
Survival of Campylobacter jejuni Co-Cultured with Salmonella spp. in Aerobic Conditions

Abstract
:1. Introduction
2. Results
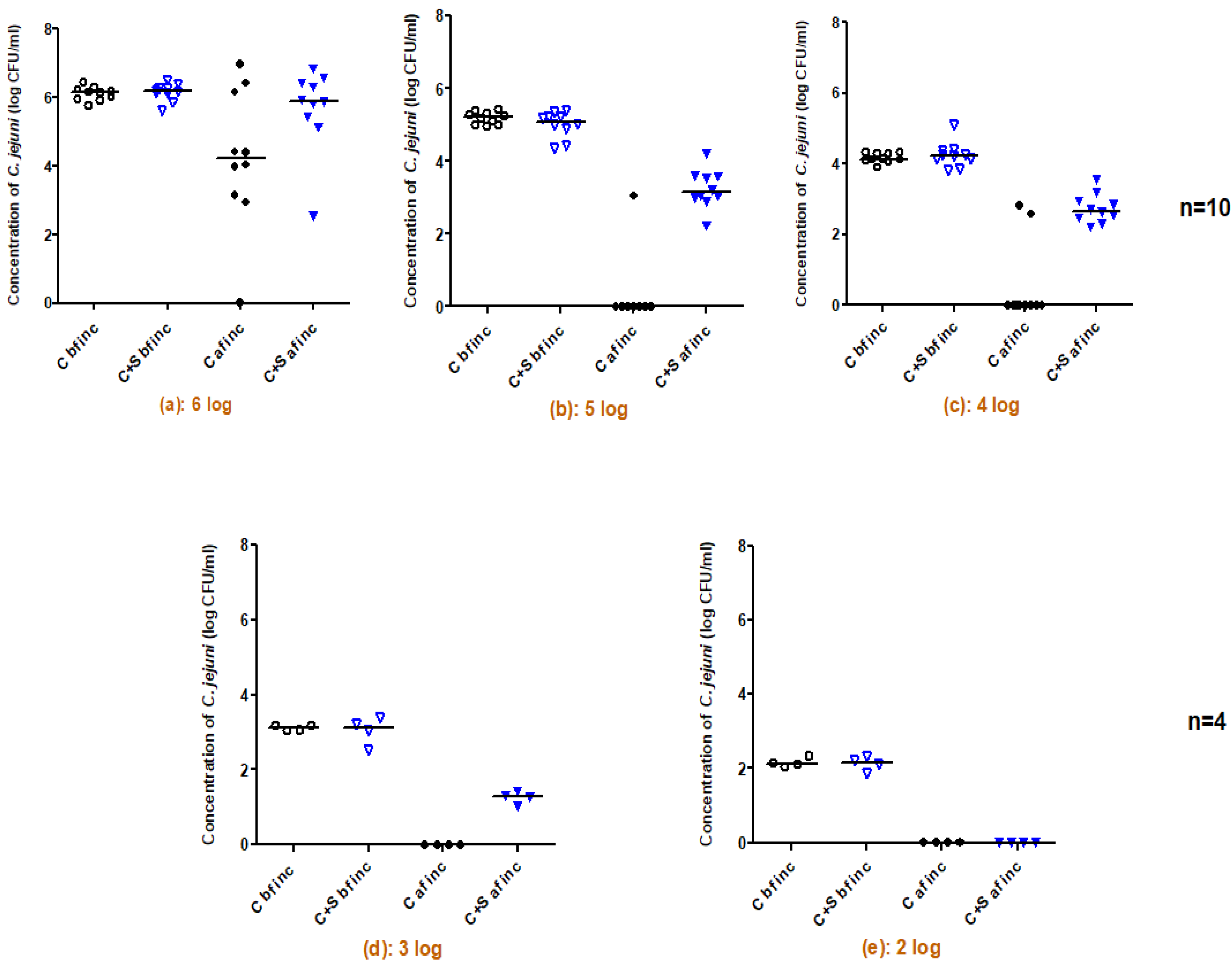
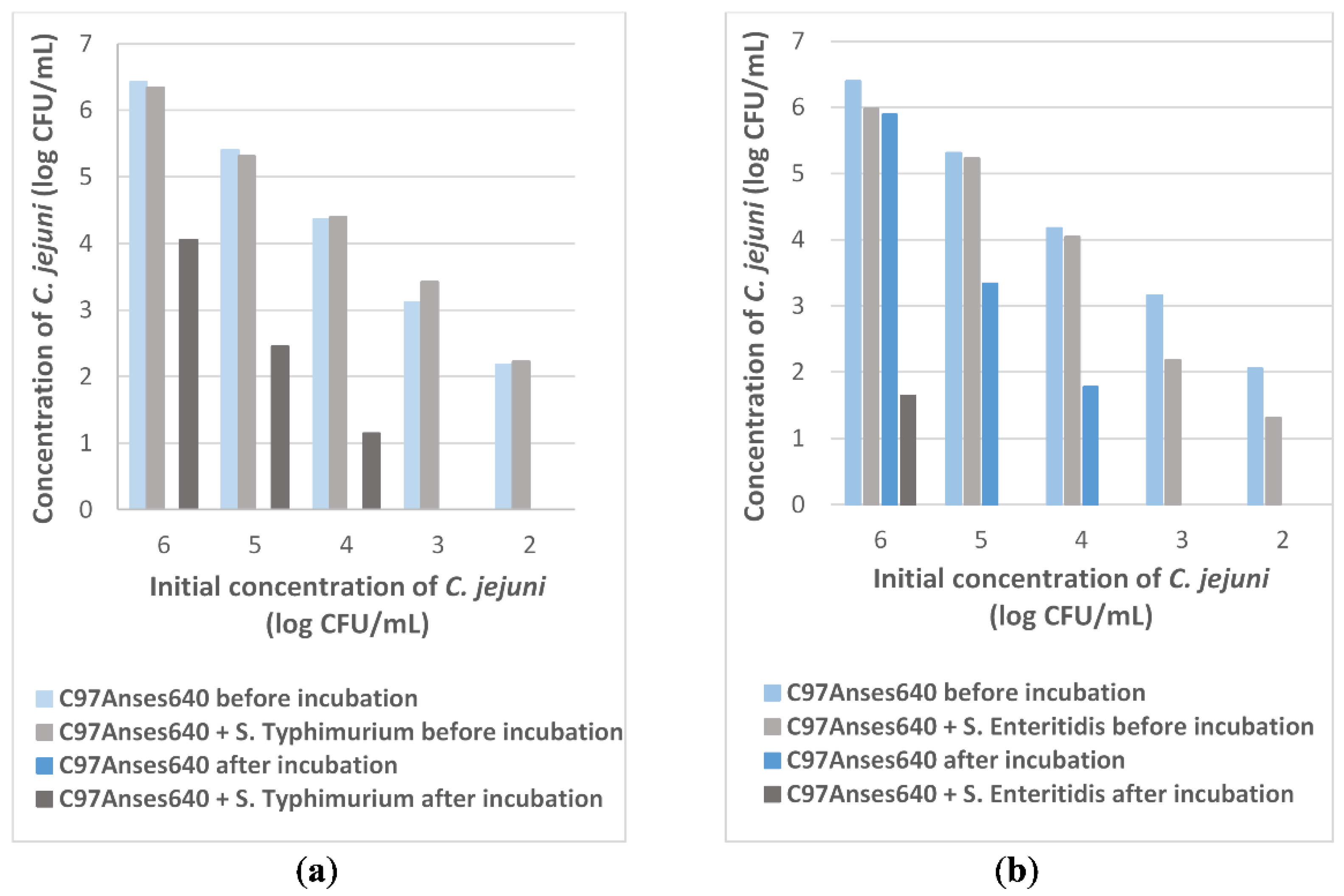
2.1. Survival of C. jejuni Co-Cultured with Salmonella spp. as a Function of Different C. jejuni Inoculum Levels
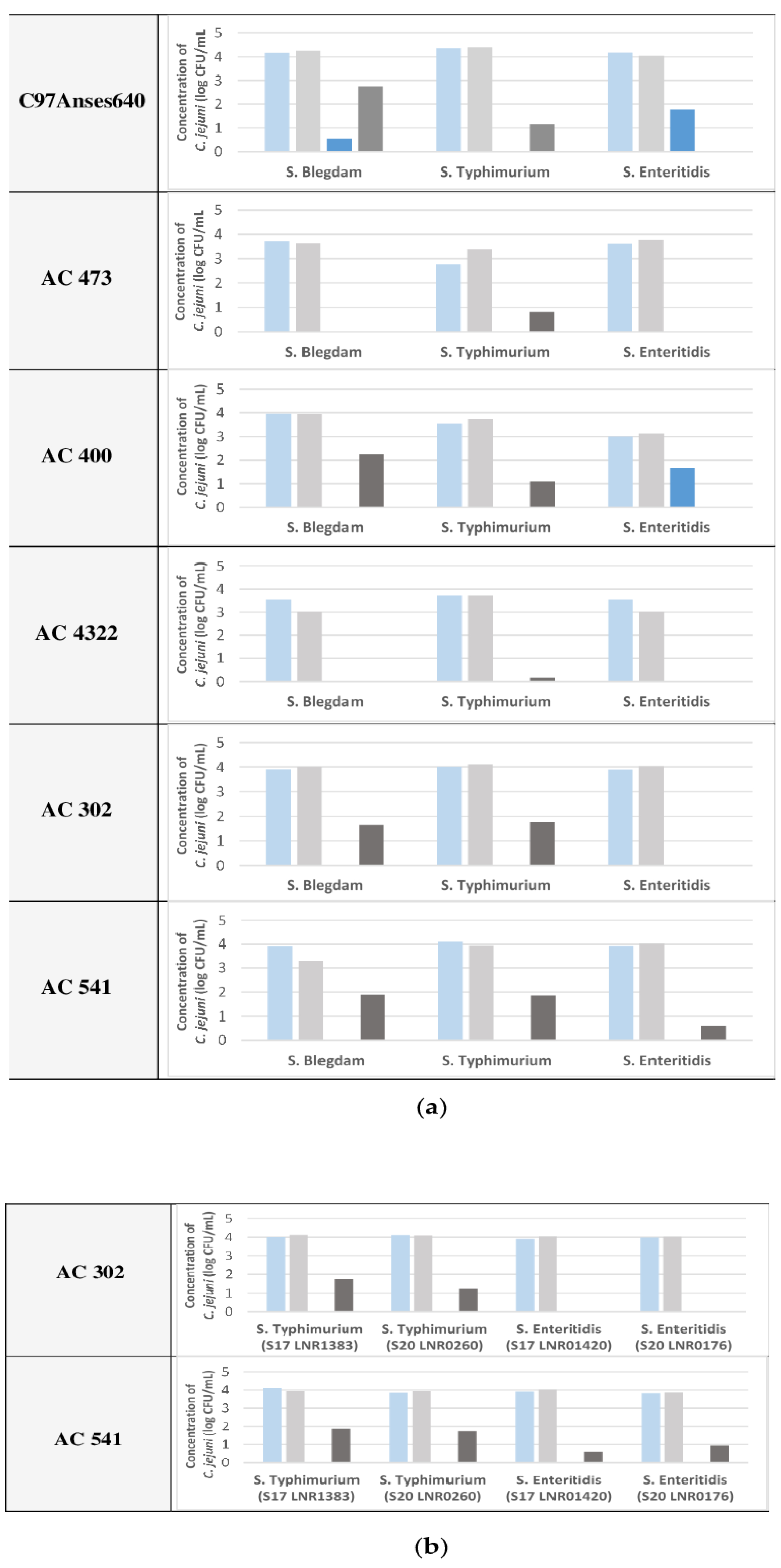
2.2. Survival of C. jejuni Co-Cultured with Salmonella spp. as a Function of Different Strains
2.3. Effect of C. jejuni on Salmonella Growth under Aerobic Conditions
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
- -
- different final concentrations of C. jejuni ranging from 2 to 6 log CFU/mL to study the survival of C. jejuni co-cultured with Salmonella spp. as a function of different C. jejuni inoculum levels.
- -
- a final concentration of 4 log CFU/mL to study the survival of C. jejuni co-cultured with Salmonella spp. as a function of different strains.
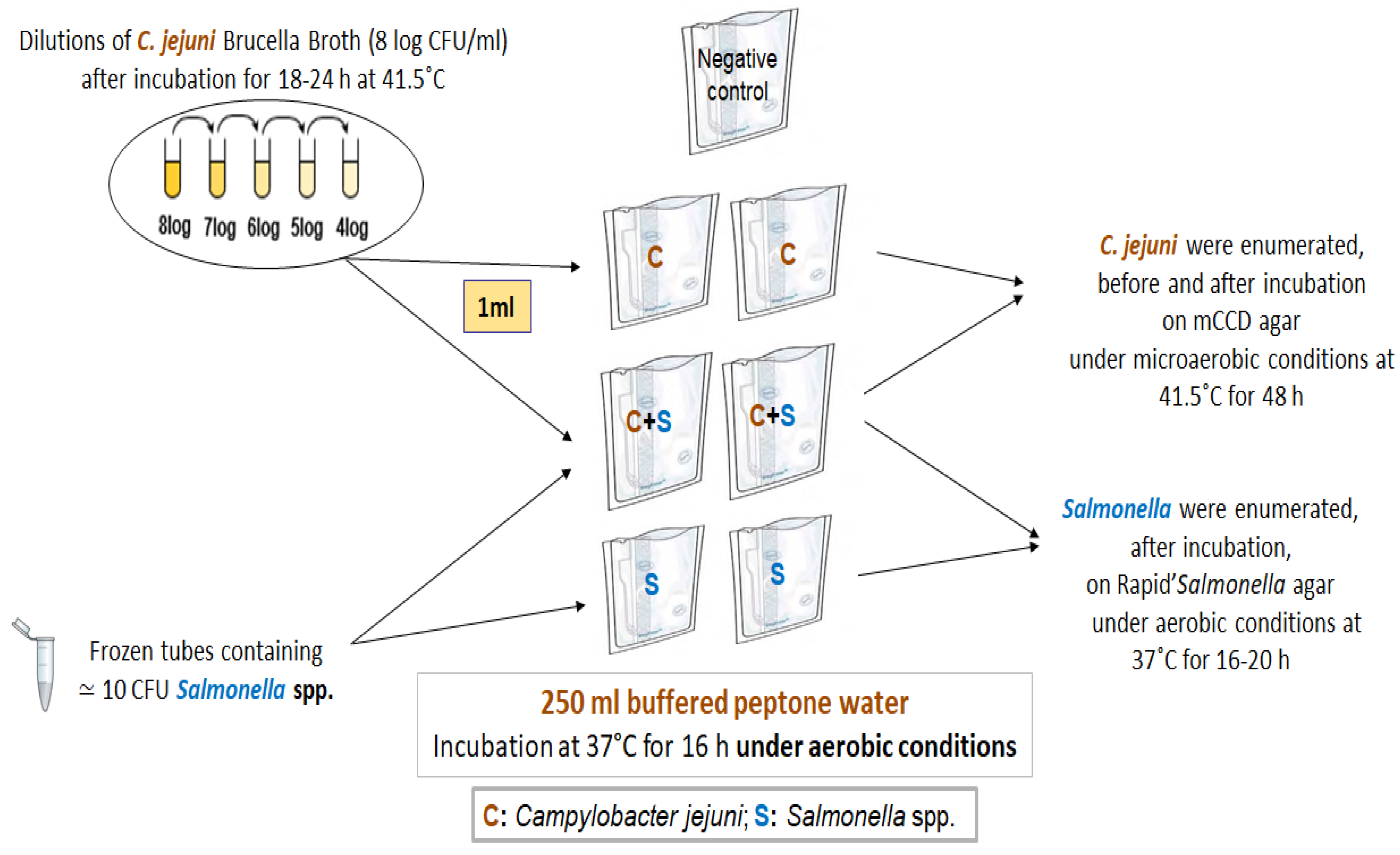
4.2. Survival Assays
4.3. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- EFSA; ECDC. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar]
- EFSA; ECDC. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, e06406. [Google Scholar]
- Sarkar, S.R.; Hossain, M.A.; Paul, S.K.; Ray, N.C.; Sultana, S.; Rahman, M.M.; Islam, A. Campylobacteriosis—An overview. Mymensingh. Med. J. 2014, 23, 173–180. [Google Scholar]
- EFSA. ECDC. The European Union One Health 2018 Zoonoses Report. EFSA J. 2019, 17, e05926. [Google Scholar]
- Igwaran, A.; Okoh, A.I. Human campylobacteriosis: A public health concern of global importance. Heliyon 2019, 5, e02814. [Google Scholar] [CrossRef] [PubMed]
- Butzler, J.P. Campylobacter, from obscurity to celebrity. Clin. Microbiol. Infect. 2004, 10, 868–876. [Google Scholar] [CrossRef] [Green Version]
- Hue, O.; Le Bouquin, S.; Lalande, F.; Allain, V.; Rouxel, S.; Petetin, I.; Quesne, S.; Laisney, M.-J.; Gloaguen, P.-Y.; Picherot, M.; et al. Prevalence of Salmonella spp. on broiler chicken carcasses and risk factors at the slaughterhouse in France in 2008. Food Control 2011, 22, 1158–1164. [Google Scholar] [CrossRef]
- Zilbauer, M.; Dorrell, N.; Wren, B.W.; Bajaj-Elliott, M. Campylobacter jejuni-mediated disease pathogenesis: An update. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 123–129. [Google Scholar] [CrossRef]
- Van Cauteren, D.; De Valk, H.; Sommen, C.; King, L.A.; Jourdan-Da Silva, N.; Weill, F.X.; Le Hello, S.; Megraud, F.; Vaillant, V.; Desenclos, J.C. Community Incidence of Campylobacteriosis and Nontyphoidal Salmonellosis, France, 2008–2013. Foodborne Pathog. Dis. 2015, 12, 664–669. [Google Scholar] [CrossRef] [Green Version]
- Skarp, C.P.A.; Hanninen, M.L.; Rautelin, H.I.K. Campylobacteriosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 103–109. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Scientific Opinion on Campylobacter in broiler meat production: Control options and performance objectives and/or targets at different stages of the food chain. EFSA J. 2011, 9, 2105. [Google Scholar] [CrossRef]
- Antunes, P.; Mourao, J.; Campos, J.; Peixe, L. Salmonellosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 110–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA. ECDC. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSA J. 2017, 15, e05077. [Google Scholar]
- Awad, W.A.; Ghareeb, K. Some aspects of control of Salmonella infection in poultry for minimising contamination in the food chain. World’s Poult. Sci. J. 2019, 70, 519–530. [Google Scholar] [CrossRef]
- Guyard-Nicodeme, M.; Tresse, O.; Houard, E.; Jugiau, F.; Courtillon, C.; El Manaa, K.; Laisney, M.J.; Chemaly, M. Characterization of Campylobacter spp. transferred from naturally contaminated chicken legs to cooked chicken slices via a cutting board. Int. J. Food Microbiol. 2013, 164, 7–14. [Google Scholar] [CrossRef]
- Saravanan, S.; Purushothaman, V.; Murthy, T.R.; Sukumar, K.; Srinivasan, P.; Gowthaman, V.; Balusamy, M.; Atterbury, R.; Kuchipudi, S.V. Molecular Epidemiology of Nontyphoidal Salmonella in Poultry and Poultry Products in India: Implications for Human Health. Indian J. Microbiol. 2015, 55, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Hugas, M.; Beloeil, P. Controlling Salmonella along the food chain in the European Union—Progress over the last ten years. Euro Surveill. 2014, 19, 20804. [Google Scholar] [CrossRef] [Green Version]
- Hue, O.; Le Bouquin, S.; Laisney, M.J.; Allain, V.; Lalande, F.; Petetin, I.; Rouxel, S.; Quesne, S.; Gloaguen, P.Y.; Picherot, M.; et al. Prevalence of and risk factors for Campylobacter spp. contamination of broiler chicken carcasses at the slaughterhouse. Food Microbiol. 2010, 27, 992–999. [Google Scholar] [CrossRef]
- Rouger, A.; Tresse, O.; Zagorec, M. Bacterial Contaminants of Poultry Meat: Sources, Species, and Dynamics. Microorganisms 2017, 5, 50. [Google Scholar] [CrossRef]
- Hue, O.; Allain, V.; Laisney, M.J.; Le Bouquin, S.; Lalande, F.; Petetin, I.; Rouxel, S.; Quesne, S.; Gloaguen, P.Y.; Picherot, M.; et al. Campylobacter contamination of broiler caeca and carcasses at the slaughterhouse and correlation with Salmonella contamination. Food Microbiol. 2011, 28, 862–868. [Google Scholar] [CrossRef]
- EFSA. Simulation-based assessment of Microbiological Criteria on Salmonella in poultry meat. EFSA J. 2011, 9, 1986. [Google Scholar]
- EFSA BIOHAZ Panel; Koutsoumanis, K.; Allende, A.; Alvarez-Ordonez, A.; Bolton, D.; Bover-Cid, S.; Davies, R.; De Cesare, A.; Herman, L.; Hilbert, F.; et al. Update and review of control options for Campylobacter in broilers at primary production. EFSA J. 2020, 18, e06090. [Google Scholar]
- EFSA BIOHAZ Panel; Koutsoumanis, K.; Allende, A.; Alvarez-Ordonez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; De Cesare, A.; Herman, L.; Hilbert, F.; et al. Scientific Opinion on the Salmonella control in poultry flocks and its public health impact. EFSA J. 2019, 17, e05596. [Google Scholar]
- Bronnec, V.; Turonova, H.; Bouju, A.; Cruveiller, S.; Rodrigues, R.; Demnerova, K.; Tresse, O.; Haddad, N.; Zagorec, M. Adhesion, Biofilm Formation, and Genomic Features of Campylobacter jejuni Bf, an Atypical Strain Able to Grow under Aerobic Conditions. Front. Microbiol. 2016, 7, 1002. [Google Scholar] [CrossRef] [Green Version]
- Hilbert, F.; Scherwitzel, M.; Paulsen, P.; Szostak, M.P. Survival of Campylobacter jejuni under conditions of atmospheric oxygen tension with the support of Pseudomonas spp. Appl. Environ. Microbiol. 2010, 76, 5911–5917. [Google Scholar] [CrossRef] [Green Version]
- Bui, X.T.; Winding, A.; Qvortrup, K.; Wolff, A.; Bang, D.D.; Creuzenet, C. Survival of Campylobacter jejuni in co-culture with Acanthamoeba castellanii: Role of amoeba-mediated depletion of dissolved oxygen. Environ. Microbiol. 2012, 14, 2034–2047. [Google Scholar] [CrossRef]
- Axelsson-Olsson, D.; Waldenstrom, J.; Broman, T.; Olsen, B.; Holmberg, M. Protozoan Acanthamoeba polyphaga as a potential reservoir for Campylobacter jejuni. Appl. Environ. Microbiol. 2005, 71, 987–992. [Google Scholar] [CrossRef] [Green Version]
- Axelsson-Olsson, D.; Ellstrom, P.; Waldenstrom, J.; Haemig, P.D.; Brudin, L.; Olsen, B. Acanthamoeba-Campylobacter coculture as a novel method for enrichment of Campylobacter species. Appl. Environ. Microbiol. 2007, 73, 6864–6869. [Google Scholar] [CrossRef] [Green Version]
- Axelsson-Olsson, D.; Olofsson, J.; Svensson, L.; Griekspoor, P.; Waldenstrom, J.; Ellstrom, P.; Olsen, B. Amoebae and algae can prolong the survival of Campylobacter species in co-culture. Exp. Parasitol. 2010, 126, 59–64. [Google Scholar] [CrossRef]
- Karki, A.B.; Ballard, K.; Harper, C.; Sheaff, R.J.; Fakhr, M.K. Staphylococcus aureus enhances biofilm formation, aerotolerance, and survival of Campylobacter strains isolated from retail meats. Sci. Rep. 2021, 11, 13837. [Google Scholar] [CrossRef]
- Greige, S.; Rivoal, K.; Osman, M.; Safadi, D.E.; Dabboussi, F.; Hage, R.E.; Viscogliosi, E.; Hamze, M.; Chemaly, M. Prevalence and genetic diversity of Campylobacter spp. in the production chain of broiler chickens in Lebanon and its association with the intestinal protozoan Blastocystis sp. Poult. Sci. 2019, 98, 5883–5891. [Google Scholar] [CrossRef] [PubMed]
- Røder, H.L.; Raghupathi, P.K.; Herschend, J.; Brejnrod, A.; Knøchel, S.; Sørensen, S.J.; Burmølle, M. Interspecies interactions result in enhanced biofilm formation by co-cultures of bacteria isolated from a food processing environment. Food Microbiol. 2015, 51, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Hanning, I.; Jarquin, R.; Slavik, M. Campylobacter jejuni as a secondary colonizer of poultry biofilms. J. Appl. Microbiol. 2008, 105, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Giaouris, E.; Heir, E.; Desvaux, M.; Hebraud, M.; Moretro, T.; Langsrud, S.; Doulgeraki, A.; Nychas, G.J.; Kacaniova, M.; Czaczyk, K.; et al. Intra- and inter-species interactions within biofilms of important foodborne bacterial pathogens. Front. Microbiol. 2015, 6, 841. [Google Scholar] [CrossRef] [PubMed]
- Kaakoush, N.O.; Castano-Rodriguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, E.; McMullen, L.; Jeon, B. Impact of oxidative stress defense on bacterial survival and morphological change in Campylobacter jejuni under aerobic conditions. Front. Microbiol. 2015, 6, 295. [Google Scholar] [CrossRef]
- Oh, E.; McMullen, L.; Jeon, B. High Prevalence of Hyper-Aerotolerant Campylobacter jejuni in Retail Poultry with Potential Implication in Human Infection. Front. Microbiol. 2015, 6, 1263. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Pocheron, A.L.; Hernould, M.; Haddad, N.; Tresse, O.; Cappelier, J.M. Description of Campylobacter jejuni Bf, an atypical aero-tolerant strain. Gut Pathog. 2015, 7, 30. [Google Scholar] [CrossRef]
- Shagieva, E.; Demnerova, K.; Michova, H. Waterborne Isolates of Campylobacter jejuni Are Able to Develop Aerotolerance, Survive Exposure to Low Temperature, and Interact With Acanthamoeba polyphaga. Front. Microbiol. 2021, 12, 730858. [Google Scholar] [CrossRef]
- Vieira, A.; Ramesh, A.; Seddon, A.M.; Karlyshev, A.V. CmeABC Multidrug Efflux Pump Contributes to Antibiotic Resistance and Promotes Campylobacter jejuni Survival and Multiplication in Acanthamoeba polyphaga. Appl. Environ. Microbiol. 2017, 83, e01600-17. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, J.; Axelsson-Olsson, D.; Brudin, L.; Olsen, B.; Ellstrom, P. Campylobacter jejuni actively invades the amoeba Acanthamoeba polyphaga and survives within non digestive vacuoles. PLoS ONE 2013, 8, e78873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snelling, W.J.; McKenna, J.P.; Lecky, D.M.; Dooley, J.S. Survival of Campylobacter jejuni in waterborne protozoa. Appl. Environ. Microbiol. 2005, 71, 5560–5571. [Google Scholar] [CrossRef] [Green Version]
- Bare, J.; Sabbe, K.; Huws, S.; Vercauteren, D.; Braeckmans, K.; van Gremberghe, I.; Favoreel, H.; Houf, K. Influence of temperature, oxygen and bacterial strain identity on the association of Campylobacter jejuni with Acanthamoeba castellanii. FEMS Microbiol. Ecol. 2010, 74, 371–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, R.O.; Galán, J.E. Campylobacter jejuni survives within epithelial cells by avoiding delivery to lysosomes. PLoS Pathog. 2008, 4, e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dirks, B.P.; Quinlan, J.J. Development of a modified gentamicin protection assay to investigate the interaction between Campylobacter jejuni and Acanthamoeba castellanii ATCC 30010. Exp. Parasitol. 2014, 140, 39–43. [Google Scholar] [CrossRef]
- Kaakoush, N.O.; Miller, W.G.; De Reuse, H.; Mendz, G.L. Oxygen requirement and tolerance of Campylobacter jejuni. Res. Microbiol. 2007, 158, 644–650. [Google Scholar] [CrossRef]
- Thepault, A.; Guyard-Nicodeme, M.; Rose, V.; Quesne, S.; Queguiner, M.; Houard, E.; Megraud, F.; Rivoal, K.; Chemaly, M. A representative overview of the genetic diversity and lipooligosaccharide sialylation in Campylobacter jejuni along the broiler production chain in France and its comparison with human isolates. Int. J. Food Microbiol. 2018, 274, 20–30. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salmonella Counts (log CFU/mL) | ||||||||
|---|---|---|---|---|---|---|---|---|
| No C. jejuni | C. jejuni Strains | |||||||
| Salmonella Serovars and Strains | AC302 | AC 400 | AC 473 | AC 541 | AC 4322 | C97Anses640 | ||
| S. Blegdam | 8.6 | 8.5 | 8.5 | 8.5 | 8.5 | 8.5 | 8.8 | |
| S. Typhimurium | S17LNR1383 | 8.5 | 8.5 | 8.5 | 8.5 | 8.6 | 8.5 | 8.5 |
| S20LNR0260 | 8.5 | 8.5 | - | - | 8.5 | - | - | |
| S. Enteritidis | S17LNR01420 | 8.5 | 8.5 | 8.5 | 8.5 | 8.5 | 8.5 | 8.6 |
| S20LNR0176 | 8.4 | 8.5 | - | - | 8.4 | - | - | |
| S. Infantis | 8.5 | 8.5 | 8.5 | 8.6 | 8.6 | 8.5 | 8.8 | |
| S. Hadar | 8.7 | 8.6 | 8.6 | 8.7 | 8.7 | 8.7 | 8.8 | |
| S. Virchow | 8.6 | 8.2 | 8.0 | 8.7 | 8.5 | 8.7 | 8.6 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anis, N.; Bonifait, L.; Quesne, S.; Baugé, L.; Yassine, W.; Guyard-Nicodème, M.; Chemaly, M. Survival of Campylobacter jejuni Co-Cultured with Salmonella spp. in Aerobic Conditions. Pathogens 2022, 11, 812. https://doi.org/10.3390/pathogens11070812
Anis N, Bonifait L, Quesne S, Baugé L, Yassine W, Guyard-Nicodème M, Chemaly M. Survival of Campylobacter jejuni Co-Cultured with Salmonella spp. in Aerobic Conditions. Pathogens. 2022; 11(7):812. https://doi.org/10.3390/pathogens11070812
Chicago/Turabian StyleAnis, Nagham, Laetitia Bonifait, Ségolène Quesne, Louise Baugé, Wissam Yassine, Muriel Guyard-Nicodème, and Marianne Chemaly. 2022. "Survival of Campylobacter jejuni Co-Cultured with Salmonella spp. in Aerobic Conditions" Pathogens 11, no. 7: 812. https://doi.org/10.3390/pathogens11070812