
Evidence of Virulent Multi-Drug Resistant and Biofilm-Forming Listeria Species Isolated from Various Sources in South Africa

 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
2. Results
2.1. Detection of Listeria spp.
2.2. Antibiotic Resistance
2.2.1. Antibiotic Resistant Profiles of L. monocytogenes Strains
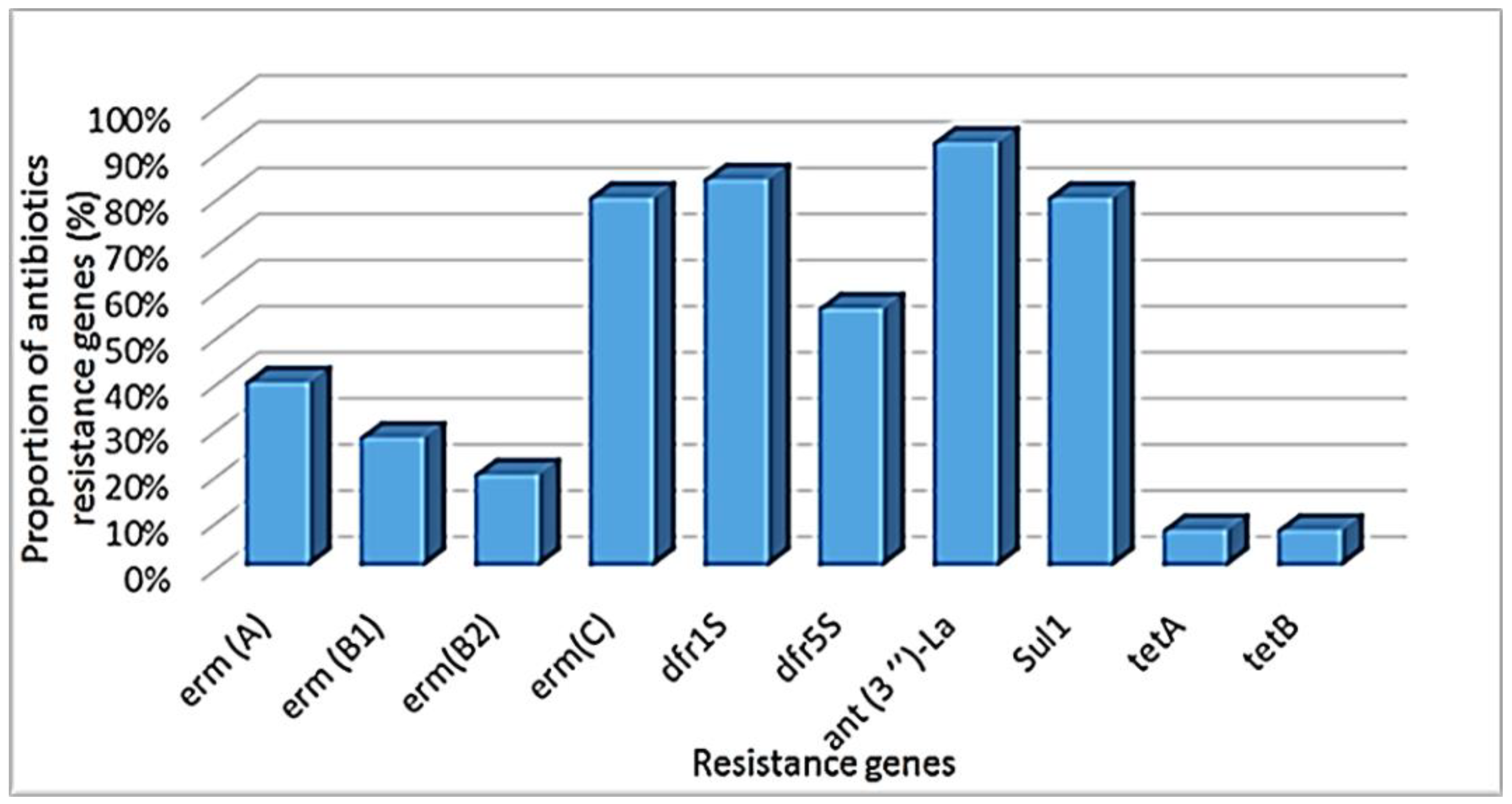
2.2.2. Antibiotic Resistance Genes
2.3. Proportion of Virulence Genes Detection in L. monocytogenes
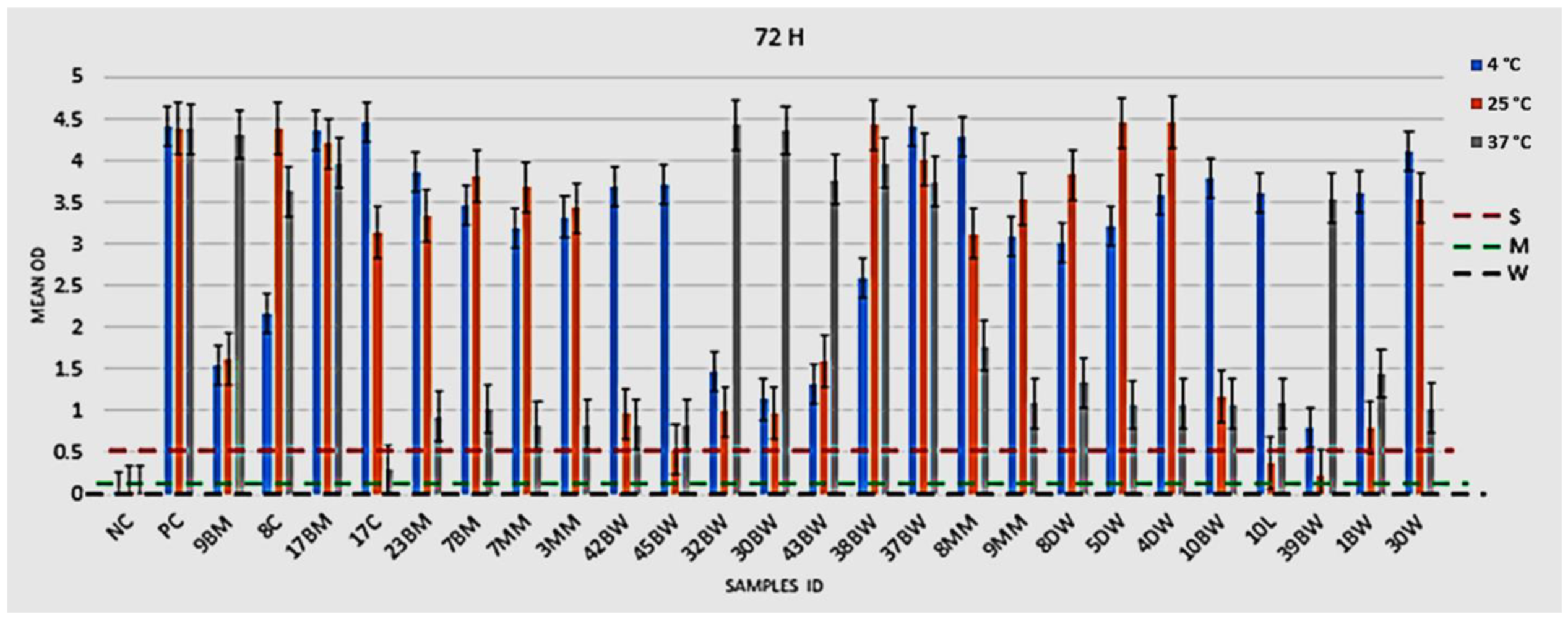
2.4. Biofilm-Forming Potential of L. monocytogenes Isolates
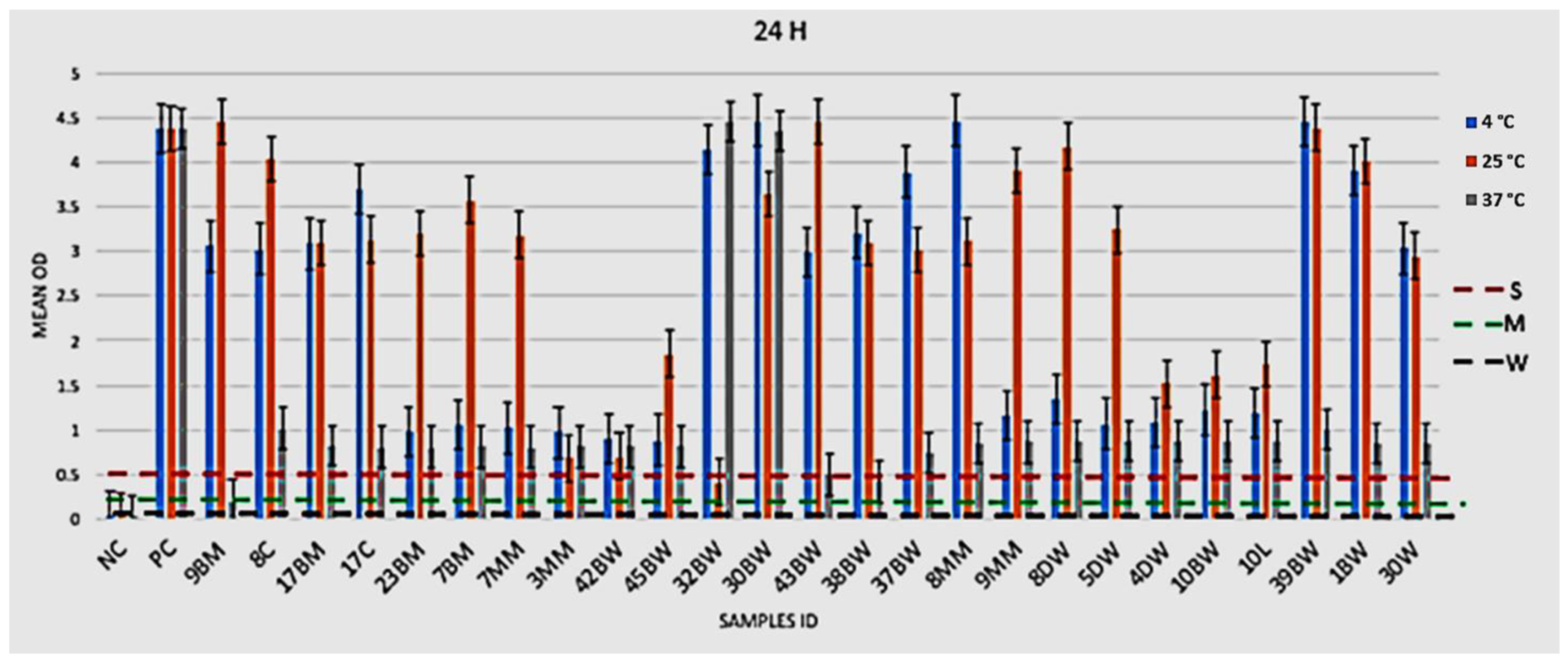
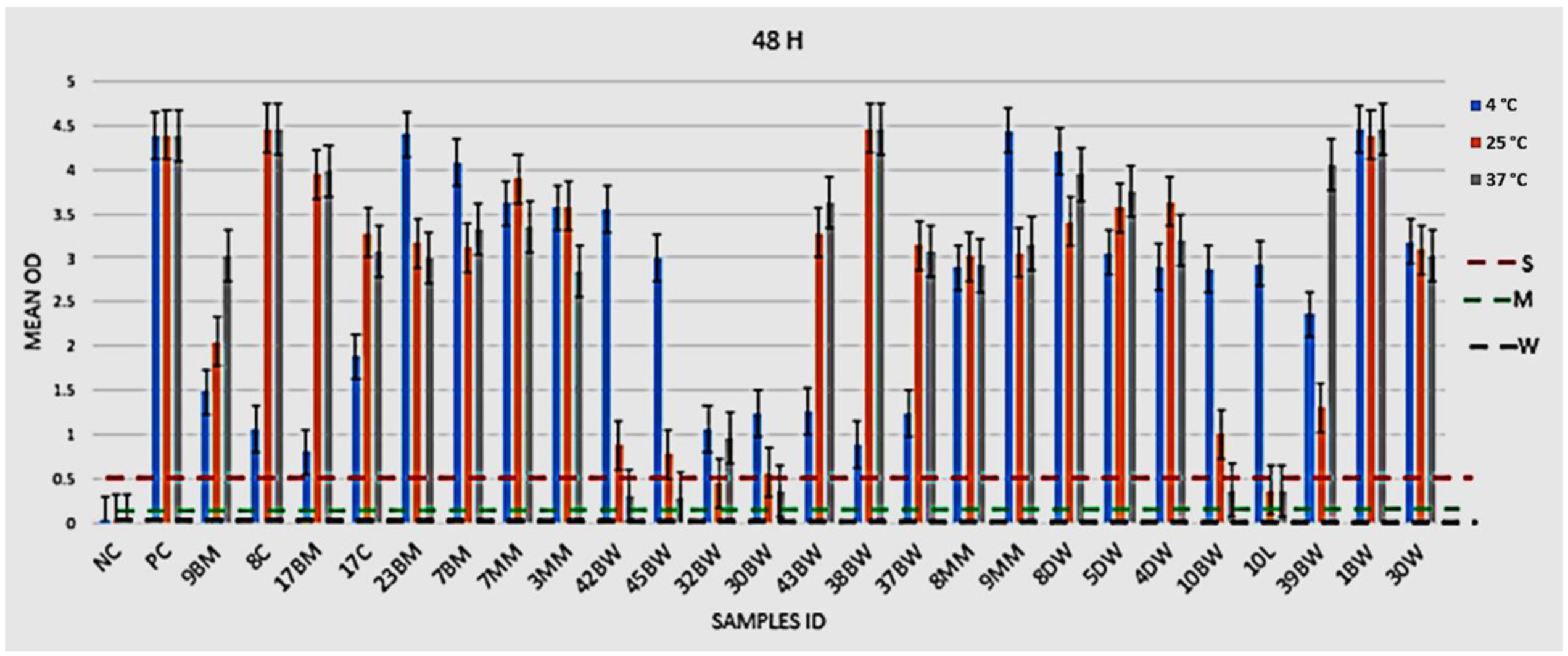
2.4.1. Phenotypic Assessment of Biofilm Formation
2.4.2. Biofilm Formation Genes
3. Discussion
4. Materials and Methods
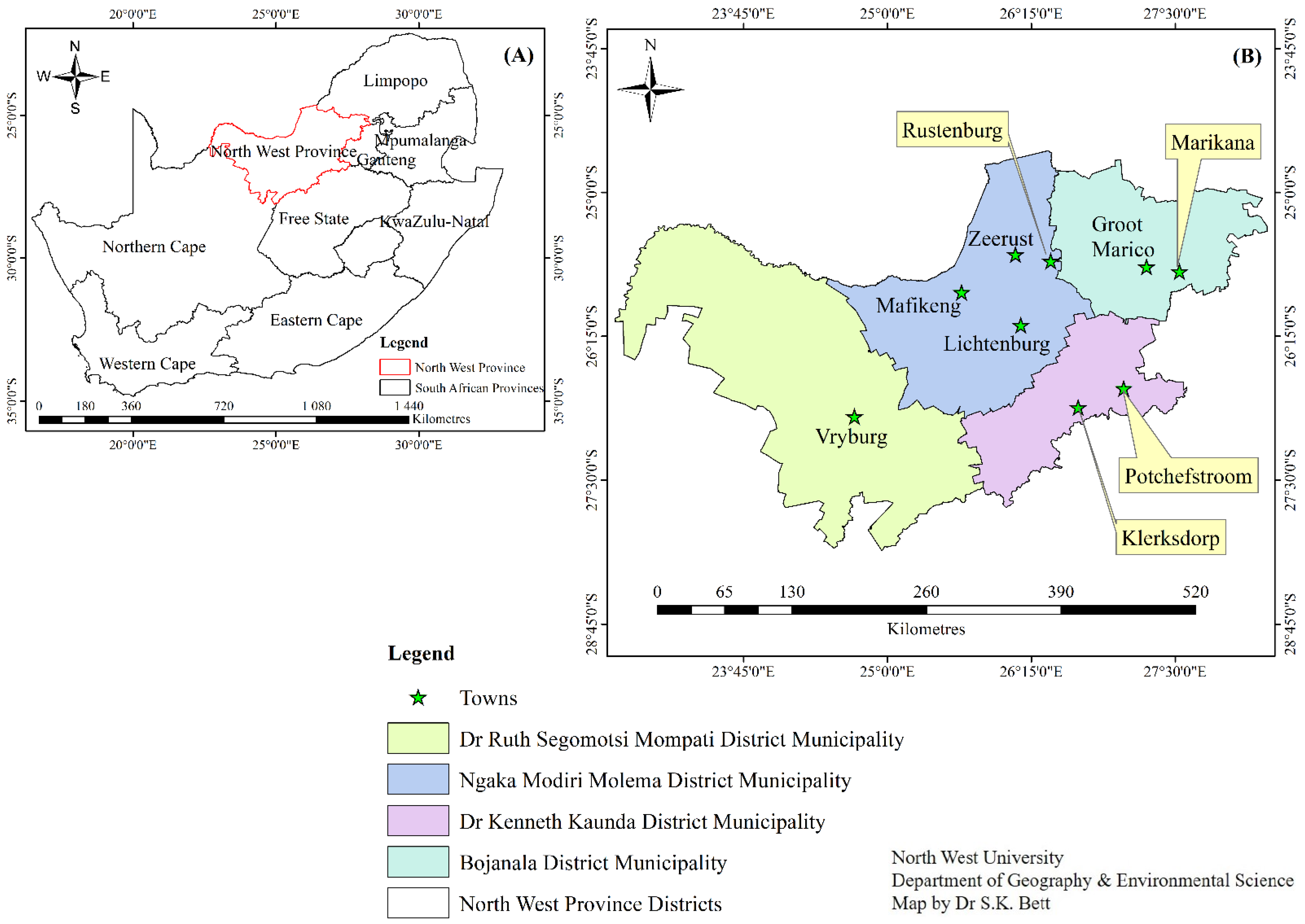
4.1. Sample Area and Sample Collection
4.2. Bacterial Strains
4.3. Isolation of Listeria spp.
4.4. Molecular Characterizations of Listeria Species Using PCR Analysis
4.4.1. Extraction of Chromosomal DNA
4.4.2. Identification of Listeria spp., Virulence Genes, Antibiotic Resistance Genes and Biofilm Genes Using PCR (Polymerase Chain Reaction) Technique
4.5. Antimicrobial Susceptibility Test
4.6. Biofilm Formation
Phenotypic Detection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- McLauchlin, J.; Mitchell, R.; Smerdon, W.; Jewell, K. Listeria monocytogenes and listeriosis: A review of hazard characterisation for use in microbiological risk assessment of foods. Int. J. Food Microbiol. 2004, 92, 15–33. [Google Scholar] [CrossRef]
- Troxler, R.; von Graevenitz, A.; Funke, G.; Wiedemann, B.; Stock, I. Natural antibiotic susceptibility of Listeria species: L. grayi, L. innocua, L. ivanovii, L. monocytogenes, L. seeligeri and L. welshimeri strains. Clin. Microbiol. Infect. 2000, 6, 525–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Møretrø, T.; Langsrud, S. Listeria monocytogenes: Biofilm formation and persistence in food-processing environments. Biofilms 2004, 1, 107–121. [Google Scholar] [CrossRef]
- Liu, D. Identification, subtyping and virulence determination of Listeria monocytogenes, an important foodborne pathogen. J. Med Microbiol. 2006, 55, 645–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, D.J.V.; Cossart, P. Recent advances in understanding Listeria monocytogenes infection: The importance of subcellular and physiological context. F1000Research 2017, 6, 1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linscott, A.J. Food-Borne Illnesses. Clin. Microbiol. Newsl. 2011, 33, 41–45. [Google Scholar] [CrossRef]
- Wang, Z.; Tao, X.; Liu, S.; Zhao, Y.; Yang, X. An Update Review on Listeria Infection in Pregnancy. Infect. Drug Resist. 2021, 14, 1967–1978. [Google Scholar] [CrossRef]
- Mateus, T.; Silva, J.; Maia, R.; Teixeira, P. Listeriosis during Pregnancy: A Public Health Concern. ISRN Obstet. Gynecol. 2013, 2013, 851712. [Google Scholar] [CrossRef] [Green Version]
- Kitada, T.; Kadoba, K.; Watanabe, R.; Koyama, T.; Nakayama, Y.; Taki, M.; Yukawa, S.; Odani, K.; Morinobu, A. Listeriosis presenting with fever, arthralgia, elevated liver enzymes, and hyperferritinaemia in pregnancy: A critical mimicker of adult-onset Still’s disease. Scand. J. Rheumatol. 2021, 51, 78–80. [Google Scholar] [CrossRef]
- Ntuli, N.; Wadula, J.; Nakwa, F.; Thomas, R.; Van Kwawegen, A.; Sepeng, L.; Seake, K.; Kgwadi, D.; Sono, L.; Ondongo-Ezhet, C.; et al. Characteristics and Outcomes of Neonates With Blood Stream Infection Due to Listeria monocytogenes. Pediatr. Infect. Dis. J. 2021, 40, 917–921. [Google Scholar] [CrossRef]
- Allen, K.J.; Wałecka-Zacharska, E.; Chen, J.C.; Katarzyna, K.-P.; Devlieghere, F.; Van Meervenne, E.; Osek, J.; Wieczorek, K.; Bania, J. Listeria monocytogenes—An examination of food chain factors potentially contributing to antimicrobial resistance. Food Microbiol. 2016, 54, 178–189. [Google Scholar] [CrossRef]
- Authority, E.F.S.; Prevention, E.C.F.D. Control, The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2014. EFSA J. 2015, 13, 4329. [Google Scholar]
- Marshall, B.M.; Levy, S.B. Food Animals and Antimicrobials: Impacts on Human Health. Clin. Microbiol. Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef] [Green Version]
- Kawacka, I.; Olejnik-Schmidt, A.; Schmidt, M.; Sip, A. Effectiveness of Phage-Based Inhibition of Listeria monocytogenes in Food Products and Food Processing Environments. Microorganisms 2020, 8, 1764. [Google Scholar] [CrossRef]
- Tchatchouang, C.-D.K.; Fri, J.; De Santi, M.; Brandi, G.; Schiavano, G.F.; Amagliani, G.; Ateba, C.N. Listeriosis Outbreak in South Africa: A Comparative Analysis with Previously Reported Cases Worldwide. Microorganisms 2020, 8, 135. [Google Scholar] [CrossRef] [Green Version]
- Munshi, M.; Sharma, M.; Deb, S. Viability of High-Pressure Technology in the Food Industry. In Handbook of Research on Food Processing and Preservation Technologies: Emerging Techniques for Food Processing, Quality, and Safety Assurance; Apple Academic Press: Palm Bay, FL, USA, 2021; Volume 5, p. 51. [Google Scholar]
- Patel, D.; Rathod, R. Ready-to-eat food perception, food preferences and food choice—A theoretical discussion. Int. J. Multidiscip. Res. Dev. 2017, 3, 198–205. [Google Scholar]
- Chaudhury, R. Determinants of consumer behavior in buying RTE foods. J. Bus. Retail. Manag. Res. 2010, 5, 76–86. [Google Scholar]
- Food Safety News. Company Recalls Imported Mushrooms after State Test Finds Listeria Monocytogenes. News Desk on 20 March 2022. Available online: https://www.foodsafetynews.com/2022/03/company-recalls-imported-mushrooms-after-state-test-finds-Listeria-monocytogenes/ (accessed on 4 April 2022).
- Food Drug Administration. Banner Smoked Fish Expands Recalls Smoked Fish Products, Salads, Pickled Fish Products, and Cream Sauce Products Because of Possible Health Risk. Company Announcement Date: 3 June 2021. Available online: https://www.fda.gov/safety/recalls-market-withdrawals-safety-alerts/banner-smoked-fish-expands-recalls-smoked-fish-products-salads-pickled-fish-products-and-cream-sauce (accessed on 7 April 2022).
- Food Drug Administration. Outbreak Investigation of Listeria Monocytogenes: Enoki Mushrooms (March 2020). Available online: https://www.fda.gov/food/outbreaks-foodborne-illness/outbreak-investigation-Listeria-monocytogenes-enoki-mushrooms-march-2020 (accessed on 7 April 2022).
- US FDA Guidance for Industry. Estimating the Maximum Safe Starting Dose in Initial Clinical Trials for Therapeutics in Adult Healthy Volunteers. Pharmacology and Toxicology. 2005. Available online: http://www.fda.gov/cder/guidance/index.htm (accessed on 25 April 2017).
- Olanya, O.M.; Hoshide, A.K.; Ijabadeniyi, O.A.; Ukuku, D.O.; Mukhopadhyay, S.; Niemira, B.A.; Ayeni, O. Cost estimation of listeriosis (Listeria monocytogenes) occurrence in South Africa in 2017 and its food safety implications. Food Control 2019, 102, 231–239. [Google Scholar] [CrossRef]
- Jones, E.M.; MacGowan, A.P. Antimicrobial chemotherapy of human infection due to Listeria monocytogenes. Eur. J. Clin. Microbiol. 1995, 14, 165–175. [Google Scholar] [CrossRef]
- Walsh, D.; Duffy, G.; Sheridan, J.; Blair, I.; McDowell, D. Antibiotic resistance among Listeria, including Listeria monocytogenes, in retail foods. J. Appl. Microbiol. 2001, 90, 517–522. [Google Scholar] [CrossRef]
- Shamloo, E.; Hosseini, H.; Moghadam, Z.A.; Larsen, M.H.; Haslberger, A.; Alebouyeh, M. Importance of Listeria monocyto-genes in food safety: A review of its prevalence, detection, and antibiotic resistance. Iran. J. Vet. Res. 2019, 20, 241. [Google Scholar]
- Borucki, M.K.; Peppin, J.D.; White, D.; Loge, F.; Call, D.R. Variation in Biofilm Formation among Strains of Listeriamonocytogenes. Appl. Environ. Microbiol. 2003, 69, 7336–7342. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-López, P.; Rodríguez-Herrera, J.J.; Vázquez-Sánchez, D.; Cabo, M.L. Current Knowledge on Listeria monocytogenes Biofilms in Food-Related Environments: Incidence, Resistance to Biocides, Ecology and Biocontrol. Foods 2018, 7, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennani, H.; Mateus, A.; Mays, N.; Eastmure, E.; Stärk, K.D.C.; Häsler, B. Overview of Evidence of Antimicrobial Use and Antimicrobial Resistance in the Food Chain. Antibiotics 2020, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.A.; Chandry, P.S.; Kaur, M.; Kocharunchitt, C.; Bowman, J.P.; Fox, E.M. Characterisation of Listeria monocytogenes food-associated isolates to assess environmental fitness and virulence potential. Int. J. Food Microbiol. 2021, 350, 109247. [Google Scholar] [CrossRef]
- Jordan, K.; Hunt, K.; Lourenco, A.; Pennone, V. Listeria monocytogenes in the Food Processing Environment. Curr. Clin. Microbiol. Rep. 2018, 5, 106–119. [Google Scholar] [CrossRef]
- Ferri, M.; Ranucci, E.; Romagnoli, P.; Giaccone, V. Antimicrobial resistance: A global emerging threat to public health systems. Crit. Rev. Food Sci. Nutr. 2015, 57, 2857–2876. [Google Scholar] [CrossRef]
- El Salabi, A.; Walsh, T.; Chouchani, C. Extended spectrum β-lactamases, carbapenemases and mobile genetic elements responsible for antibiotics resistance in Gram-negative bacteria. Crit. Rev. Microbiol. 2012, 39, 113–122. [Google Scholar] [CrossRef]
- Beceiro, A.; Tomás, M.; Bou, G. Antimicrobial Resistance and Virulence: A Successful or Deleterious Association in the Bacterial World? Clin. Microbiol. Rev. 2013, 26, 185–230. [Google Scholar] [CrossRef] [Green Version]
- Manganyi, M.; Regnier, T.; Olivier, E. Antimicrobial activities of selected essential oils against Fusarium oxysporum isolates and their biofilms. S. Afr. J. Bot. 2015, 99, 115–121. [Google Scholar] [CrossRef]
- Gonçalves, I.R.; Dantas, R.C.C.; Ferreira, M.L.; Batistão, D.W.D.F.; Gontijo-Filho, P.P.; Ribas, R.M. Carbapenem-resistant Pseudomonas aeruginosa: Association with virulence genes and biofilm formation. Braz. J. Microbiol. 2016, 48, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Velge, P.; Bottreau, E.; Van-Langendonck, N.; Kaeffer, B. Cell proliferation enhances entry of Listeria monocytogenes into intestinal epithelial cells by two proliferation-dependent entry pathways. J. Med. Microbiol. 1997, 46, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Cossart, P. Invasion of mammalian cells by Listeria monocytogenes: Functional mimicry to subvert cellular functions. Trends Cell Biol. 2003, 13, 23–31. [Google Scholar] [CrossRef]
- Chatterjee, S.S.; Hossain, H.; Otten, S.; Kuenne, C.; Kuchmina, K.; Machata, S.; Domann, E.; Chakraborty, T.; Hain, T. Intracellular Gene Expression Profile of Listeria monocytogenes. Infect. Immun. 2006, 74, 1171–1180. [Google Scholar] [CrossRef] [Green Version]
- Kreft, J.; Vázquez-Boland, J.A. Regulation of virulence genes in Listeria. Int. J. Med Microbiol. 2001, 291, 145–157. [Google Scholar] [CrossRef]
- Volokhov, D.V.; Duperrier, S.; Neverov, A.A.; George, J.; Buchrieser, C.; Hitchins, A.D. The Presence of the Internalin Gene in Natural Atypically Hemolytic Listeria innocua Strains Suggests Descent from L. monocytogenes. Appl. Environ. Microbiol. 2007, 73, 1928–1939. [Google Scholar] [CrossRef] [Green Version]
- De las Heras, A.; Cain, R.J.; Bielecka, M.K.; Vazquez-Boland, J.A. Regulation of Listeria virulence: PrfA master and commander. Curr. Opin. Microbiol. 2011, 14, 118–127. [Google Scholar] [CrossRef]
- Quereda, J.J.; Morón-García, A.; Palacios-Gorba, C.; Dessaux, C.; Portillo, F.G.-D.; Pucciarelli, M.G.; Ortega, A.D. Pathogenicity and virulence of Listeria monocytogenes: A trip from environmental to medical microbiology. Virulence 2021, 12, 2509–2545. [Google Scholar] [CrossRef]
- Vázquez-Boland, J.A.; Domínguez-Bernal, G.; González-Zorn, B.; Kreft, J.; Goebel, W. Pathogenicity islands and virulence evolution in Listeria. Microbes Infect. 2001, 3, 571–584. [Google Scholar] [CrossRef]
- Schiavano, G.F.; Ateba, C.N.; Petruzzelli, A.; Mele, V.; Amagliani, G.; Guidi, F.; De Santi, M.; Pomilio, F.; Blasi, G.; Gattuso, A.; et al. Whole-Genome Sequencing Characterization of Virulence Profiles of Listeria monocytogenes Food and Human Isolates and In Vitro Adhesion/Invasion Assessment. Microorganisms 2021, 10, 62. [Google Scholar] [CrossRef]
- Mukherjee, K.; Raju, R.; Fischer, R.; Vilcinskas, A. Galleria Mellonella as a Model Host to Study Gut Microbe Homeostasis and Brain Infection by the Human Pathogen Listeria Monocytogenes. Yellow Biotechnol. 2013, 135, 27–39. [Google Scholar] [CrossRef]
- Jian, Z.; Zeng, L.; Xu, T.; Sun, S.; Yan, S.; Yang, L.; Huang, Y.; Jia, J.; Dou, T. Antibiotic resistance genes in bacteria: Occurrence, spread, and control. J. Basic Microbiol. 2021, 61, 1049–1070. [Google Scholar] [CrossRef]
- EFSA; ECDC. European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018, 16, e05500. [Google Scholar] [CrossRef]
- Muchaamba, F.; Eshwar, A.K.; Stevens, M.J.A.; von Ah, U.; Tasara, T. Variable Carbon Source Utilization, Stress Resistance, and Virulence Profiles Among Listeria monocytogenes Strains Responsible for Listeriosis Outbreaks in Switzerland. Front. Microbiol. 2019, 10, 957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matle, I.; Mbatha, K.R.; Lentsoane, O.; Magwedere, K.; Morey, L.; Madoroba, E. Occurrence, serotypes, and characteristics of Listeria monocytogenes in meat and meat products in South Africa between 2014 and 2016. J. Food Saf. 2019, 39, e12629. [Google Scholar] [CrossRef]
- Matle, I.; Mafuna, T.; Madoroba, E.; Mbatha, K.R.; Magwedere, K.; Pierneef, R. Population Structure of Non-ST6 Listeria monocytogenes Isolated in the Red Meat and Poultry Value Chain in South Africa. Microorganisms 2020, 8, 1152. [Google Scholar] [CrossRef]
- Rip, D.; Gouws, P.A. PCR–Restriction Fragment Length Polymorphism and Pulsed-Field Gel Electrophoresis Characterization of Listeria monocytogenes Isolates from Ready-to-Eat Foods, the Food Processing Environment, and Clinical Samples in South Africa. J. Food Prot. 2020, 83, 518–533. [Google Scholar] [CrossRef]
- Parichanon, P.; Sattayakhom, A.; Matan, N.; Matan, N. Antimicrobial activity of lime oil in the vapour phase against Listeria monocytogenes on ready-to-eat salad during cold storage and its possible mode of action. Food Control 2022, 132, 108486. [Google Scholar] [CrossRef]
- Makumbe, H.; Tabit, F.; Dlamini, B. Prevalence, Molecular Identification, Antimicrobial Resistance, and Disinfectant Susceptibility of Listeria innocua Isolated from Ready-to-Eat Foods Sold in Johannesburg, South Africa. J. Food Qual. Hazards Control 2021, 8, 131–139. [Google Scholar] [CrossRef]
- Keet, R.; Rip, D. Listeria monocytogenes isolates from Western Cape, South Africa exhibit resistance to multiple antibiotics and contradicts certain global resistance patterns. AIMS Microbiol. 2021, 7, 40–58. [Google Scholar] [CrossRef]
- Archer, D.L. The evolution of FDA’s policy on Listeria monocytogenes in ready-to-eat foods in the United States. Curr. Opin. Food Sci. 2018, 20, 64–68. [Google Scholar] [CrossRef]
- Thomas, J.; Govender, N.; McCarthy, K.M.; Erasmus, L.K.; Doyle, T.J.; Allam, M.; Ismail, A.; Ramalwa, N.; Sekwadi, P.; Ntshoe, G.; et al. Outbreak of Listeriosis in South Africa Associated with Processed Meat. N. Engl. J. Med. 2020, 382, 632–643. [Google Scholar] [CrossRef]
- Mazzotta, A.S.; Gombas, D.E. Heat Resistance of an Outbreak Strain of Listeria monocytogenes in Hot Dog Batter. J. Food Prot. 2001, 64, 321–324. [Google Scholar] [CrossRef]
- WHO. Listeriosis–Spain [Internet]. Cited 7 August 2020. Available online: https://www.who.int/csr/don/16-september-2019-listeriosis-spain/en/ (accessed on 4 May 2022).
- Gelbíčová, T.; Zobaníková, M.; Tomáštíková, Z.; Van Walle, I.; Ruppitsch, W.; Karpíšková, R. An outbreak of listeriosis linked to turkey meat products in the Czech Republic, 2012–2016. Epidemiol. Infect. 2018, 146, 1407–1412. [Google Scholar] [CrossRef] [Green Version]
- Pappelbaum, K.; Grif, K.; Heller, I.; Würzner, R.; Hein, I.; Ellerbroek, L.; Wagner, M. Monitoring Hygiene On- and At-Line Is Critical for Controlling Listeria monocytogenes during Produce Processing. J. Food Prot. 2008, 71, 735–741. [Google Scholar] [CrossRef]
- Alvarez-Ordóñez, A.; Leong, D.; Hunt, K.; Scollard, J.; Butler, F.; Jordan, K. Production of safer food by understanding risk factors for L. monocytogenes occurrence and persistence in food processing environments. J. Food Saf. 2018, 38, e12516. [Google Scholar] [CrossRef]
- Leong, D.; NicAogáin, K.; Luque-Sastre, L.; McManamon, O.; Hunt, K.; Alvarez-Ordóñez, A.; Scollard, J.; Schmalenberger, A.; Fanning, S.; O’Byrne, C.; et al. A 3-year multi-food study of the presence and persistence of Listeria monocytogenes in 54 small food businesses in Ireland. Int. J. Food Microbiol. 2017, 249, 18–26. [Google Scholar] [CrossRef]
- Viswanath, P.; Murugesan, L.; Knabel, S.J.; Verghese, B.; Chikthimmah, N.; Laborde, L.F. Incidence of Listeria monocytogenes and Listeria spp. in a Small-Scale Mushroom Production Facility. J. Food Prot. 2013, 76, 608–615. [Google Scholar] [CrossRef]
- Gianfranceschi, M.; Gattuso, A.; Tartaro, S.; Aureli, P. Incidence of Listeria monocytogenes in food and environmental samples in Italy between 1990 and 1999: Serotype distribution in food, environmental and clinical samples. Eur. J. Epidemiol. 2002, 18, 1001–1006. [Google Scholar] [CrossRef]
- Grif, K.; Patscheider, G.; Dierich, M.P.; Allerberger, F. Incidence of Fecal Carriage of Listeria monocytogenes in Three Healthy Volunteers: A One-Year Prospective Stool Survey. Eur. J. Clin. Microbiol. 2003, 22, 16–20. [Google Scholar] [CrossRef]
- Castro, H.; Jaakkonen, A.; Hakkinen, M.; Korkeala, H.; Lindström, M. Occurrence, Persistence, and Contamination Routes of Listeria monocytogenes Genotypes on Three Finnish Dairy Cattle Farms: A Longitudinal Study. Appl. Environ. Microbiol. 2018, 84, e02000-17. [Google Scholar] [CrossRef] [Green Version]
- Odjadjare, E.E.O.; Okoh, A.I. Prevalence and distribution of Listeria pathogens in the final effluents of a rural wastewater treatment facility in the Eastern Cape Province of South Africa. World J. Microbiol. Biotechnol. 2009, 26, 297–307. [Google Scholar] [CrossRef]
- Enurah, L.U.; Aboaba, O.O.; Nwachukwu, S.C.U.; Nwosuh, C.I. Antibiotic resistant profiles of food (fresh raw milk) and en-vironmental (abattoir effluents) isolates of Listeria monocytogenes from the six zones of Nigeria. Appl. Microbiol. Biotechnol. 2019, 2, 124–132. [Google Scholar] [CrossRef]
- Heidarzadeh, S.; Pourmand, M.R.; Hasanvand, S.; Pirjani, R.; Afshar, D.; Noori, M.; Dallal, M.M.S. Antimicrobial Susceptibility, Serotyping, and Molecular Characterization of Antibiotic Resistance Genes in Listeria monocytogenes Isolated from Pregnant Women with a History of Abortion. Iran. J. Public Health 2021, 50, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Morvan, A.; Moubareck, C.; Leclercq, A.; Hervé-Bazin, M.; Bremont, S.; Lecuit, M.; Courvalin, P.; Le Monnier, A. Antimicrobial Resistance of Listeria monocytogenes Strains Isolated from Humans in France. Antimicrob. Agents Chemother. 2010, 54, 2728–2731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makhubalo, K.; Manganyi, M.; Kumar, A.; Mbewe, M.; Ateba, C.N. Phenotypic and Genetic Characterization of Sorbitol-Fermenting Escherichia coli O157: H7 Isolated from Retail Beef and Mince Beef. J. Hum. Ecol. 2016, 56, 20–30. [Google Scholar] [CrossRef]
- Tshitshi, L.; Manganyi, M.C.; Montso, P.K.; Mbewe, M.; Ateba, C.N. Extended Spectrum Beta-Lactamase-Resistant Determinants among Carbapenem-Resistant Enterobacteriaceae from Beef Cattle in the North West Province, South Africa: A Critical Assessment of Their Possible Public Health Implications. Antibiotics 2020, 9, 820. [Google Scholar] [CrossRef] [PubMed]
- Du, X.-J.; Zhang, X.; Wang, X.-Y.; Su, Y.; Li, P.; Wang, S. Isolation and characterization of Listeria monocytogenes in Chinese food obtained from the central area of China. Food Control 2016, 74, 9–16. [Google Scholar] [CrossRef]
- Gaballa, A.; Oropeza, V.G.; Wiedmann, M.; Boor, K.J. Cross Talk between SigB and PrfA in Listeria monocytogenes Facilitates Transitions between Extra- and Intracellular Environments. Microbiol. Mol. Biol. Rev. 2019, 83, e00034-19. [Google Scholar] [CrossRef]
- Kanki, M.; Naruse, H.; Taguchi, M.; Kumeda, Y. Characterization of specific alleles in InlA and PrfA of Listeria monocytogenes isolated from foods in Osaka, Japan and their ability to invade Caco-2 cells. Int. J. Food Microbiol. 2015, 211, 18–22. [Google Scholar] [CrossRef]
- Soni, D.K.; Singh, R.K.; Singh, D.V.; Dubey, S.K. Characterization of Listeria monocytogenes isolated from Ganges water, human clinical and milk samples at Varanasi, India. Infect. Genet. Evol. 2012, 14, 83–91. [Google Scholar] [CrossRef]
- López, V.; Navas, J.; Martínez-Suárez, J.V. Low Potential Virulence Associated with Mutations in the inlA and prfA Genes in Listeria monocytogenes Isolated from Raw Retail Poultry Meat. J. Food Prot. 2013, 76, 129–132. [Google Scholar] [CrossRef]
- Ferreira, V.B.; Wiedmann, M.; Teixeira, P.; Stasiewicz, M.J. Listeria monocytogenes Persistence in Food-Associated Environments: Epidemiology, Strain Characteristics, and Implications for Public Health. J. Food Prot. 2014, 77, 150–170. [Google Scholar] [CrossRef]
- Costerton, J.W.; Geesey, G.G.; Cheng, K.-J. How Bacteria Stick. Sci. Am. 1978, 238, 86–95. [Google Scholar] [CrossRef]
- Yousef, A.; Lado, B. Characteristics of Listeria monocytogenes Important to Food Processors. In Listeria, Listeriosis and Food Safety, 3rd ed.; Ryser, E.T., Marth, E.H., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 157–213. [Google Scholar] [CrossRef]
- George, S.M.; Lund, B.M.; Brocklehurst, T. The effect of pH and temperature on initiation of growth of Listeria monocytogenes. Lett. Appl. Microbiol. 1988, 6, 153–156. [Google Scholar] [CrossRef]
- Lee, B.-H.; Cole, S.; Badel-Berchoux, S.; Guillier, L.; Felix, B.; Krezdorn, N.; Hébraud, M.; Bernardi, T.; Sultan, I.; Piveteau, P. Biofilm Formation of Listeria monocytogenes Strains Under Food Processing Environments and Pan-Genome-Wide Association Study. Front. Microbiol. 2019, 10, 2698. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Qiao, J.; Lu, Z.; Fen, Z.; Tao, Y.; Lv, F.; Zhao, H.; Zhang, C.; Bie, X. Influence of different factors on biofilm formation of Listeria monocytogenes and the regulation of cheY gene. Food Res. Int. 2020, 137, 109405. [Google Scholar] [CrossRef]
- Hassanien, A.; Shaker, E. Virulence Potential of Listeria monocytogenes Recovered from Ice cream and Aborted Women Samples in Sohag city, Egypt. Adv. Anim. Vet.-Sci. 2021, 9, 1829–1837. [Google Scholar] [CrossRef]
- Djordjevic, D.; Wiedmann, M.; McLandsborough, L.A. Microtiter Plate Assay for Assessment of Listeria monocytogenes Biofilm Formation. Appl. Environ. Microbiol. 2002, 68, 2950–2958. [Google Scholar] [CrossRef] [Green Version]
- Maje, M.D.; Tchatchouang, C.D.K.; Manganyi, M.C.; Fri, J.; Ateba, C.N. Characterisation of Vibrio Species from Surface and Drinking Water Sources and Assessment of Biocontrol Potentials of Their Bacteriophages. Int. J. Microbiol. 2020, 2020, 8863370. [Google Scholar] [CrossRef]
- Curiale, M.S.; Lewus, C. Detection of Listeria monocytogenes in Samples Containing Listeria innocua. J. Food Prot. 1994, 57, 1048–1051. [Google Scholar] [CrossRef]
- Silva, A.S.; Duarte, E.A.; De Oliveira, T.A.; Evangelista-Barreto, N.S. Identification of Listeria monocytogenes in cattle meat using biochemical methods and amplification of the hemolysin gene. Anais da Academia Brasileira de Ciências 2020, 92, e20180557. [Google Scholar] [CrossRef]
- Notermans, S.H.; Dufrenne, J.; Domann, E.; Chakraborty, T. Phosphatidylinositol-specific phospholipase C activity as a marker to distinguish between pathogenic and nonpathogenic Listeria species. Appl. Environ. Microbiol. 1991, 57, 2666–2670. [Google Scholar] [CrossRef] [Green Version]
- Paziak-Domańska, B.; Bogusławska, E.; Więckowska-Szakiel, M.; Kotłowski, R.; Różalska, B.; Chmiela, M.; Kur, J.; Dąbrowski, W.; Rudnicka, W. Evaluation of the API test, phosphatidylinositol-specific phospholipase C activity and PCR method in identification of Listeria monocytogenes in meat foods. FEMS Microbiol. Lett. 1999, 171, 209–214. [Google Scholar] [CrossRef]
- Furrer, B.; Candrian, U.; Hoefelein, C.; Luethy, J. Detection and identification of Listeria monocytogenes in cooked sausage products and in milk by in vitro amplification of haemolysin gene fragments. J. Appl. Bacteriol. 1991, 70, 372–379. [Google Scholar] [CrossRef]
- Beargie, R.A.; Bracken, E.C.; Riley, H.D. Micromethod (Spot-Plate) Determination of In Vitro Antibiotic Susceptibility. Appl. Microbiol. 1965, 13, 279–280. [Google Scholar] [CrossRef]
- Charpentier, E.; Courvalin, P. Antibiotic Resistance in Listeria spp. Antimicrob. Agents Chemother. 1999, 43, 2103–2108. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing. In CLSI Supplement M100, 27th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Bertrand, S.; Huys, G.; Yde, M.; D’Haene, K.; Tardy, F.; Vrints, M.; Swings, J.; Collard, J.-M. Detection and characterization of tet(M) in tetracycline-resistant Listeria strains from human and food-processing origins in Belgium and France. J. Med. Microbiol. 2005, 54, 1151–1156. [Google Scholar] [CrossRef]
- Mahm, M.M. Occurrence of Listeria monocytogenes in Raw Milk and Dairy Products in Noorabad, Iran. J. Anim. Vet.-Adv. 2010, 9, 16–19. [Google Scholar] [CrossRef] [Green Version]
- Ryu, J.; Park, S.H.; Yeom, Y.S.; Shrivastav, A.; Lee, S.-H.; Kim, Y.-R.; Kim, H.-Y. Simultaneous detection of Listeria species isolated from meat processed foods using multiplex PCR. Food Control 2013, 32, 659–664. [Google Scholar] [CrossRef]
- Coroneo, V.; Carraro, V.; Aissani, N.; Sanna, A.; Ruggeri, A.; Succa, S.; Meloni, B.; Pinna, A.; Sanna, C. Detection of Virulence Genes and Growth Potential in Listeria monocytogenes Strains Isolated from Ricotta Salata Cheese. J. Food Sci. 2015, 81, M114–M120. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Lawrence, M.L.; Austin, F.W.; Ainsworth, A.J. A multiplex PCR for species- and virulence-specific determination of Listeria monocytogenes. J. Microbiol. Methods 2007, 71, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Lomonaco, S.; Patti, R.; Knabel, S.J.; Civera, T. Detection of virulence-associated genes and epidemic clone markers in Listeria monocytogenes isolates from PDO Gorgonzola cheese. Int. J. Food Microbiol. 2012, 160, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Malik, S.; Vaidya, V.; Barbuddhe, S. Listeria monocytogenes in spontaneous abortions in humans and its detection by multiplex PCR. J. Appl. Microbiol. 2007, 103, 1889–1896. [Google Scholar] [CrossRef]
- Egervärn, M.; Roos, S.; Lindmark, H. Identification and characterization of antibiotic resistance genes in Lactobacillus reuteri and Lactobacillus plantarum. J. Appl. Microbiol. 2009, 107, 1658–1668. [Google Scholar] [CrossRef]
- Sutcliffe, J.; Grebe, T.; Tait-Kamradt, A.; Wondrack, L. Detection of erythromycin-resistant determinants by PCR. Antimicrob. Agents Chemother. 1996, 40, 2562–2566. [Google Scholar] [CrossRef] [Green Version]
- Delmani, F.; Jaran, A.S.; Tarazi, Y.A.; Masaadeh, H.; Zaki, O.; Irbid, T. Characterization of ampicillin resistant gene (blaTEM-1) isolated from E. coli in Northern Jordan. Asian J. Biomed. Pharm. Sci. 2017, 7, 11–15. [Google Scholar]
- Grape, M.; Motakefi, A.; Pavuluri, S.; Kahlmeter, G. Standard and real-time multiplex PCR methods for detection of trimethoprim resistance dfr genes in large collections of bacteria. Clin. Microbiol. Infect. 2007, 13, 1112–1118. [Google Scholar] [CrossRef] [Green Version]
- Bacci, C.; Boni, E.; Alpigiani, I.; Lanzoni, E.; Bonardi, S.; Brindani, F. Phenotypic and genotypic features of antibiotic resistance in Salmonella enterica isolated from chicken meat and chicken and quail carcasses. Int. J. Food Microbiol. 2012, 160, 16–23. [Google Scholar] [CrossRef]
- Poppe, C.; Martin, L.C.; Gyles, C.L.; Reid-Smith, R.; Boerlin, P.; McEwen, S.A.; Prescott, J.F.; Forward, K.R. Acquisition of Resistance to Extended-Spectrum Cephalosporins by Salmonella enterica subsp. enterica Serovar Newport and Escherichia coli in the Turkey Poult Intestinal Tract. Appl. Environ. Microbiol. 2005, 71, 1184–1192. [Google Scholar] [CrossRef] [Green Version]
- Warke, S.R.; Ingle, V.C.; Kurkure, N.V.; Tembhurne, P.A.; Prasad, M.; Chaudhari, S.P.; Barbuddhe, S.B. Biofilm Formation and Associated Genes in Listeria Monocytogenes. Indian J. Vet.-Sci. Biotechnol. 2017, 12, 7–12. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Source | Number Analyzed | Listeria spp. (%) | L. monocytogenes (%) |
|---|---|---|---|
| Vegetables | 8 | 3 (1.12) | 3 (5.66) |
| Meat/meat products | 108 | 33 (12.36) | 8 (15.09) |
| Water | 143 | 14 (5.24) | 14 (26.41) |
| Cattle feces | 8 | 3 (1.12) | 0 (0) |
| Isolate Source | Sampling Site | AP | TS | E | MEM | DA | GM | NOR | T | NOV | OX | NA | K | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MEAT/MEAT PRODUCTS | Meat | Lichtenburg n = 4 | 1 | 0 | 2 | 1 | 4 | 0 | 4 | 4 | 4 | 4 | 4 | 4 |
| Mince | Lichtenburg n = 2 | 0 | 1 | 2 | 1 | 2 | 2 | 1 | 2 | 2 | 2 | 2 | 2 | |
| Zeerust n = 2 | 1 | 1 | 2 | 1 | 2 | 2 | 0 | 2 | 2 | 2 | 2 | 2 | ||
| Total | N = 8 | 2(25) | 2(25) | 6(75) | 3(37) | 8(100) | 4(50) | 5(62.5) | 8(100) | 8(100) | 8(100) | 8(100) | 8(100) | |
| VEGETABLES | Cucumber | Zeerust n = 2 | 0 | 1 | 1 | 1 | 2 | 2 | 0 | 2 | 2 | 2 | 2 | 2 |
| Lettuces | Zeerust n = 1 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | |
| Total (%) | N = 3 | 0(0) | 1(33.33) | 2(66.67) | 1(33.33) | 3(100) | 3(100) | 0(0) | 3(100) | 3(100) | 3(100) | 3(100) | 3(100) | |
| WATER | Borehole | Lichtenburg n = 3 | 1 | 1 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Vryburg n = 3 | 0 | 1 | 2 | 2 | 3 | 3 | 0 | 3 | 3 | 3 | 3 | 3 | ||
| Mafikeng n = 5 | 1 | 4 | 3 | 0 | 5 | 5 | 0 | 5 | 5 | 5 | 5 | 1 | ||
| Dam | Mafikeng n = 2 | 0 | 0 | 2 | 0 | 2 | 2 | 0 | 2 | 2 | 2 | 2 | 2 | |
| Zeerust n = 1 | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | ||
| Total | N = 14 | 2(14.30) | 6(42.86) | 11(78.57) | 5(35.71) | 14(100) | 14(100) | 3(21.43) | 14(100) | 14(100) | 14(100) | 14(100) | 10(71.43) | |
| Resistance Pattern | MAR Phenotype | Number of Observed | MAR Index |
|---|---|---|---|
| I | E-DA-GM-T-NOV-OX-NA-K | 7 | 0.7 |
| II | MEM-DA-GM-T-NOV-OX-NA-K | 2 | 0.7 |
| III | AP-DA-GM-T-NOV-OX-NA-K | 2 | 0.7 |
| VI | TS-E-MEM-DA-GM-NOR-T-NOV-OX-NA-K | 1 | 0.8 |
| V | AP-TS-E-MEM-DA-GM-T-NOV-OX-NA-K | 1 | 0.9 |
| VI | TS-E-MEM-DA-GM-T-NOV-OX-NA-K | 1 | 0.8 |
| VII | DA-GM-T-NOV-OX-NA-K | 1 | 0.6 |
| VIII | AP-E-MEM-DA-GM-T-NOV-OX-NA-K | 1 | 0.8 |
| IX | E-MEM-DA-GM-T-NOV-OX-NA-K | 2 | 0.75 |
| X | TS-E-DA-GM-T-NOV-OX-NA-K | 2 | 0.75 |
| XI | TS-DA-GM-T-NOV-OX-NA-K | 1 | 0.7 |
| XII | TS-E-DA-GM-T-NOV-OX-NA | 1 | 0.7 |
| XIII | TS-E-DA-T-NOV-OX-NA-K | 1 | 0.7 |
| Incubation Temperature | Incubation Time (h) | Non Adherent (%) | Weak (%) | Moderate (%) | Strong (%) |
|---|---|---|---|---|---|
| 4 °C | 24 | 0 (0) | 0 (0) | 0 (0) | 25 (100) |
| 48 | 0 (0) | 0 (0) | 0 (0) | 25 (100) | |
| 72 | 0 (0) | 0 (0) | 0 (0) | 25 (100) | |
| 25 °C | 24 | 0 (0) | 0 (0) | 1 (4) | 24 (96) |
| 48 | 0 (0) | 0 (0) | 2 (8) | 23 (92) | |
| 72 | 0 (0) | 0 (0) | 2 (8) | 23 (92) | |
| 37 °C | 24 | 0 (0) | 0 (0) | 1 (4) | 24 (96) |
| 48 | 0 (0) | 0 (0) | 5 (20) | 20 (80) | |
| 72 | 0 (0) | 0 (0) | 1 (4) | 24 (96) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaptchouang Tchatchouang, C.-D.; Fri, J.; Montso, P.K.; Amagliani, G.; Schiavano, G.F.; Manganyi, M.C.; Baldelli, G.; Brandi, G.; Ateba, C.N. Evidence of Virulent Multi-Drug Resistant and Biofilm-Forming Listeria Species Isolated from Various Sources in South Africa. Pathogens 2022, 11, 843. https://doi.org/10.3390/pathogens11080843
Kaptchouang Tchatchouang C-D, Fri J, Montso PK, Amagliani G, Schiavano GF, Manganyi MC, Baldelli G, Brandi G, Ateba CN. Evidence of Virulent Multi-Drug Resistant and Biofilm-Forming Listeria Species Isolated from Various Sources in South Africa. Pathogens. 2022; 11(8):843. https://doi.org/10.3390/pathogens11080843
Chicago/Turabian StyleKaptchouang Tchatchouang, Christ-Donald, Justine Fri, Peter Kotsoana Montso, Giulia Amagliani, Giuditta Fiorella Schiavano, Madira Coutlyne Manganyi, Giulia Baldelli, Giorgio Brandi, and Collins Njie Ateba. 2022. "Evidence of Virulent Multi-Drug Resistant and Biofilm-Forming Listeria Species Isolated from Various Sources in South Africa" Pathogens 11, no. 8: 843. https://doi.org/10.3390/pathogens11080843