
Impact of the Cultivation System and Plant Cultivar on Arbuscular Mycorrhizal Fungi of Spelt (Triticum aestivum ssp. Spelta L.) in a Short-Term Monoculture

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Number of AMF Spores

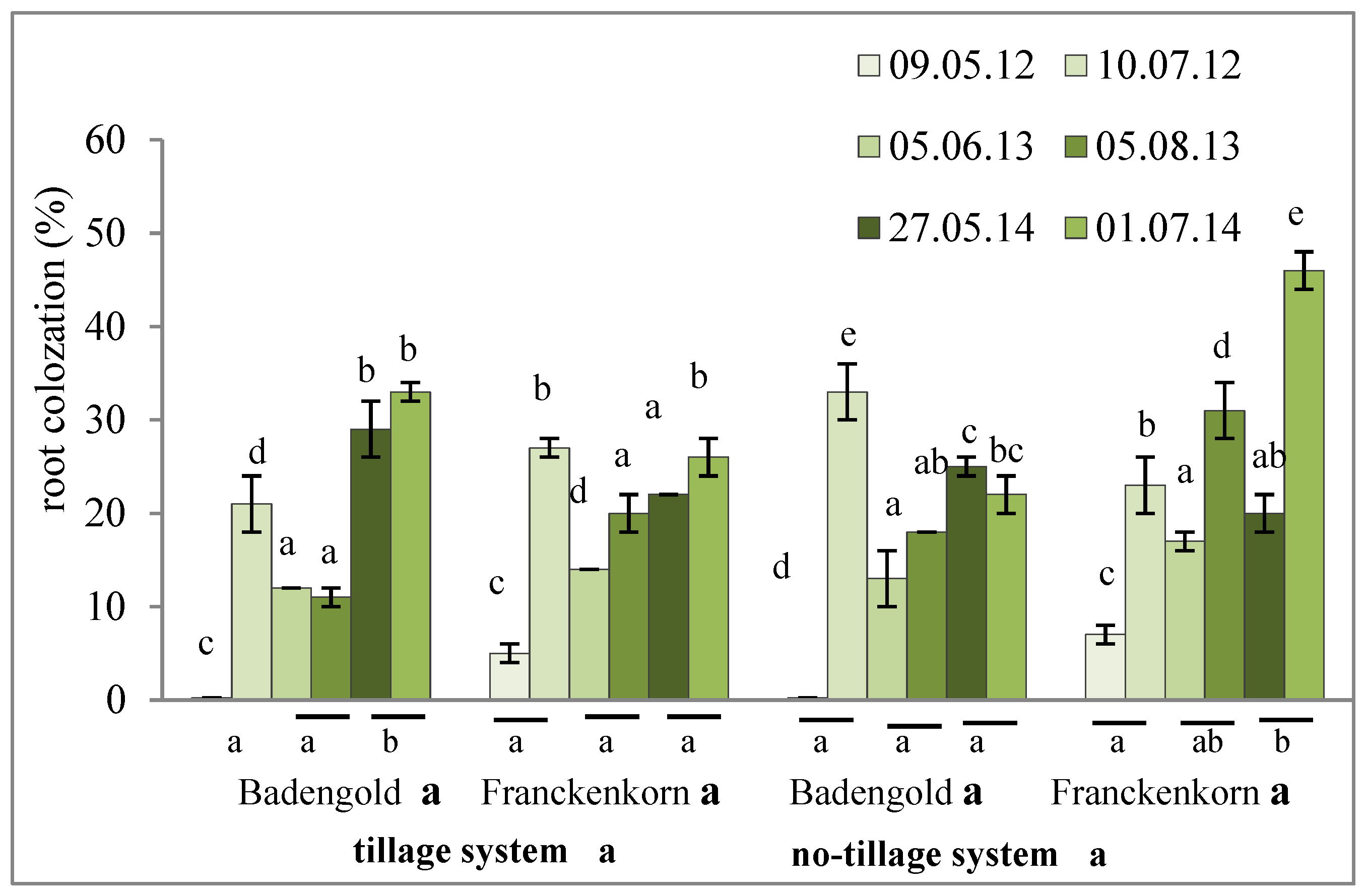
2.2. Degree of Endomycorrhizal Colonization

2.3. Relationships between the Total Number of Spores and AMF Root Colonization (Correlation Analysis) in Relation to Sampling Dates (PCA Analysis)

3. Discussion
4. Materials and Methods
4.1. Characteristics of the Experimental Object
4.2. Meteorological Conditions

4.3. Preparation of Research Material
4.4. Determination of the Number of AMF Spores
4.5. Determination of the Degree of Root Colonization by AMF
4.6. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P. The rhizosphere microbione and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; Ver Loren van Themaat, E. Schulze-Lefert Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [Green Version]
- Tester, M.; Smith, S.E.; Smith, F.A. The phenomenon of non-mycorrhizal plants. Can. J. Bot. 1987, 65, 419–431. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 2nd ed.; Academic Press: San Diego, CA, USA, 1997; p. 605. [Google Scholar]
- Schüßler, A.; Schwarzott, D.; Walker, C. A new phylum the Glomeromycota: Phylogeny and evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- Harrier, L.A.; Watson, C.A. The role of arbuscular mycorrhizal fungi in sustainable cropping systems. Adv. Agron. 2003, 79, 185–225. [Google Scholar]
- Read, D.J.; Perez-Moreno, Z. Mycorrhizas and nutrient cycling in ecosystems a journey towards relevance? New Phytol. 2003, 137, 475–492. [Google Scholar] [CrossRef]
- Hayman, D.S. The physiology of vesicular-arbuscular endomycorrhizal symbiosis. Can. J. Bot. 1983, 61, 944–963. [Google Scholar] [CrossRef]
- Johansen, A.; Finlay, R.D.; Olsson, P.A. Nitrogen metabolism of external hyphae of the arbuscular mycorrhizal fungus Glomus intraradices. New Phytol. 1996, 133, 705–712. [Google Scholar] [CrossRef]
- Harrier, L.A.; Watson, C.A. The potential role of arbuscular mycorrhizal (AM) fungi of plants against soil-borne pathogenes in organic and/or other sustainable farming systems. Pest. Manag. Sci. 2004, 60, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Veresoglou, S.D.; Chen, B.; Rilling, M.C. Arbuscular mycorrhiza and soil nitrogen cycling. Soil Biol. Biochem. 2012, 46, 53–62. [Google Scholar] [CrossRef]
- Hodge, A. Plant nitrogen capture from organic matter as affected by spatial dispersion interspecific competition and mycorrhizal colonization. New Phytol. 2003, 157, 303–314. [Google Scholar] [CrossRef]
- Jamiołowska, A.; Księżniak, A.; Hetman, B.; Kopacki, M.; Skwaryło-Bednarz, B.; Gałązka, A.; Thanoon, A.H. Interactions of arbuscular mycorrhizal fungi with plants and soil microflora. Acta Sci. Pol. Hortorum Cultus 2017, 16, 89–95. [Google Scholar] [CrossRef]
- Whipps, J.M. Prospects and limitations for mycorrhizas in biocontrol of root pathogens. Can. J. Bot. 2004, 82, 1198–1227. [Google Scholar] [CrossRef]
- Lee, B.-R.; Muneer, S.; Avice, J.-C.; Jyng, W.-L.; Kim, T.-H. Mycorrhizal colonization and P-supplement effects on N uptake and N assimilation in perennial ryegrass under well-watered and drought-stressed conditions. Mycorrhiza 2012, 22, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Johansson, J.F.; Paul, L.R.; Finlay, R.D. Microbial interactions in the mycorrhizosphere and their significance for sustainable agriculture. FEMS Microbiol. Ecol. 2004, 48, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.-H.; Yao, Q.; Sun, X.-T.; Hu, Y.-L. Colonization, ALP activity and plant growth promotion of native and exotic arbuscular mycorrhizal fungi at low pH. Soil Biol. Biochem. 2007, 39, 942–950. [Google Scholar] [CrossRef]
- Minz, D.; Ofek, M. Rhizosphere Microorganisms in Multicellular Life Forms; Rosenberg, E., Gophna, U., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 105–122. [Google Scholar]
- Yang, Y.; Han, X.; Liang, Y.; Ghosh, A.; Chen, J.; Tang, M. The combined effects of arbuscular mycorrhizal fungi (AMF) and lead (Pb) stress on Pb accumulation plant growth parameters, photosynthesis and antioxidant enzymes in Robinia pseudoacacia L. PLoS ONE 2015, 10, e0145726. [Google Scholar] [CrossRef]
- Rogers, J.B.; Laidlaw, A.S.; Christie, P. The role of arbuscular mycorrhizal fungi in the transfer of nutrients between white clover and perennial ryegrass. Chemosphere 2001, 42, 153–159. [Google Scholar] [CrossRef]
- Hart, M.M.; Reader, R.J.; Klironomos, J.N. Plant coexistence mediated by arbuscular mycorrhizal fungi. Trends Biochem. Sci. 2003, 18, 418–423. [Google Scholar] [CrossRef]
- Hodge, A. Microbial ecology of the arbuscular mycorrhiza. FEMS Microbiol. Ecol. 2000, 32, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Thanoon, R.H.; Jamiołkowska, A. Effect of mycorrhiza on the diversity and structure of the population of saprotrophic fungi occurring in the rhizosphere of tomato plants. Ann. Hortic. 2018, 28, 15–24. Available online: http://polona.pl/item/144274984 (accessed on 23 March 2022). [CrossRef]
- Roger-Estrade, J.; Anger, C.; Bertrand, M.; Guy, R. Tillage and soil ecology: Partners for sustainable agriculture. Soil Tillage Res. 2010, 11, 33–40. [Google Scholar] [CrossRef]
- Jansa, J.; Gryndler, M. Biotic environment of the arbuscular mycorrhizal fungi in soil. In Arbuscular Mycorrhizas: Physiology and Function; Koltai, H., Kapulnik, Y., Eds.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010; pp. 209–236. [Google Scholar]
- Jamiołowska, A.; Księżniak, A.; Gałązka, A.; Hetman, B.; Kopacki, M.; Skwaryło-Bednarz, B. Impact of abiotic factors on development of the community of arbuscular mycorrhizal fungi in the soil: A review. Int. Agrophys. 2018, 32, 133–140. [Google Scholar] [CrossRef]
- Allen, E.B.; Allen, M.F.; Helni, D.J.; Trappe, J.M.; Molina, R.; Rincon, E. Patterns and regulation of mycorrhizal plant and fungal diversity. Plant Soil 1995, 170, 47–62. [Google Scholar] [CrossRef]
- Schalamuk, S.; Velazqueaz, S.; Chidichimo, H.; Cabello, M. Fungal spore diversity of arbuscular mycorrhizal fungi associated with spring wheat: Effect of tillage. Mycologia 2006, 98, 22–28. [Google Scholar] [CrossRef]
- Code-Menum, B.J.; Berch, S.M.; Bomke, A.A. Seasonal colonization of arbuscular mycorrhizal fungi in temperate grasslands along a wide hydrologie gradient. Mycorrhiza 1991, 15, 291–299. [Google Scholar]
- Matsubara, Y.; Harada, T. Effect of arbuscular mycorrhizal fungus infection and growth and mineral nutrient content of Asparagus officinolis L. seedlings. J. Jpn. Soc. Hortic. Sci. 1996, 65, 303–309. [Google Scholar] [CrossRef]
- Księżniak, A.; Kobus, J. Dynamics of endomycorrhizal colonization of cereal roots in different arable soils. In Microorganisms in the Environment. Occurrence, Activity, Importance; Barabasz, W., Ed.; Publishing House of the Agricultural University: Kraków, Poland, 1997; pp. 315–319. (In Polish) [Google Scholar]
- Sauza-Andrade, D.; Collazzi-Filho, A.; Giller, K.E. The soil microbial community and soil tillage. In Soil Tillage in Agroecosystems; El Titi, A., Ed.; CRC Press: Boca Raton, FL, USA, 2003; pp. 267–296. [Google Scholar]
- Usuki, K.; Yamamoto, H.; Tazawa, J. Effect of previous cropping and tillage system on growth of maize and symbiotic association with arbuscular mycorrhizal fungi in central region of Japan. Jpn. J. Crop Sci. 2007, 76, 394–400. [Google Scholar] [CrossRef]
- Sivakumar, N. Effect of edafic factors and seasonal variation on spore density and root colonization of arbuscular mycorrhizal fungi in sugarcane fields. Ann. Microb. 2013, 61, 151–160. [Google Scholar] [CrossRef]
- Muthukumar, T.; Udaiyan, K. Seasonality of vesicular-arbuscular mycorrhizae in sedoes in a semi-arid tropical grassland. Acta Oecologica 2002, 23, 337–347. [Google Scholar] [CrossRef]
- Valsalakumar, N.; Ray, J.G.; Potty, V.R. Arbuscular mycorrhizal fungi associated with green gram in South India. Agron. J. 2007, 99, 1260–1267. [Google Scholar] [CrossRef]
- Panwar, V.; Meghvansi, M.K.; Siddiqui, S. Short-term temporal variation in sporulation dynamics of arbuscular mycorrhizal (AM) fungi and physico-chemical edaphic properties of wheat rhizosphere. Saudi J. Biol. Sci. 2011, 18, 247–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasrullah Sharif, M.; Rubina, K.; Burni, T. Occurrence and distribution of arbuscular mycorrhizal fungi in wheat and maize crops of malakand division of north west frontier province. Pak. J. Bot. 2010, 42, 1301–1312. [Google Scholar]
- Khanam, D.; Mridha, M.A.U.; Solaiman, A.R.M.; Hossain, T. Effect of edaphic factors on root colonization and spore population of arbuscular mycorrhizal fungi. Bull. Inst. Trop. Agric. Kyushu Univ. 2006, 29, 97–104. [Google Scholar]
- Księżniak, A.; Kobus, J. Occurence of spores of VAM fungi and colonisation of roots of cropplants in various soils in Poland. In Ekologiczne Aspekty Mikrobiologii Gleby; Sawicka, A., Durska, G., Eds.; Publishing House of the Agricultural University: Poznań, Poland, 1998; pp. 149–154. (In Polish) [Google Scholar]
- Smith, S.E.; Manjarrez, M.; Stonor, R.; McNeill, A.; Smith, A. Indigenous arbuscular mycorrhizal (AM) fungi contribute to wheat phosphate uptake in a semi-arid fields environment show by tracking with radioactive phosphorus. Appl. Soil Ecol. 2015, 96, 68–74. [Google Scholar] [CrossRef]
- Kuźniar, A.; Włodarczyk, K.; Grządziel, J.; Goraj, W.; Gałązka, A.; Wolińska, A. Culture-independent analysis of an endophytic core in two species of wheat: Triticum aestivum L. (cv. ‘Holandia’) and the first report of microbiome in Triticum spelta L. (cv. ‘Rokosz’). Syst. Appl. Microbiol. 2020, 43, 126025. [Google Scholar] [CrossRef]
- Ratajczak, K.; Sulewska, H.; Błaszczyk, L.; Basińska-Barczak, A.; Mikołajczak, K.; Salamon, S.; Szymańska, G.; Dryjański, L. Growth and photosynthetic activity of selected spelt varieties (Triticum aestivum ssp. spelta L.) cultivated under drought conditions with different endophilic core mivrobiomes. Int. J. Mol. Sci. 2020, 21, 7987. [Google Scholar] [CrossRef]
- Salamon, S.; Mikołajczak, K.; Błaszczyk, L.; Ratajczak, K.; Sulewska, H. Changes in root-associated fungal communities in Triticum aestivum ssp. spelta L. and Triticum eastivum ssp. vulgare L. under drought stress and various soil processing. PLoS ONE 2020, 15, e0240037. [Google Scholar] [CrossRef]
- Korniłłowicz-Kowalska, T.; Andruszczak, S.; Bohacz, J.; Kraska, P.; Możejko, M.; Kwiecińska-Poppe, E. The effect of tillage and no-tillage system on culturable fungal communities in the rhizosphere and soil of two spelt cultivars. Appl. Soil Ecol. 2022, 174, 104413. [Google Scholar] [CrossRef]
- Iwaniuk, A.; Błaszkowski, J. Arbuscular fungi and mycorrhizae of agricultural soils of the western Pomerania, part I. Occurrence of arbuscular fungi and mycorrhizae. Acta Mycol. 2004, 39, 65–91. [Google Scholar] [CrossRef]
- Kowalczyk, S.; Błaszkowski, J. Arbuskularne grzyby mikoryzowe gleb województwa lubuskiego. Acta Agrobot. 2005, 58, 453–474. (In Polish) [Google Scholar] [CrossRef] [Green Version]
- Castillo, C.G.; Rubio, R.; Rouanet, J.L.; Borie, F. Early effect of tillage and crop rotation on arbuscular mycorrhizal fungal propagules in an Ultisol. Biol. Fertil. Soils 2006, 43, 83–92. [Google Scholar] [CrossRef]
- Kabir, Z. Tillage or non-tillage: Impact on mycorrhizae. Can. J. Plant Sci. 2005, 85, 23–29. Available online: https://cdnsciencepub.com/doi/pdf/10.4141/P03-160 (accessed on 18 February 2022). [CrossRef] [Green Version]
- Sheng, M.; Lalande, E.; Hamel, C.; Ziadi, N. Effect of long-term tillage and mineral phosphorus fertilization on arbuscular mycorrhizal fungi in a humid continental zone of Eastern Canada. Plant Sci. 2013, 369, 599–613. [Google Scholar] [CrossRef]
- Koide, R.T.; Schreiner, R.P. Regulation of the vesicular-arbuscular mycorrhizal symbiosis. Annu. Rev. Plant Biol. 1992, 43, 557–581. [Google Scholar] [CrossRef]
- Błaszkowski, J. Comparative studies of the occurence of arbuskular fungi and mycorrhizae (Glomales) in cultivated and uncultivated soils of Poland. Acta Mycol. 1993, 28, 93–140. [Google Scholar] [CrossRef] [Green Version]
- Gosling, P.; Ozaki, A.; Jones, J.; Turner, M.; Rayns, F.; Bending, G.D. Organic management of tilled agricultural soils results in rapid increase in colonization potential and spore populations of arbuscular mycorrhizal fungi. Agric. Ecosyst. Environ. 2010, 139, 273–279. [Google Scholar] [CrossRef]
- Jansa, J.; Oberholzer, H.-R.; Egli, S. Environmental determinants of the arbuscular mycorrhizal fungal infectivity of Swiss agricultural soils. Eur. J. Soil Biol. 2009, 45, 400–408. [Google Scholar] [CrossRef]
- Wu, S.C.; Cao, Z.H.; Li, Z.G.; Cheung, K.C.; Wong, M.H. Effects of biofertilizer containing N-fixer P and K solubilizers and AM fungi on maize growth: A greenhouse trial. Geoderma 2005, 12, 155–166. [Google Scholar] [CrossRef]
- Mobasser, H.R.; Moradgholi, A.; Mehrabon, A.; Koohkan, S. Investigation of mycorrhizal effect on agronomic trains and protein procent of corn varieties in sista. Int. J. Agric. Sci. 2012, 2, 108–119. [Google Scholar]
- Andruszczak, S.; Kwiecińska-Poppe, E.; Kraska, P.; Pałys, E. The influence of some plant protection chemical means on leaf area and their tip angle of some winter spelt wheat varieties (Triticum aestivum ssp. spelta L.). Prog. Plant Prot. 2012, 52, 163–166. [Google Scholar]
- Iannucci, A.; Fragasso, M.; Bellegia, R.; Nigro, F.; Papa, R. Evolution of the crop rhizosphere: Impact of domestication on roat exudates in tetraploid wheat (Triticum turgidum L.). Front. Plant Sci. 2017, 8, 2124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korniłłowicz-Kowalska, T.; Wojdyło-Kotwica, B.; Bohacz, J.; Możejko, M.T. Biodiversity of saprotrophic of the root zone of grasses and dovers in a permanent meadow in a post-bog habital in relations to the growing season and fertilization (Zaklęsłość sosnowicka, Western Polesye). In Environmental Engineering in Polesyr, Book 3 Polysye; Urban, D., Dobrowolski, R., Jeznach, J., Eds.; International Scientific Publications Brest-Rivne-Warsaw-Ryazan, Belarus-Ukraine-Poland-Russia; Warsaw Uniwersity of Life Sciences Publishing House: Warsaw, Poland, 2020; pp. 485–538. (In Polish) [Google Scholar]
- Panwar, J.; Tarafdar, J.C. Arbuscular mycorrhizal fungal dynamics under Mitragyna parvifolia (Roxb.) Korth. in Thar Desert. Appl. Soil Ecol. 2006, 34, 200–208. [Google Scholar] [CrossRef]
- Rodríguez-Echeverría, S.; Gera Hol, W.H.; Freitas, H.; Eason, W.R.; Cook, R. Arbuscular mycorrhizal fungi of Ammophila arenaria (L.) Link: Spore abundance and root colonization in six location of the European coast. Eur. J. Soil Biol. 2008, 44, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Li, L.F.; Yang, A.N.; Zhao, Z.-W. Seasonality of arbuscular mycorrhizal symbiosis and dark septate endophytes in a grassland site in southwest China. FEMS Microbiol. Ecol. 2005, 54, 367–373. [Google Scholar] [CrossRef]
- Titus, J.H.; Leps, J. The response of arbuscular mycorrhizae to fertilization, mowing and removal of dominant species in a diverse oligotrophic wet meadow. Am. J. Bot. 2010, 87, 392–401. [Google Scholar] [CrossRef] [Green Version]
- IUSS Working Group WRB. World Reference Base of Soil Resources 2006, 2nd ed.; World Soil Recourses Reports No. 103; FAO: Rome, Italy, 2006. [Google Scholar]
- Andruszczak, S. Reaction of winter spelt cultivars to reduced tillage system and chemical plant protection. Zemdirbyste-Agriculture 2017, 104, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Okoń, S.; Paczos-Grzęda, E.; Kraska, P.; Kwiecińska-Poppe, E.; Pałys, E. Assessment of genetic similarity of spelt (Triticum aestivum ssp. spelta L.) cultivars based on RAPD markers. Biul. Inst. Hod. Aklim. Rośl. 2009, 252, 35–41. [Google Scholar]
- Andruszczak, S.; Kraska, P.; Kwiecińska-Poppe, E.; Skowrońska, M. Cultivar and foliar feeding of plants as factors determining the chemical composition of spelt (Triticum aestivum ssp. spelta L.) grain. Appl. Ecol. Environ. Res. 2020, 18, 2949–2958. [Google Scholar] [CrossRef]
- Kraska, P.; Andruszczak, S.; Kwiecińska-Poppe, E.; Pałys, E. Effect of chemical crop protection on the content of some elements in grain of spelt wheat (Triticum aestivum ssp. spelta). J. Elem. 2013, 18, 79–90. [Google Scholar] [CrossRef]
- Andruszczak, S.; Kraska, P.; Kwiecińska-Poppe, E.; Pałys, E. The effect of tillage system and herbicide application on weed infestation of crops of winter spelt wheat (Triticum aestivum ssp. spelta) cultivars. Acta Agrobot. 2013, 66, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Allen, M.; Moore, T.; Christensen, M.; Stanton, N. Growth of vesicular-arbuscular-mycorrhizal and nonmycorrhizal Bouteloua gracilis in a defined medium. Mycologia 1979, 71, 666–669. [Google Scholar] [CrossRef]
- Philips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Kormanik, P.P.; McGraw, A.C. Quantification of vesicular-arbuscular mycorrhizae in plant roots. In Methods and Principles of Mycorrhizal Research; Schenk, N.C., Ed.; American Phytopathological Society: St. Paul, MN, USA, 1982; pp. 37–45. [Google Scholar]
- Omar, M.B.; Bolland, L.; Heather, W.A. A permanent mounting medium for fungi. Bull. Br. Mycol. Soc. 1979, 19, 337–341. [Google Scholar] [CrossRef]
- Koske, R.E.; Tessler, B. A convenient permanent slide mounting medium. Mycol. Soc. Am. Newsl. 1983, 34, 59. [Google Scholar]
- Nilsson, M.; Andreas, A.; Lagerkvist, A. Effect of accelerated carbonation and zero valent iron on metal leaching from bottom ash. Waste Manag. 2016, 51, 97–104. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | Tillage System | No-Tillage System | ||||
|---|---|---|---|---|---|---|
| Cultivar Badengold | Cultivar Franckenkorn | Means for Both Varieties | Cultivar Badengold | Cultivar Franckenkorn | Means for Both Varieties | |
| 2012 | ||||||
| 9.05. | 96.08 (736) * | 90.70 (1186) | 92.70 (961) | 87.69 (577) | 89.78 (1169) | 89.08 (873) |
| 10.07. | 92.18 (1277) | 90.60 (868) | 91.51 (1073) | 91.96 (927) | 99.87 (995) | 92.94 (961) |
| Means 2012 | 93.60 (1007) | 90.7 (10.37) | 92.0 (1017) | 90.28 (752) | 91.61 (1082) | 91.06 (917) |
| 2013 | ||||||
| 05.06. | 96.25 (1103) | 90.83 (1050) | 93.54 (1077) | 92.07 (1022) | 86.90 1333) | 89.11 (1178) |
| 05.08. | 95.89 (1589) | 89.35 (1033) | 93.20 (1310) | 97.97 (1209) | 87.26 (1740) | 91.38 (1475) |
| Means 2013 | 96.07 (1345) | 90.13 (1042) | 93.40 (1194) | 95.22 (1116) | 87.13 1537) | 90.39 (1327) |
| 2014 | ||||||
| 27.05. | 98.33 (1244) | 96.55 (1008) | 97.49 (1126) | 90.03 (1536) | 96.80 (1812) | 93.57 (1674) |
| 01.07. | 86.04 (962) | 89.46 (1376) | 88.02 (1169) | 82.36 (1695) | 83.42 (2022) | 82.95 (1859) |
| Means 2014 | 92.53 (1103) | 92.33 (1192) | 92.43 (1148) | 85.87 (1616) | 90.16 (1917) | 87.82 (1767) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bohacz, J.; Korniłłowicz-Kowalska, T.; Rybczyńska-Tkaczyk, K.; Andruszczak, S. Impact of the Cultivation System and Plant Cultivar on Arbuscular Mycorrhizal Fungi of Spelt (Triticum aestivum ssp. Spelta L.) in a Short-Term Monoculture. Pathogens 2022, 11, 844. https://doi.org/10.3390/pathogens11080844
Bohacz J, Korniłłowicz-Kowalska T, Rybczyńska-Tkaczyk K, Andruszczak S. Impact of the Cultivation System and Plant Cultivar on Arbuscular Mycorrhizal Fungi of Spelt (Triticum aestivum ssp. Spelta L.) in a Short-Term Monoculture. Pathogens. 2022; 11(8):844. https://doi.org/10.3390/pathogens11080844
Chicago/Turabian StyleBohacz, Justyna, Teresa Korniłłowicz-Kowalska, Kamila Rybczyńska-Tkaczyk, and Sylwia Andruszczak. 2022. "Impact of the Cultivation System and Plant Cultivar on Arbuscular Mycorrhizal Fungi of Spelt (Triticum aestivum ssp. Spelta L.) in a Short-Term Monoculture" Pathogens 11, no. 8: 844. https://doi.org/10.3390/pathogens11080844