
Bendiocarb and Malathion Resistance in Two Major Malaria Vector Populations in Cameroon Is Associated with High Frequency of the G119S Mutation (Ace-1) and Overexpression of Detoxification Genes

 , , ,
, , ,
Abstract
:1. Background
2. Results
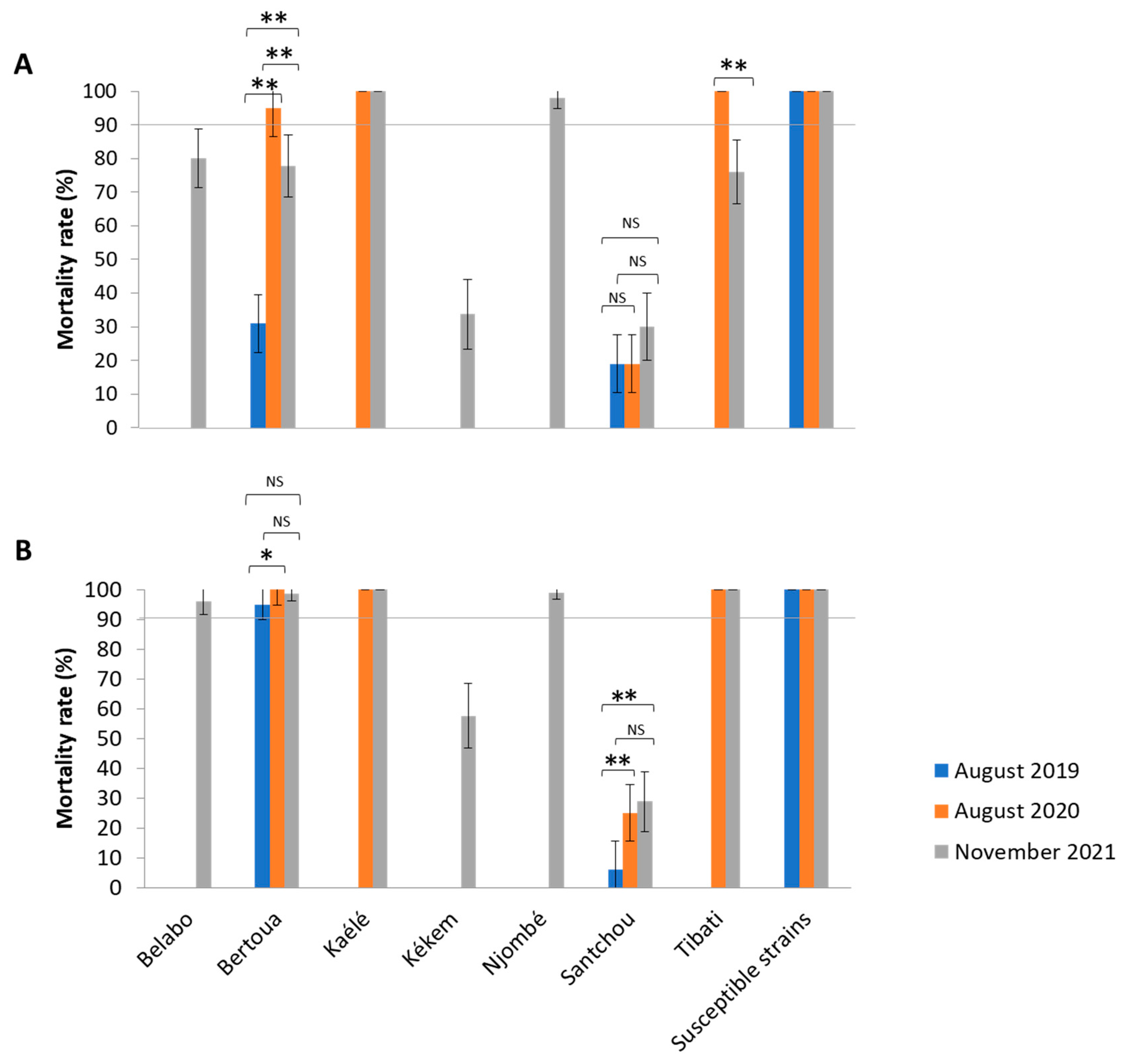
2.1. Bioassays Results
2.2. Molecular Identification of Species of the Anopheles gambiae Complex
2.3. Screening of Target Site Mutations (Ace-1 G119S)
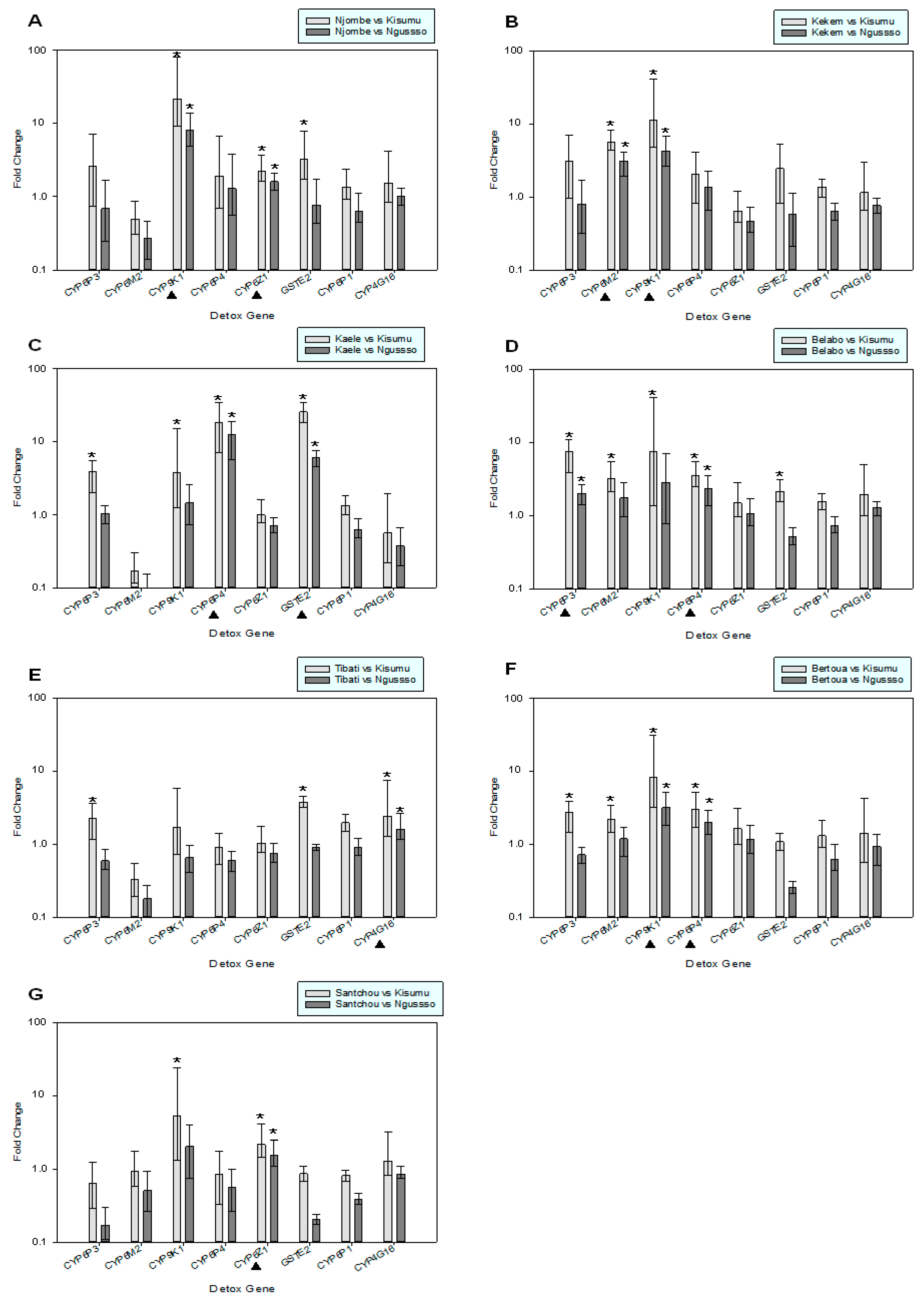
2.4. Expression Analysis of Detoxification Genes
3. Discussion
4. Methods
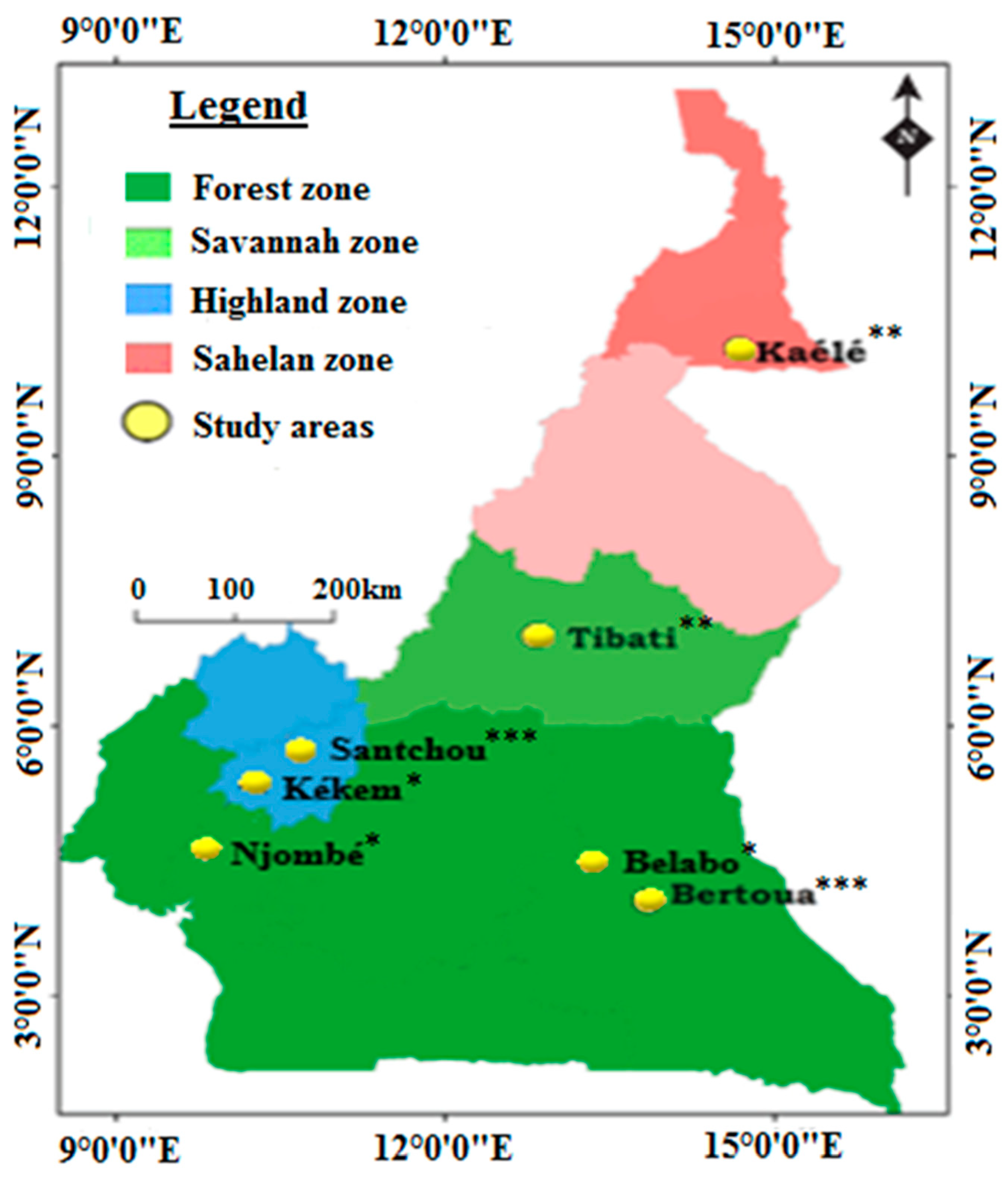
4.1. Study Site
4.2. Collection of Mosquito Larvae, Rearing, and Conservation
4.3. Insecticide Bioassay
4.4. Mosquito Processing
4.4.1. Total RNA and DNA Extraction from Mosquito Pools
4.4.2. Species Identification
4.5. Genotyping of G119S Mutation
4.6. Resistance Gene Expression Analyses
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van den Berg, H.; da Silva Bezerra, H.S.; Al-Eryani, S.; Chanda, E.; Nagpal, B.N.; Knox, T.B.; Velayudhan, R.; Yadav, R.S. Recent Trends in Global Insecticide Use for Disease Vector Control and Potential Implications for Resistance Management. Sci. Rep. 2021, 11, 23867. [Google Scholar] [CrossRef] [PubMed]
- Zaim, M.; Aitio, A.; Nakashima, N. Safety of Pyrethroid-Treated Mosquito Nets. Med. Vet. Entomol. 2000, 14, 1–5. [Google Scholar] [CrossRef]
- World Malaria Report 2020. Available online: https://www.who.int/publications-detail-redirect/9789240015791 (accessed on 1 June 2022).
- Antonio-Nkondjio, C.; Sonhafouo-Chiana, N.; Ngadjeu, C.S.; Doumbe-Belisse, P.; Talipouo, A.; Djamouko-Djonkam, L.; Kopya, E.; Bamou, R.; Awono-Ambene, P.; Wondji, C.S. Review of the Evolution of Insecticide Resistance in Main Malaria Vectors in Cameroon from 1990 to 2017. Parasit. Vectors 2017, 10, 472. [Google Scholar] [CrossRef] [PubMed]
- Djiappi-Tchamen, B.; Nana-Ndjangwo, M.S.; Mavridis, K.; Talipouo, A.; Nchoutpouen, E.; Makoudjou, I.; Bamou, R.; Mayi, A.M.P.; Awono-Ambene, P.; Tchuinkam, T.; et al. Analyses of Insecticide Resistance Genes in Aedes Aegypti and Aedes Albopictus Mosquito Populations from Cameroon. Genes 2021, 12, 828. [Google Scholar] [CrossRef] [PubMed]
- Ranson, H.; Lissenden, N. Insecticide Resistance in African Anopheles Mosquitoes: A Worsening Situation That Needs Urgent Action to Maintain Malaria Control. Trends Parasitol. 2016, 32, 187–196. [Google Scholar] [CrossRef]
- Talipouo, A.; Mavridis, K.; Nchoutpouen, E.; Djiappi-Tchamen, B.; Fotakis, E.A.; Kopya, E.; Bamou, R.; Kekeunou, S.; Awono-Ambene, P.; Balabanidou, V.; et al. High Insecticide Resistance Mediated by Different Mechanisms in Culex Quinquefasciatus Populations from the City of Yaoundé, Cameroon. Sci. Rep. 2021, 11, 7322. [Google Scholar] [CrossRef]
- Hemingway, J.; Vontas, J.; Poupardin, R.; Raman, J.; Lines, J.; Schwabe, C.; Matias, A.; Kleinschmidt, I. Country-Level Operational Implementation of the Global Plan for Insecticide Resistance Management. Proc. Natl. Acad. Sci. USA 2013, 110, 9397–9402. [Google Scholar] [CrossRef] [Green Version]
- Vontas, J.; Katsavou, E.; Mavridis, K. Cytochrome P450-Based Metabolic Insecticide Resistance in Anopheles and Aedes Mosquito Vectors: Muddying the Waters. Pestic. Biochem. Physiol. 2020, 170, 104666. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Weiss, D.J.; Cameron, E.; Bisanzio, D.; Mappin, B.; Dalrymple, U.; Battle, K.; Moyes, C.L.; Henry, A.; Eckhoff, P.A.; et al. The Effect of Malaria Control on Plasmodium Falciparum in Africa between 2000 and 2015. Nature 2015, 526, 207–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Churcher, T.S.; Lissenden, N.; Griffin, J.T.; Worrall, E.; Ranson, H. The Impact of Pyrethroid Resistance on the Efficacy and Effectiveness of Bednets for Malaria Control in Africa. eLife 2016, 5, e16090. [Google Scholar] [CrossRef]
- Agossa, F.R.; Aïkpon, R.; Azondékon, R.; Govoetchan, R.; Padonou, G.G.; Oussou, O.; Oké-Agbo, F.; Akogbéto, M.C. Efficacy of Various Insecticides Recommended for Indoor Residual Spraying: Pirimiphos Methyl, Potential Alternative to Bendiocarb for Pyrethroid Resistance Management in Benin, West Africa. Trans. R. Soc. Trop. Med. Hyg. 2014, 108, 84–91. [Google Scholar] [CrossRef]
- Blaschke, T.F.; Lumpkin, M.; Hartman, D. The World Health Organization Prequalification Program and Clinical Pharmacology in 2030. Clin. Pharmacol. Ther. 2020, 107, 68–71. [Google Scholar] [CrossRef]
- World Health Organization. World Health Statistics 2012; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Dengela, D.; Seyoum, A.; Lucas, B.; Johns, B.; George, K.; Belemvire, A.; Caranci, A.; Norris, L.C.; Fornadel, C.M. Multi-Country Assessment of Residual Bio-Efficacy of Insecticides Used for Indoor Residual Spraying in Malaria Control on Different Surface Types: Results from Program Monitoring in 17 PMI/USAID-Supported IRS Countries. Parasit. Vectors 2018, 11, 71. [Google Scholar] [CrossRef]
- Aïkpon, R.; Agossa, F.; Ossè, R.; Oussou, O.; Aïzoun, N.; Oké-Agbo, F.; Akogbéto, M. Bendiocarb Resistance in Anopheles Gambiae s.l. Populations from Atacora Department in Benin, West Africa: A Threat for Malaria Vector Control. Parasit. Vectors 2013, 6, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djogbénou, L.; Weill, M.; Hougard, J.-M.; Raymond, M.; Akogbéto, M.; Chandre, F. Characterization of Insensitive Acetylcholinesterase (Ace-1R) in Anopheles Gambiae (Diptera: Culicidae): Resistance Levels and Dominance. J. Med. Entomol. 2007, 44, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Edi, C.V.A.; Koudou, B.G.; Jones, C.M.; Weetman, D.; Ranson, H. Multiple-Insecticide Resistance in Anopheles Gambiae Mosquitoes, Southern Côte d’Ivoire. Emerg. Infect. Dis. 2012, 18, 1508–1511. [Google Scholar] [CrossRef] [PubMed]
- Antonio-Nkondjio, C.; Poupardin, R.; Tene, B.F.; Kopya, E.; Costantini, C.; Awono-Ambene, P.; Wondji, C.S. Investigation of Mechanisms of Bendiocarb Resistance in Anopheles Gambiae Populations from the City of Yaoundé, Cameroon. Malar. J. 2016, 15, 424. [Google Scholar] [CrossRef] [Green Version]
- Menze, B.D.; Wondji, M.J.; Tchapga, W.; Tchoupo, M.; Riveron, J.M.; Wondji, C.S. Bionomics and Insecticides Resistance Profiling of Malaria Vectors at a Selected Site for Experimental Hut Trials in Central Cameroon. Malar. J. 2018, 17, 317. [Google Scholar] [CrossRef]
- Aïkpon, R.; Sèzonlin, M.; Ossè, R.; Akogbéto, M. Evidence of Multiple Mechanisms Providing Carbamate and Organophosphate Resistance in Field An. Gambiae Population from Atacora in Benin. Parasit. Vectors 2014, 7, 568. [Google Scholar] [CrossRef]
- Essandoh, J.; Yawson, A.E.; Weetman, D. Acetylcholinesterase (Ace-1) Target Site Mutation 119S Is Strongly Diagnostic of Carbamate and Organophosphate Resistance in Anopheles Gambiae s.s. and Anopheles Coluzzii across Southern Ghana. Malar. J. 2013, 12, 404. [Google Scholar] [CrossRef] [Green Version]
- Bamou, R.; Sonhafouo-Chiana, N.; Mavridis, K.; Tchuinkam, T.; Wondji, C.S.; Vontas, J.; Antonio-Nkondjio, C. Status of Insecticide Resistance and Its Mechanisms in Anopheles Gambiae and Anopheles Coluzzii Populations from Forest Settings in South Cameroon. Genes 2019, 10, 741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binyang, A.J.; Elanga-Ndille, E.; Tene-Fossog, B.; Ndo, C.; Nouage, L.; Assatse, T.; Fotso-Toguem, Y.; Tabue, R.; Zeukeng, F.; Nguiffo, D.N.; et al. Distribution of Acetylcholinesterase (Ace-1R) Target-Site G119S Mutation and Resistance to Carbamates and Organophosphates in Anopheles Gambiae Sensu Lato Populations from Cameroon. Parasit. Vectors 2022, 15, 53. [Google Scholar] [CrossRef]
- Elanga-Ndille, E.; Nouage, L.; Ndo, C.; Binyang, A.; Assatse, T.; Nguiffo-Nguete, D.; Djonabaye, D.; Irwing, H.; Tene-Fossog, B.; Wondji, C.S. The G119S Acetylcholinesterase (Ace-1) Target Site Mutation Confers Carbamate Resistance in the Major Malaria Vector Anopheles Gambiae from Cameroon: A Challenge for the Coming IRS Implementation. Genes 2019, 10, 790. [Google Scholar] [CrossRef] [Green Version]
- Pouokam, G.B.; Lemnyuy Album, W.; Ndikontar, A.S.; Sidatt, M.E.H. A Pilot Study in Cameroon to Understand Safe Uses of Pesticides in Agriculture, Risk Factors for Farmers’ Exposure and Management of Accidental Cases. Toxics 2017, 5, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamdem, C.; Fossog, B.T.; Simard, F.; Etouna, J.; Ndo, C.; Kengne, P.; Boussès, P.; Etoa, F.-X.; Awono-Ambene, P.; Fontenille, D.; et al. Anthropogenic Habitat Disturbance and Ecological Divergence between Incipient Species of the Malaria Mosquito Anopheles Gambiae. PLoS ONE 2012, 7, e39453. [Google Scholar] [CrossRef] [Green Version]
- Tene Fossog, B.; Ayala, D.; Acevedo, P.; Kengne, P.; Ngomo Abeso Mebuy, I.; Makanga, B.; Magnus, J.; Awono-Ambene, P.; Njiokou, F.; Pombi, M.; et al. Habitat Segregation and Ecological Character Displacement in Cryptic African Malaria Mosquitoes. Evol. Appl. 2015, 8, 326–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibrahim, S.S.; Fadel, A.N.; Tchouakui, M.; Terence, E.; Wondji, M.J.; Tchoupo, M.; Kérah-Hinzoumbé, C.; Wanji, S.; Wondji, C.S. High Insecticide Resistance in the Major Malaria Vector Anopheles Coluzzii in Chad Republic. Infect. Dis. Poverty 2019, 8, 100. [Google Scholar] [CrossRef]
- Fadel, A.N.; Ibrahim, S.S.; Tchouakui, M.; Terence, E.; Wondji, M.J.; Tchoupo, M.; Wanji, S.; Wondji, C.S. A Combination of Metabolic Resistance and High Frequency of the 1014F Kdr Mutation Is Driving Pyrethroid Resistance in Anopheles Coluzzii Population from Guinea Savanna of Cameroon. Parasit. Vectors 2019, 12, 263. [Google Scholar] [CrossRef]
- Asidi, A.N.; N’ Guessan, R.; Koffi, A.A.; Curtis, C.F.; Hougard, J.-M.; Chandre, F.; Corbel, V.; Darriet, F.; Zaim, M.; Rowland, M.W. Experimental Hut Evaluation of Bednets Treated with an Organophosphate (Chlorpyrifos-Methyl) or a Pyrethroid (Lambdacyhalothrin) Alone and in Combination against Insecticide-Resistant Anopheles Gambiae and Culex Quinquefasciatus Mosquitoes. Malar. J. 2005, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Antonio-Nkondjio, C.; Fossog, B.T.; Ndo, C.; Djantio, B.M.; Togouet, S.Z.; Awono-Ambene, P.; Costantini, C.; Wondji, C.S.; Ranson, H. Anopheles Gambiae Distribution and Insecticide Resistance in the Cities of Douala and Yaoundé (Cameroon): Influence of Urban Agriculture and Pollution. Malar. J. 2011, 10, 154. [Google Scholar] [CrossRef] [Green Version]
- Yunta, C.; Hemmings, K.; Stevenson, B.; Koekemoer, L.L.; Matambo, T.; Pignatelli, P.; Voice, M.; Nász, S.; Paine, M.J.I. Cross-Resistance Profiles of Malaria Mosquito P450s Associated with Pyrethroid Resistance against WHO Insecticides. Pestic. Biochem. Physiol. 2019, 161, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Adolfi, A.; Poulton, B.; Anthousi, A.; Macilwee, S.; Ranson, H.; Lycett, G.J. Functional Genetic Validation of Key Genes Conferring Insecticide Resistance in the Major African Malaria Vector, Anopheles Gambiae. Proc. Natl. Acad. Sci. USA 2019, 116, 25764–25772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nwane, P.; Etang, J.; Chouaibou, M.; Toto, J.C.; Kerah-Hinzoumbé, C.; Mimpfoundi, R.; Awono-Ambene, H.P.; Simard, F. Trends in DDT and Pyrethroid Resistance in Anopheles Gambiaes.s. Populations from Urban and Agro-Industrial Settings in Southern Cameroon. BMC Infect. Dis. 2009, 9, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balabanidou, V.; Kampouraki, A.; MacLean, M.; Blomquist, G.J.; Tittiger, C.; Juárez, M.P.; Mijailovsky, S.J.; Chalepakis, G.; Anthousi, A.; Lynd, A.; et al. Cytochrome P450 Associated with Insecticide Resistance Catalyzes Cuticular Hydrocarbon Production in Anopheles Gambiae. Proc. Natl. Acad. Sci. USA 2016, 113, 9268–9273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kala Chouakeu, N.A.; Ngingahi, L.G.; Bamou, R.; Talipouo, A.; Ngadjeu, C.S.; Mayi, M.P.A.; Kopya, E.; Awono-Ambene, P.; Tchuinkam, T.; Antonio Nkondjio, C. Knowledge, Attitude, and Practices (KAP) of Human Populations towards Malaria Control in Four Ecoepidemiological Settings in Cameroon. J. Trop. Med. 2021, 2021, e9925135. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Test Procedures for Insecticide Resistance Monitoring in Malaria Vector Mosquitoes, 2nd ed.; World Health Organization: Geneva, Switzerland, 2018; ISSN 978-92-4-151157-5. [Google Scholar]
- Bass, C.; Nikou, D.; Vontas, J.; Williamson, M.S.; Field, L.M. Development of High-Throughput Real-Time PCR Assays for the Identification of Insensitive Acetylcholinesterase (Ace-1R) in Anopheles Gambiae. Pestic. Biochem. Physiol. 2010, 96, 80–85. [Google Scholar] [CrossRef]
- Wipf, N.C.; Duchemin, W.; Kouadio, F.-P.A.; Fodjo, B.K.; Sadia, C.G.; Mouhamadou, C.S.; Vavassori, L.; Mäser, P.; Mavridis, K.; Vontas, J.; et al. Multi-Insecticide Resistant Malaria Vectors in the Field Remain Susceptible to Malathion, despite the Presence of Ace1 Point Mutations. PLoS Genet. 2022, 18, e1009963. [Google Scholar] [CrossRef]
- Mavridis, K.; Wipf, N.; Müller, P.; Traoré, M.M.; Muller, G.; Vontas, J. Detection and Monitoring of Insecticide Resistance Mutations in Anopheles Gambiae: Individual vs. Pooled Specimens. Genes 2018, 9, 479. [Google Scholar] [CrossRef] [Green Version]
- Mavridis, K.; Wipf, N.; Medves, S.; Erquiaga, I.; Müller, P.; Vontas, J. Rapid Multiplex Gene Expression Assays for Monitoring Metabolic Resistance in the Major Malaria Vector Anopheles Gambiae. Parasit. Vectors 2019, 12, 9. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Population/Sites | Sites Characteristics | Sample Size | Species ID | % Frequency |
|---|---|---|---|---|
| Kisumu | Susceptible strain | 40 | An. gambiae | 100 |
| Ngousso | 40 | An. coluzzii | 100 | |
| Tibati | Sahelo-Sudanese (humid savannah) | 80 | An. coluzzii | 100 |
| Kaélé | Sahelian zone | An. arabiensis | 10.0 | |
| 60 | An. gambiae | 4.0 | ||
| An. coluzzii | 86.0 | |||
| Kékem | Highland grassfields | 80 | An. gambiae | 100 |
| Santchou | 120 | An. gambiae | 100 | |
| Bertoua | Forest zone | 79 | An. gambiae | 87.0 |
| An. coluzzii | 13.0 | |||
| Njombé | 54 | An. gambiae | 40.9 | |
| An. coluzzii | 59.1 | |||
| Belabo | 84 | An. gambiae | 82.0 | |
| An. coluzzii | 18.0 |
| PopulatioN | Category | Sample Size | % G119S (ace-1) |
|---|---|---|---|
| Kisumu | Susceptible lab strain | 40 | 0.0 ± 0.0 |
| Ngousso | Susceptible lab strain | 40 | 0.0 ± 0.0 |
| Njombé | Bendiocarb survivors | 3 | 75.5 |
| Malathion survivors | 1 | 100.0 | |
| Unexposed | 20 | 4.5 ± 2.5 | |
| Kékem | Bendiocarb survivors | 20 | 84.6 ± 5.1 |
| Unexposed | 20 | 41.1 ± 18.3 | |
| Bélabo | Bendiocarb survivors | 20 | 54.1 ± 9.6 |
| Malathion survivors | 4 | 72.3 | |
| Unexposed | 20 | 10.25 ± 5.9 | |
| Tibati | Bendiocarb survivors | 20 | 81.3 ± 18.7 |
| Unexposed | 20 | 0.0 ± 0.0 | |
| Bertoua | Bendiocarb survivors | 18 | 71.0 ± 9.9 |
| Malathion survivors | 1 | 77.1 | |
| Unexposed | 20 | 36.1 ± 9.4 | |
| Santchou | Bendiocarb survivors | 20 | 87.3 ± 1.75 |
| Malathion survivors | 40 | 86.93 ± 8.0 | |
| Unexposed | 20 | 87.9 ± 12.2 |
| Study sites | Kékem | Njombé | Belabo | Kaélé a | Tibati a | Santchou a | Bertoua a |
|---|---|---|---|---|---|---|---|
| Administrative region | West | Littoral | East | Far North | Adamawa | West | East |
| Coordinates | 5°10′ N, 10°02′ E | 4°64′ N, 9°67′ E | 4°56′ N, 13°18′ E | 10°50′ N, 14°56′ E | 12°37′ N, 12°37′ E | 5°58′ N, 9°58′ E | 4°34′ N, 13°41′ E |
| Domain | Highland Grassfields | Forest zone | Forest zone | Sahelian zone | Sahelo soudanese | Highland Grassfields | Forest zone |
| Climate | Equatorial | Equatorial | Subtropical | Sahelian | Tropical humid | Equatorial | Subtropical |
| Seasons | Dry season (November to March), rainy season (April to October) | Dry season (December to February), rainy season (March to November) | Dry season (December to March and July), rainy season (March to June and August to November) | Dry season (October to May), rainy season (June to September) | Dry season (November to April), rainy season (May to October) | Dry season (November to March), rainy season (April to October), | Dry season (December to March and July), rainy season (March to June and August to November) |
| Vegetation | Grassland | Grassland | Evergreen degraded forest | Wooded Savanah | Grassy Savanah | Grassland | Evergreen degraded forest |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngangue-Siewe, I.N.; Ndjeunia-Mbiakop, P.; Kala-Chouakeu, N.A.; Bamou, R.; Talipouo, A.; Djamouko-Djonkam, L.; Vontas, J.; Mavridis, K.; Tombi, J.; Tchuinkam, T.; et al. Bendiocarb and Malathion Resistance in Two Major Malaria Vector Populations in Cameroon Is Associated with High Frequency of the G119S Mutation (Ace-1) and Overexpression of Detoxification Genes. Pathogens 2022, 11, 824. https://doi.org/10.3390/pathogens11080824
Ngangue-Siewe IN, Ndjeunia-Mbiakop P, Kala-Chouakeu NA, Bamou R, Talipouo A, Djamouko-Djonkam L, Vontas J, Mavridis K, Tombi J, Tchuinkam T, et al. Bendiocarb and Malathion Resistance in Two Major Malaria Vector Populations in Cameroon Is Associated with High Frequency of the G119S Mutation (Ace-1) and Overexpression of Detoxification Genes. Pathogens. 2022; 11(8):824. https://doi.org/10.3390/pathogens11080824
Chicago/Turabian StyleNgangue-Siewe, Idriss Nasser, Paulette Ndjeunia-Mbiakop, Nelly Armanda Kala-Chouakeu, Roland Bamou, Abdou Talipouo, Landre Djamouko-Djonkam, John Vontas, Konstantinos Mavridis, Jeannette Tombi, Timoléon Tchuinkam, and et al. 2022. "Bendiocarb and Malathion Resistance in Two Major Malaria Vector Populations in Cameroon Is Associated with High Frequency of the G119S Mutation (Ace-1) and Overexpression of Detoxification Genes" Pathogens 11, no. 8: 824. https://doi.org/10.3390/pathogens11080824