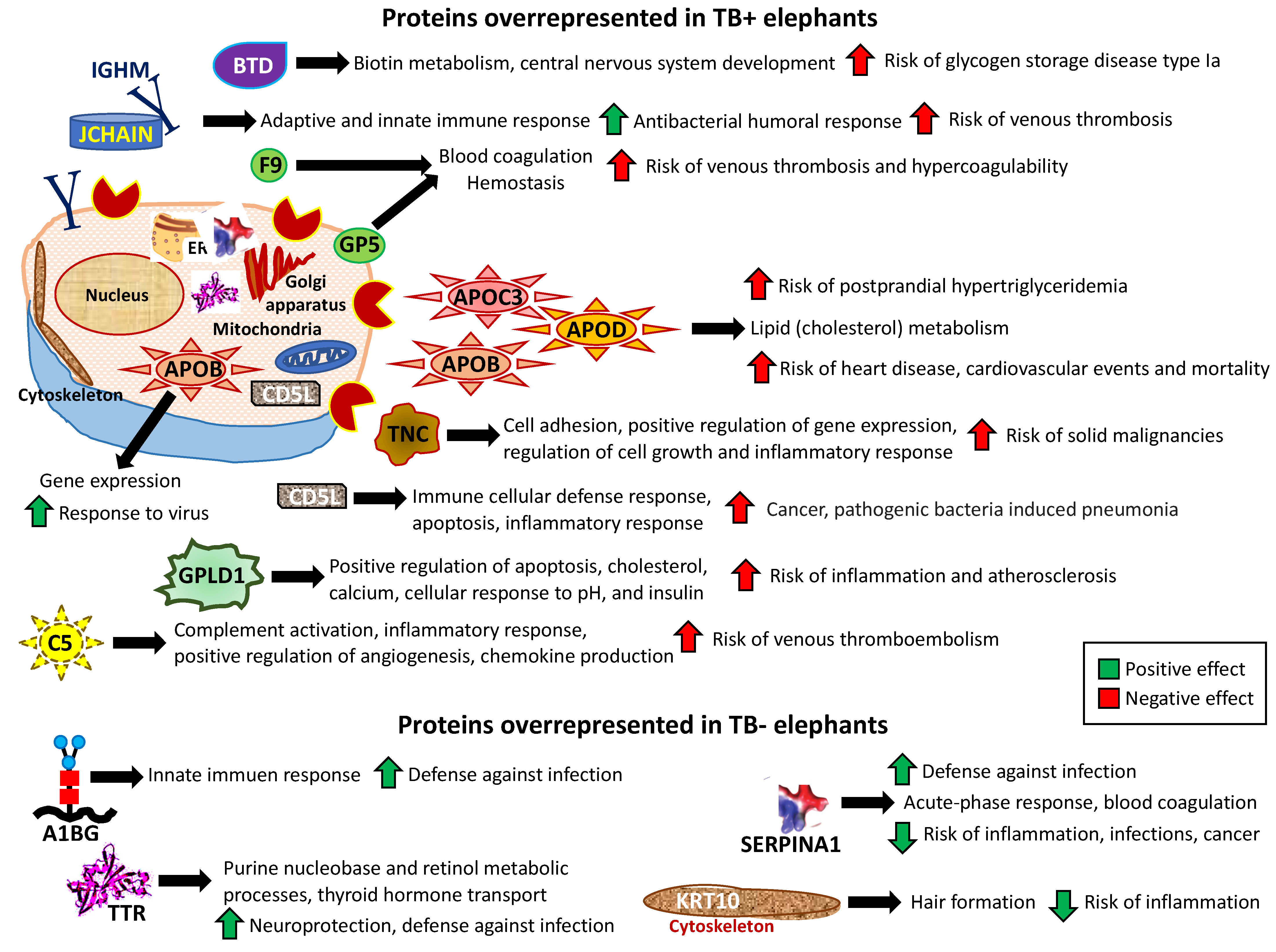
3.3. Characterization of Non-Ig Protein Profiles and Correlation with TB
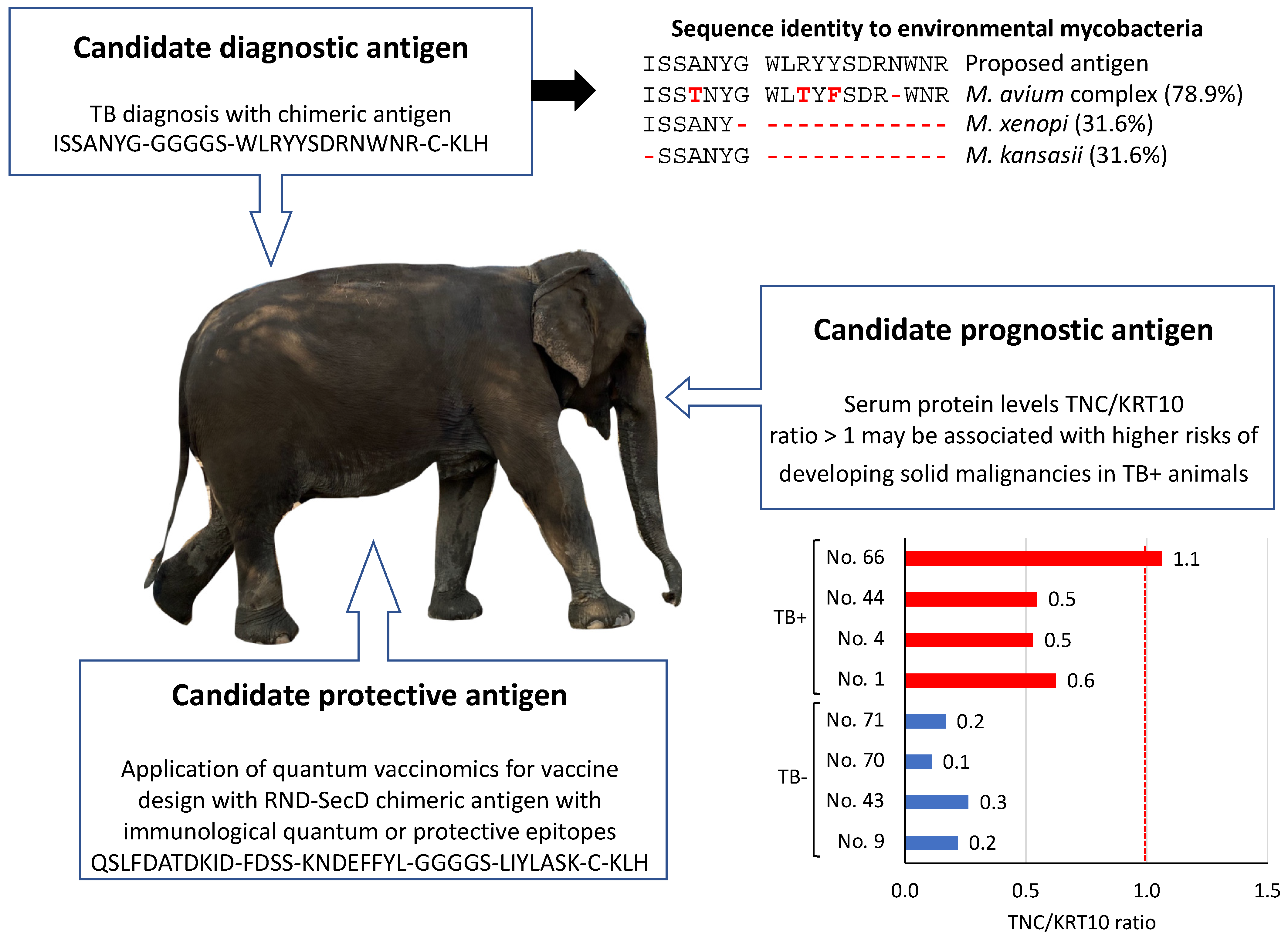
3.4. Implications for Translational Biotechnology
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO (2021) Tuberculosis. Available online: https://www.who.int/news-room/fact-sheets/detail/tuberculosis (accessed on 16 March 2022).
- Brites, D.; Gagneux, S. Co-evolution ofMycobacterium tuberculosisandHomo sapiens. Immunol. Rev. 2015, 264, 6–24. [Google Scholar] [CrossRef] [PubMed]
- Malone, K.M.; Gordon, S.V. Mycobacterium tuberculosis Complex Members Adapted to Wild and Domestic Animals. Adv. Exp. Med. Biol. 2017, 1019, 135–154. [Google Scholar] [CrossRef]
- Talip, B.A.; Sleator, R.D.; Lowery, C.J.; Dooley, J.S.; Snelling, W.J. An Update on Global Tuberculosis (TB). Infect. Dis. Res. Treat. 2013, 6, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Iyer, A.K. Veterinary science in India, ancient and modern with special reference to tuberculosis. Agric. Livest. India 1937, 7, 718–724. [Google Scholar]
- Mikota, S.K.; Larsen, R.S.; Montai, R.J. Tuberculosis in Elephants in North America. Zoo Biol. 2000, 19, 393–403. [Google Scholar] [CrossRef]
- Lewerin, S.S.; Eld, K.; Bölske, G.; Olsson, S.; Röken, B.; Ghebremichael, S.; Koivula, T.; Källenius, G. Outbreak of Mycobacterium tuberculosis infection among captive Asian elephants in a Swedish zoo. Veter- Rec. 2005, 156, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Mikota, S.K. A Brief History of TB in Elephants. Available online: https://www.aphis.usda.gov/animal_welfare/downloads/elephant/ABriefHistoryofTBinElephants.pdf (accessed on 17 November 2021).
- Pradhan, N.M.B.; Williams, A.C.; Dhakal, M. Current Status of Asian Elephants in Nepal. Gajah 2020, 35, 87–92. [Google Scholar]
- Shrestha, S.; Shrestha, J. Asian elephants and their status in Nepal: A review. J. Agric. Nat. Resour. 2021, 4, 227–237. [Google Scholar] [CrossRef]
- Bale, R. Captive Elephants Freed From a Life in Chains. Available online: https://www.nationalgeographic.com/animals/article/wildlife-solar-fence-chain-free-elephants-nepal (accessed on 6 April 2022).
- Szydlowski, M. Elephants in Nepal: Correlating disease, tourism, and welfare. J. Appl. Anim. Welf. Sci. 2022, 25, 126–138. [Google Scholar] [CrossRef]
- Paudel, S.; Mikota, S.K.; Nakajima, C.; Gairhe, K.P.; Maharjan, B.; Thapa, J.; Poudel, A.; Shimozuru, M.; Suzuki, Y.; Tsubota, T. Molecular characterization of Mycobacterium tuberculosis isolates from elephants of Nepal. Tuberculosis 2014, 94, 287–292. [Google Scholar] [CrossRef]
- Thapa, J.; Nakajima, C.; Maharjan, B.; Poudell, A.; Suzuki, Y. Molecular characterization of Mycobacterium orygis isolates from wild animals of Nepal. Jpn. J. Vet. Res. 2015, 63, 15–158. [Google Scholar]
- Thapa, J.; Paudel, S.; Sadaula, A.; Shah, Y.; Maharjan, B.; Kaufman, G.E.; McCauley, D.; Gairhe, K.P.; Tsubota, T.; Suzuki, Y.; et al. Mycobacterium orygis–Associated Tuberculosis in Free-Ranging Rhinoceros, Nepal, 2015. Emerg. Infect. Dis. 2016, 22, 570–572. [Google Scholar] [CrossRef] [PubMed]
- Ellwanger, J.H.; Chies, J.A.B. Zoonotic spillover: Understanding basic aspects for better prevention. Genet. Mol. Biol. 2021, 44, e20200355. [Google Scholar] [CrossRef] [PubMed]
- Gompo, T.R.; Shrestha, A.; Ranjit, E.; Gautam, B.; Ale, K.; Shrestha, S.; Bhatta, D.D. Risk factors of tuberculosis in human and its association with cattle TB in Nepal: A one health approach. One Health 2020, 10, 100156. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Kerr, T.J.; de Waal, C.R.; Goosen, W.J.; Streicher, E.M.; Hausler, G.; Rossouw, L.; Manamela, T.; van Schalkwyk, L.; Kleynhans, L.; et al. Mycobacterium bovis Infection in Free-Ranging African Elephants. Emerg. Infect. Dis. 2021, 27, 990–992. [Google Scholar] [CrossRef]
- Greenwald, R.; Lyashchenko, O.; Esfandiari, J.; Miller, M.; Mikota, S.; Olsen, J.H.; Ball, R.; Dumonceaux, G.; Schmitt, D.; Moller, T.; et al. Highly Accurate Antibody Assays for Early and Rapid Detection of Tuberculosis in African and Asian Elephants. Clin. Vaccine Immunol. 2009, 16, 605–612. [Google Scholar] [CrossRef]
- Lyashchenko, K.P.; Greenwald, R.; Esfandiari, J.; Mikota, S.; Miller, M.; Moller, T.; Vogelnest, L.; Gairhe, K.P.; Robbe-Austerman, S.; Gai, J.; et al. Field Application of Serodiagnostics To Identify Elephants with Tuberculosis prior to Case Confirmation by Culture. Clin. Vaccine Immunol. 2012, 19, 1269–1275. [Google Scholar] [CrossRef]
- Mikota, S.K.; Kaufman, G.; Dhakal, I.P.; Pandey, B.D. Tuberculosis in Nepal: Elephants, Human, Livestock and Wildlife. Proceedings AAZV AAWV Joint Conference. 2009. Available online: http://www.rhinoresourcecenter.com/pdf_files/150/1506973531.pdf. (accessed on 10 February 2022).
- Paudel, S.; Mikota, S.K.; Thapa, J.; Lyashchenko, K.P.; Gairhe, K.P.; Dhakal, I.P.; Subedi, N.; Maharjan, B.; Subedi, S.; Kaufman, G.E.; et al. Serodiagnosis of elephant tuberculosis: A useful tool for early identification of infected elephants at the captive-wild interface. Eur. J. Wildl. Res. 2018, 64, 70. [Google Scholar] [CrossRef]
- Backues, K.A.; Wiedner, E.B. Recommendations for the diagnosis, treatment and management of tuberculosis, Mycobacterium tuberculosis, in elephants in human care. Int. Zoo Yearb. 2019, 53, 116–127. [Google Scholar] [CrossRef]
- Goosen, W.J.; Kerr, T.; Kleynhans, L.; Buss, P.; Cooper, D.; Warren, R.M.; Van Helden, P.D.; Schröder, B.; Parsons, S.; Miller, M.A. The VetMAX™ M. tuberculosis complex PCR kit detects MTBC DNA in antemortem and postmortem samples from white rhinoceros (Ceratotherium simum), African elephants (Loxodonta africana) and African buffaloes (Syncerus caffer). BMC Veter- Res. 2020, 16, 1–6. [Google Scholar] [CrossRef]
- Songthammanuphap, S.; Puthong, S.; Pongma, C.; Buakeaw, A.; Prammananan, T.; Warit, S.; Tipkantha, W.; Kaewkhunjob, E.; Yindeeyoungyeon, W.; Palaga, T. Detection of Mycobacterium tuberculosis complex infection in Asian elephants (Elephas maximus) using an interferon gamma release assay in a captive elephant herd. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pohlentz, G.; Kölbl, S.; Peter-Katalinić, J. High sequence coverage by in-capillary proteolysis of native proteins and simultaneous analysis of the resulting peptides by nanoelectrospray ionization-mass spectrometry and tandem mass spectrometry. PROTEOMICS 2005, 5, 1758–1763. [Google Scholar] [CrossRef] [PubMed]
- Cappellini, E.; Jensen, L.J.; Szklarczyk, D.; Ginolhac, A.; da Fonseca, R.A.R.; Stafford, J.T.W.; Holen, S.R.; Collins, M.J.; Orlando, L.; Willerslev, E.; et al. Proteomic Analysis of a Pleistocene Mammoth Femur Reveals More than One Hundred Ancient Bone Proteins. J. Proteome Res. 2011, 11, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Yazhini, A.; Srinivasan, N.; Sandhya, S. Signatures of conserved and unique molecular features in Afrotheria. Sci. Rep. 2021, 11, 1–15. [Google Scholar] [CrossRef]
- Villar, M.; Urra, J.M.; Rodríguez-Del-Río, F.J.; Artigas-Jerónimo, S.; Jiménez-Collados, N.; Ferreras-Colino, E.; Contreras, M.; de Mera, I.G.F.; Estrada-Peña, A.; Gortázar, C.; et al. Characterization by Quantitative Serum Proteomics of Immune-Related Prognostic Biomarkers for COVID-19 Symptomatology. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Yi, X.; Sun, Y.; Bi, X.; Du, J.; Zhang, C.; Quan, S.; Zhang, F.; Sun, R.; Qian, L.; et al. Proteomic and Metabolomic Characterization of COVID-19 Patient Sera. Cell 2020, 182, 59–72.e15. [Google Scholar] [CrossRef]
- Kunik, V.; Ashkenazi, S.; Ofran, Y. Paratome: An online tool for systematic identification of antigen-binding regions in antibodies based on sequence or structure. Nucleic Acids Res. 2012, 40, W521–W524. [Google Scholar] [CrossRef]
- Gillespie, M.; Jassal, B.; Stephan, R.; Milacic, M.; Rothfels, K.; Senff-Ribeiro, A.; Griss, J.; Sevilla, C.; Matthews, L.; Gong, C.; et al. The reactome pathway knowledgebase 2022. Nucleic Acids Res 2022, 50, D687–D692. [Google Scholar] [CrossRef]
- Fabregat, A.; Korninger, F.; Viteri, G.; Sidiropoulos, K.; Marin-Garcia, P.; Ping, P.; Wu, G.; Stein, L.; D’Eustachio, P.; Hermjakob, H. Reactome graph database: Efficient access to complex pathway data. PLOS Comput. Biol. 2018, 14, e1005968. [Google Scholar] [CrossRef]
- Lyashchenko, K.P.; Gortázar, C.; Miller, M.A.; Waters, W.R. Spectrum of antibody profiles in tuberculous elephants, cervids, and cattle. Veter- Microbiol. 2018, 214, 89–92. [Google Scholar] [CrossRef]
- Yong, H.; Choi, G.-E.; Lee, B.S.; Whang, J.; Shin, S.J. Disseminated Infection Due toMycobacterium aviumSubsp.aviumin an Asian Elephant (Elephas maximus). J. Zoo Wildl. Med. 2011, 42, 743–746. [Google Scholar] [CrossRef] [PubMed]
- Ong, B.L.; Ngeow, Y.F.; Razak, M.F.A.A.; Yakubu, Y.; Zakaria, Z.; Mutalib, A.R.; Hassan, L.; Ng, H.F.; Verasahib, K. Tuberculosis in captive Asian elephants (Elephas maximus) in Peninsular Malaysia. Epidemiol. Infect. 2013, 141, 1481–1487. [Google Scholar] [CrossRef] [PubMed]
- Zachariah, A.; Pandiyan, J.; Madhavilatha, G.; Mundayoor, S.; Chandramohan, B.; Sajesh, P.; Santhosh, S.; Mikota, S.K. Mycobacterium tuberculosis in Wild Asian Elephants, Southern India. Emerg. Infect. Dis. 2017, 23, 504–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Primm, T.P.; Lucero, C.A.; Falkinham, J.O. Health Impacts of Environmental Mycobacteria. Clin. Microbiol. Rev. 2004, 17, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. RND transporters in the living world. Res. Microbiol. 2018, 169, 363–371. [Google Scholar] [CrossRef]
- De Rossi, E.; Aínsa, J.A.; Riccardi, G. Role of mycobacterial efflux transporters in drug resistance: An unresolved question. FEMS Microbiol. Rev. 2006, 30, 36–52. [Google Scholar] [CrossRef]
- Verma, D.; Parasa, V.R.; Raffetseder, J.; Martis, M.; Mehta, R.B.; Netea, M.; Lerm, M. Anti-mycobacterial activity correlates with altered DNA methylation pattern in immune cells from BCG-vaccinated subjects. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Nijeholt, J.A.L.; Driessen, A.J.M. The bacterial Sec-translocase: Structure and mechanism. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1016–1028. [Google Scholar] [CrossRef]
- Majlessi, L.; Prados-Rosales, R.; Casadevall, A.; Brosch, R. Release of mycobacterial antigens. Immunol. Rev. 2015, 264, 25–45. [Google Scholar] [CrossRef]
- O’Brien, K.D.; Pineda, C.; Chiu, W.S.; Bowen, R.; Deeg, M.A. Glycosylphosphatidylinositol-Specific Phospholipase D Is Expressed by Macrophages in Human Atherosclerosis and Colocalizes With Oxidation Epitopes. Circulation 1999, 99, 2876–2882. [Google Scholar] [CrossRef]
- Wolf, B.; Freehauf, C.L.; Thomas, J.A.; Gordon, P.L.; Greene, C.L.; Ward, J.C. Markedly elevated serum biotinidase activity may indicate glycogen storage disease type Ia. J. Inherit. Metab. Dis. 2003, 26, 805–809. [Google Scholar] [CrossRef] [PubMed]
- Fatima, I.; Sadaf, S.; Musharraf, S.G.; Hashmi, N.; Akhtar, M.W. CD5 molecule-like and transthyretin as putative biomarkers of chronic myeloid leukemia - an insight from the proteomic analysis of human plasma. Sci. Rep. 2017, 7, 40943. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Yan, X.; Zhang, Q.; Yin, Y.; Cao, J. CD5L contributes to the pathogenesis of methicillin-resistant Staphylococcus aureus-induced pneumonia. Int. Immunopharmacol. 2019, 72, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Aran, G.; Sanjurjo, L.; Barcena, C.; Simon-Coma, M.; Téllez, É.; Vázquez-Vitali, M.; Garrido, M.; Guerra, L.; Díaz, E.; Ojanguren, I.; et al. CD5L is upregulated in hepatocellular carcinoma and promotes liver cancer cell proliferation and antiapoptotic responses by binding to HSPA5 (GRP78). FASEB J. 2018, 32, 3878–3891. [Google Scholar] [CrossRef] [PubMed]
- Huaman, M.A.; Henson, D.; Ticona, E.; Sterling, T.R.; Garvy, B.A. Tuberculosis and cardiovascular disease: Linking the epidemics. Trop. Dis. Travel Med. Vaccines 2015, 1, 10. [Google Scholar] [CrossRef] [PubMed]
- Wayne, L.; Chandler, M.D. Chronic Elevated Levels of Factor VIII and Other Coagulation Factors. In Transfusion Medicine and Hemostasis, 3rd ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2019; pp. 907–908. [Google Scholar]
- Guan, Y.; Hou, X.; Tian, P.; Ren, L.; Tang, Y.; Song, A.; Zhao, J.; Gao, L.; Song, G. Elevated Levels of Apolipoprotein CIII Increase the Risk of Postprandial Hypertriglyceridemia. Front. Endocrinol. 2021, 12, 646185. [Google Scholar] [CrossRef]
- Skjeflo, E.W.; Brækkan, S.K.; Ludviksen, J.K.; Snir, O.; Hindberg, K.; Mollnes, T.E.; Hansen, J.-B. Elevated plasma concentration of complement factor C5 is associated with risk of future venous thromboembolism. Blood 2021, 138, 2129–2137. [Google Scholar] [CrossRef]
- Szaluś-Jordanow, O.; Augustynowicz-Kopeć, E.; Czopowicz, M.; Olkowski, A.; Łobaczewski, A.; Rzewuska, M.; Sapierzyński, R.; Wiatr, E.; Garncarz, M.; Frymus, T. Intracardiac tuberculomas caused by Mycobacterium tuberculosis in a dog. BMC Veter- Res. 2016, 12, 1–6. [Google Scholar] [CrossRef]
- Huaman, M.A.; Qualls, J.E.; Jose, S.; Schmidt, S.M.; Moussa, A.; Kuhel, D.G.; Konaniah, E.; Komaravolu, R.K.; Fichtenbaum, C.J.; Deepe, G.S.J.; et al. Mycobacterium bovis Bacille-Calmette-Guérin Infection Aggravates Atherosclerosis. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- López-López, J.P.; Posada-Martínez, E.L.; Saldarriaga, C.; Wyss, F.; Ponte-Negretti, C.I.; Alexander, B.; Miranda-Arboleda, A.F.; Martínez-Sellés, M.; Baranchuk, A.; the Neglected Tropical Diseases; et al. Tuberculosis and the Heart. J. Am. Heart Assoc. 2021, 10, e019435. [Google Scholar] [CrossRef]
- Basham, C.; Smith, S.J.; Romanowski, K.; Johnston, J.C. Cardiovascular morbidity and mortality among persons diagnosed with tuberculosis: A systematic review and meta-analysis. PLoS ONE 2020, 15, e0235821. [Google Scholar] [CrossRef] [PubMed]
- Reichelt, J.; Breiden, B.; Sandhoff, K.; Magin, T.M. Loss of keratin 10 is accompanied by increased sebocyte proliferation and differentiation. Eur. J. Cell Biol. 2004, 83, 747–759. [Google Scholar] [CrossRef] [PubMed]
- Mattii, M.; Lovászi, M.; Garzorz, N.; Atenhan, A.; Quaranta, M.; Lauffer, F.; Konstantinow, A.; Küpper, M.; Zouboulis, C.; Kemény, L.; et al. Sebocytes contribute to skin inflammation by promoting the differentiation of T helper 17 cells. Br. J. Dermatol. 2017, 178, 722–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, S.-Y.; Lai, Y.-R.; Kung, C.-T.; Tsai, N.-W.; Su, C.-M.; Huang, C.-C.; Wang, H.-C.; Cheng, B.-C.; Su, Y.-J.; Lin, W.-C.; et al. α-1-Acid Glycoprotein Concentration as an Outcome Predictor in Adult Patients with Sepsis. BioMed Res. Int. 2019, 2019, 1–9. [Google Scholar] [CrossRef]
- Miravitlles, M.; Herepath, M.; Priyendu, A.; Sharma, S.; Vilchez, T.; Vit, O.; Haensel, M.; Lepage, V.; Gens, H.; Greulich, T. Disease burden associated with alpha-1 antitrypsin deficiency: Systematic and structured literature reviews. Eur. Respir. Rev. 2022, 31, 210262. [Google Scholar] [CrossRef]
- Liz, M.A.; Coelho, T.; Bellotti, V.; Fernandez-Arias, M.I.; Mallaina, P.; Obici, L. A Narrative Review of the Role of Transthyretin in Health and Disease. Neurol. Ther. 2020, 9, 395–402. [Google Scholar] [CrossRef]
- de la Fuente, J. Translational biotechnology for the control of ticks and tick-borne diseases. Ticks Tick. Borne Dis. 2021, 12, 101738. [Google Scholar] [CrossRef]
- Abegglen, L.M.; Caulin, A.F.; Chan, A.; Lee, K.; Robinson, R.; Campbell, M.S.; Kiso, W.K.; Schmitt, D.L.; Waddell, P.J.; Bhaskara, S.; et al. Potential Mechanisms for Cancer Resistance in Elephants and Comparative Cellular Response to DNA Damage in Humans. JAMA 2015, 314, 1850–1860. [Google Scholar] [CrossRef]
- Seluanov, A.; Gladyshev, V.N.; Vijg, J.; Gorbunova, V. Mechanisms of cancer resistance in long-lived mammals. Nat. Cancer 2018, 18, 433–441. [Google Scholar] [CrossRef]
- Contreras, M.; Artigas-Jerónimo, S.; Comín, J.J.P.; de la Fuente, J. A Quantum Vaccinomics Approach Based on Protein–Protein Interactions. Vaccine Des. 2021, 287–305. [Google Scholar] [CrossRef]
- de la Fuente, J.; Contreras, M. Additional considerations for ant-tick vaccine research. Expert Rev Vaccines 2022, 3, 1–3. [Google Scholar] [CrossRef]
- Davenne, T.; McShane, H. Why don’t we have an effective tuberculosis vaccine yet? Expert Rev. Vaccines 2016, 15, 1009–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villar, M.; Rajbhandari, R.M.; Artigas-Jerónimo, S.; Contreras, M.; Sadaula, A.; Karmacharya, D.; Alves, P.C.; Gortázar, C.; de la Fuente, J. Differentially Represented Proteins in Response to Infection with Mycobacterium tuberculosis Identified by Quantitative Serum Proteomics in Asian Elephants. Pathogens 2022, 11, 1010. https://doi.org/10.3390/pathogens11091010
Villar M, Rajbhandari RM, Artigas-Jerónimo S, Contreras M, Sadaula A, Karmacharya D, Alves PC, Gortázar C, de la Fuente J. Differentially Represented Proteins in Response to Infection with Mycobacterium tuberculosis Identified by Quantitative Serum Proteomics in Asian Elephants. Pathogens. 2022; 11(9):1010. https://doi.org/10.3390/pathogens11091010
Chicago/Turabian StyleVillar, Margarita, Rajesh Man Rajbhandari, Sara Artigas-Jerónimo, Marinela Contreras, Amir Sadaula, Dibesh Karmacharya, Paulo Célio Alves, Christian Gortázar, and José de la Fuente. 2022. "Differentially Represented Proteins in Response to Infection with Mycobacterium tuberculosis Identified by Quantitative Serum Proteomics in Asian Elephants" Pathogens 11, no. 9: 1010. https://doi.org/10.3390/pathogens11091010