
Genetic Diversity and Haplotype Analysis of Cattle Hydatid Cyst Isolates Using Mitochondrial Markers in Turkey

 ,
, 
Abstract
:1. Introduction
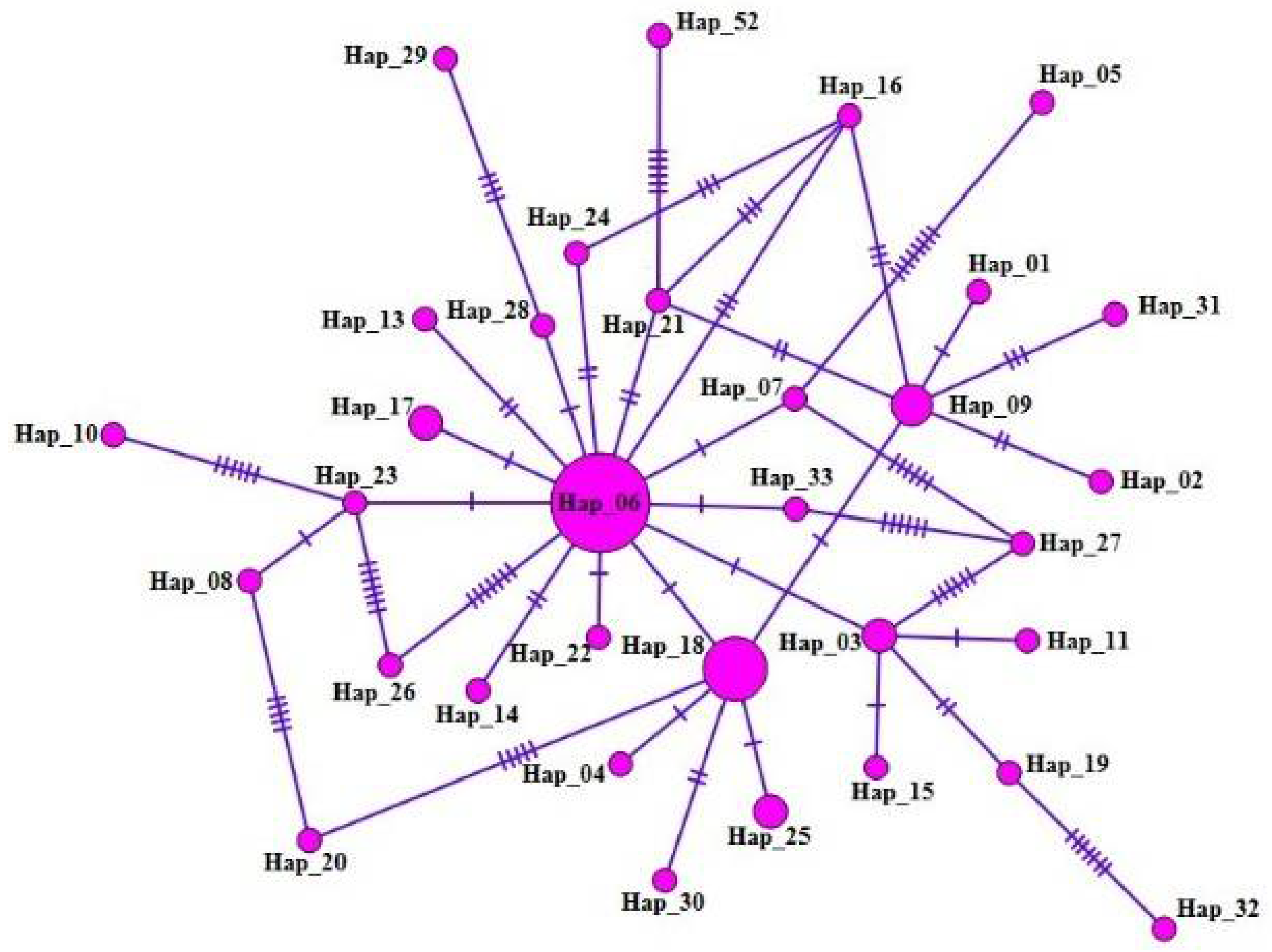
2. Results
3. Discussion
4. Material and Methods
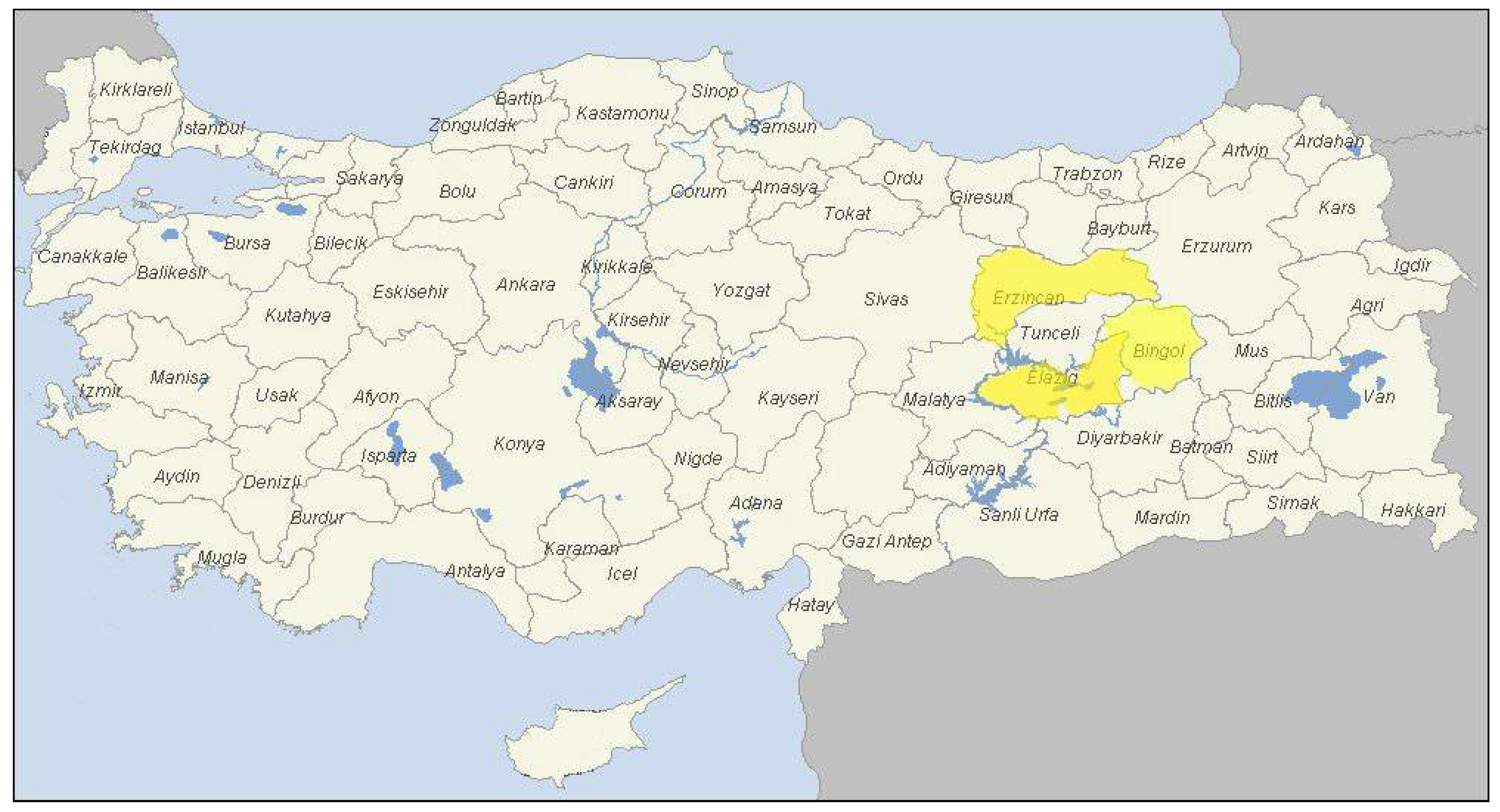
4.1. Study Area and Sample Collection
4.2. Genomic DNA Isolation from Individual Cyst Samples
4.3. PCR Amplification of mt-CO1 Gene Fragment
4.4. DNA Sequence Analysis
4.5. Alignment and Phylogenetic Analysis of mt-CO1 Gene
4.6. Haplotype Networks, Nucleotide Polymorphism, Diversity, and Neutrality Indices
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Romig, T.; Deplazes, P.; Jenkins, D.; Giraudoux, P.; Massolo, A.; Craig, P.S. Ecology and life cycle patterns of Echinococcus species. Adv. Parasitol. 2017, 95, 213–314. [Google Scholar] [PubMed]
- Budke, C.M.; Deplazes, P.; Torgerson, P.R. Global socioeconomic impact of cystic echinococcosis. Emerg. Infec. Dis. 2006, 12, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Gessese, A.T. Review on epidemiology and public health significance of hydatidosis. Vet. Med. Int. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sharbatkhori, M.; Tanzif, A.; Rostami, S.; Rostami, M.; Harandi, M.F. Echinococcus granulosus sensu lato genotypes in domestic livestock and humans in Golestan Province, Iran. Rev. Inst. Med. Trop. Sao. Paulo. 2016, 58, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuitton, D.A.; McManus, D.P.; Rogan, M.T.; Romig, T.; Gottstein, B.; Naidich, A. International consensus on terminology to be used in the field of echinococcoses. Parasite 2020, 27, 41. [Google Scholar] [CrossRef]
- Deplazes, P.; Rinaldi, L.; Alvarez Rojas, C.A.; Torgerson, P.R.; Harandi, M.F.; Romig, T.; Antolova, D.; Schurer, J.M.; Lahmar, S.; Cringoli, G.; et al. Global distribution of alveolar and cystic echinococcosis. Adv. Parasitol. 2017, 95, 315–493. [Google Scholar] [CrossRef] [Green Version]
- Borhani, M.; Fathi, S.; Darabi, E.; Jalousian, F.; Simsek, S.; Ahmed, H.; Kesik, H.K.; Hosseini, S.H.; Romig, T.; Harandi, M.F.; et al. Echinococcoses in Iran, Turkey, and Pakistan: Old diseases in the new millennium. Clin. Microbiol. Rev. 2021, 34, e00290-20. [Google Scholar] [CrossRef]
- Köse, M.; Sevimli, F.K. Prevalence of cystic echinococcosis in slaughtered cattle in Afyonkarahisar. Turkiye Parazitol. Derg. 2008, 32, 27–30. [Google Scholar]
- Simsek, S.; Eroksuz, Y. Occurrence and molecular characterization of Echinococcus granulosus in Turkish mouflon (Ovis gmelinii anatolica). Acta. Trop. 2009, 109, 167–169. [Google Scholar] [CrossRef]
- Balkaya, I.; Simsek, S. Prevalence and economic importance of hydatidosis and fasciolosis in slaughtered cattle in Erzurum province of Turkey. Kafkas Univ. Vet. Fak. Derg. 2010, 16, 793–797. [Google Scholar] [CrossRef]
- Simsek, S.; Balkaya, I.; Koroglu, E. Epidemiological survey and molecular characterization of Echinococcus granulosus in cattle in an endemic area of eastern Turkey. Vet. Parasitol. 2010, 172, 347–349. [Google Scholar] [CrossRef] [PubMed]
- Beyhan, Y.E.; Umur, S. Molecular characterization and prevalence of cystic echinococcosis in slaughtered water buffaloes in Turkey. Vet. Parasitol. 2011, 181, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Oguz, B.; Deger, S. Cystic echinococcosis and cysticerci of Taenia hydatigena in cattle and sheep slaughtered in a van local slaughterhouse. Turkiye Parazitol. Derg. 2013, 37, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Sariozkan, S.; Yalcin, C. Estimating the production losses due to cystic echinococcosis in ruminants in Turkey. Vet. Parasitol. 2009, 163, 330–334. [Google Scholar] [CrossRef]
- Mehmood, S.; Simsek, S.; Celik, F.; Kesik, H.K.; Kilinc, S.G.; Haroon, A. Molecular survey on cattle and sheep hydatidosis and first detection of Echinococcus canadensis (G6/G7) in sheep in Turkey. Parasitology 2020, 147, 1055–1062. [Google Scholar] [CrossRef]
- Kesik, H.K.; Simsek, S.; Kilinc, S.G.; Koroglu, E. Identifcation of antigen B (AgB) Gene polymorphism in cattle and sheep isolates of Echinococcus granulosus and investigation of effects on serological diagnosis. Acta. Trop. 2019, 199, 105099. [Google Scholar] [CrossRef]
- Higuita, N.I.A.; Brunetti, E.; McCloskey, C. Cystic echinococcosis. J. Clin. Micro. 2016, 54, 518–523. [Google Scholar] [CrossRef] [Green Version]
- Nakao, M.; McManus, D.P.; Schantz, P.M.; Craig, P.S.; Ito, A.A. Molecular phylogeny of the genus Echinococcus inferred from complete mitochondrial genomes. Parasitology 2007, 134, 713–722. [Google Scholar] [CrossRef] [Green Version]
- Kinkar, L.; Laurimäe, T.; Acosta-Jamett, G.; Andresiuk, V.; Balkaya, I.; Casulli, A.; Gasser, R.B.; van der Giessen, J.; Gonzalez, L.M.; Haag, K.L.; et al. Global phylogeography and genetic diversity of the zoonotic tapeworm Echinococcus granulosus sensu stricto genotype G1. Int. J. Parasitol. 2018, 48, 729–742. [Google Scholar] [CrossRef]
- Yanagida, T.; Mohammadzadeh, T.; Kamhawi, S.; Nakao, M.; Sadjjadi, S.M.; Hijjawi, N.; Abdel-Hafez, S.K.; Sako, Y.; Okamoto, M.; Ito, A. Genetic polymorphisms of Echinococcus granulosus sensu stricto in the Middle East. Parasitol. Int. 2012, 61, 599–603. [Google Scholar] [CrossRef]
- Casulli, A.; Interisano, M.; Sreter, T.; Chitimia, L.; Kirkova, Z.; La Rosa, G.; Pozio, E. Genetic variability of Echinococcus granulosus sensu stricto in Europe inferred by mitochondrial DNA sequences. Infect. Gen. Evol. 2012, 12, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Kinkar, L.; Laurimäe, T.; Simsek, S.; Balkaya, I.; Casulli, A.; Manfredi, M.T.; Ponce-Gordo, F.; Varcasia, A.; Lavikainen, A.; Gonzalez, L.M.; et al. High-resolution phylogeography of zoonotic tapeworm Echinococcus granulosus sensu stricto genotype G1 with an emphasis on its distribution in Turkey, Italy and Spain. Parasitology 2016, 143, 1790–1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesik, H.K.; Kilinc, S.G.; Simsek, S.; Gul, A. Occurrence of liver hydatid cysts in a donkey and molecular characterization of Echinococcus equinus. J. Parasitol. 2019, 105, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Laurimäe, T.; Kinkar, L.; Andresiuk, V.; Haag, K.L.; Ponce-Gordo, F.; Acosta-Jamett, G.; Garate, T.; Gonzalez, L.M.; Saarma, U. Genetic diversity and phylogeography of highly zoonotic Echinococcus granulosus genotype G1 in the Americas (Argentina, Brazil, Chile and Mexico) based on 8279 bp of mtDNA. Infec. Genet. Evol. 2016, 45, 290–296. [Google Scholar] [CrossRef]
- Rojas, C.A.A.; Ebi, D.; Paredes, R.; Acosta-Jamett, G.; Urriola, N.; Roa, J.C.; Manterola, C.; Cortes, S.; Romig, T.; Scheerlinck, J.P.; et al. High intraspecifc variability of Echinococcus granulosus sensu stricto in Chile. Parasitol. Int. 2017, 66, 112–115. [Google Scholar] [CrossRef]
- Boufana, B.; Lahmar, S.; Rebaï, W.; Ben Safta, Z.; Jebabli, L.; Ammar, A.; Kachti, M.; Aouadi, S.; Craig, P.S. Genetic variability and haplotypes of Echinococcus isolates from Tunisia. Trans. R. Soc. Trop. Med. Hyg. 2014, 108, 706–714. [Google Scholar] [CrossRef] [Green Version]
- Avcioglu, H.; Guven, E.; Balkaya, I.; Kirman, R.; Akyuz, M.; Bia, M.M.; Gulbeyen, H.; Yaya, S. The situation of echinococcosis in stray dogs in Turkey: The first finding of Echinococcus multilocularis and Echinococcus ortleppi. Parasitology 2021, 148, 1092–1098. [Google Scholar] [CrossRef]
- Kesik, H.K.; Celik, F.; Simsek, S.; Kilinc, S.G.; Ahmed, H.; Gul, A. Molecular characterization and haplotype analyses of lung hydatid cyst isolates of cattle and frst report of Echinococcus canadensis (G6/G7) in cattle isolates in Turkey. Acta. Parasitol. 2021, 66, 1538–1547. [Google Scholar] [CrossRef]
- Macin, S.; Orsten, S.; Samadzade, R.; Colak, B.; Cebeci, H.; Fındık, D. Human and animal cystic echinococcosis in Konya, Turkey: Molecular identification and the first report of E. equinus from human host in Turkey. Parasitol. Res. 2021, 120, 563–568. [Google Scholar] [CrossRef]
- Mehmood, N.; Dessì, G.; Ahmed, F.; Joanny, G.; Tamponi, C.; Cappai, M.G.; Varcasia, A.; Scala, A. Genetic diversity and transmission patterns of Echinococcus granulosus sensu stricto among domestic ungulates of Sardinia, Italy. Parasitol. Res. 2021, 120, 2533–2542. [Google Scholar] [CrossRef]
- Nakao, M.; Sako, Y.; Yokoyama, N.; Fukunaga, M.; Ito, A. Mitochondrial genetic code in cestodes. Mol. Biochem. Parasitol. 2000, 111, 415–424. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; LI, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-delbarrio, J.C.; Guirao-rico, S.; Librado, P.; Ramos-onsins, S.E.; Sánchez-gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics. 1989, 123, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods. Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Nucleotide Positions | 6 | 40 | 112 | 142 | 143 | 159 | 180 | 271 | 302 | 307 | 355 | 358 | 361 | 377 | 383 | 402 | 422 | 423 | 428 | 430 | 438 | 448 | 458 | 461 | |
| NC_044548 (Reference sequence) | C | C | T | T | C | T | C | C | C | A | C | T | T | A | C | G | A | G | A | T | C | C | T | T | |
| Hap_01 | T | T | T | ||||||||||||||||||||||
| Hap_02 | T | T | |||||||||||||||||||||||
| Hap_03 | T | ||||||||||||||||||||||||
| Hap_04 | C | T | |||||||||||||||||||||||
| Hap_05 | T | C | |||||||||||||||||||||||
| Hap_06 | |||||||||||||||||||||||||
| Hap_07 | |||||||||||||||||||||||||
| Hap_08 | |||||||||||||||||||||||||
| Hap_09 | T | T | |||||||||||||||||||||||
| Hap_10 | A | ||||||||||||||||||||||||
| Hap_11 | T | G | |||||||||||||||||||||||
| Hap_12 | T | T | |||||||||||||||||||||||
| Hap_13 | C | G | |||||||||||||||||||||||
| Hap_14 | |||||||||||||||||||||||||
| Hap_15 | T | G | |||||||||||||||||||||||
| Hap_16 | T | C | T | ||||||||||||||||||||||
| Hap_17 | T | ||||||||||||||||||||||||
| Hap_18 | T | ||||||||||||||||||||||||
| Hap_19 | T | G | T | ||||||||||||||||||||||
| Hap_20 | T | ||||||||||||||||||||||||
| Hap_21 | T | ||||||||||||||||||||||||
| Hap_22 | |||||||||||||||||||||||||
| Hap_23 | |||||||||||||||||||||||||
| Hap_24 | T | C | |||||||||||||||||||||||
| Hap_25 | T | ||||||||||||||||||||||||
| Hap_26 | C | ||||||||||||||||||||||||
| Hap_27 | T | A | |||||||||||||||||||||||
| Hap_28 | |||||||||||||||||||||||||
| Hap_29 | |||||||||||||||||||||||||
| Hap_30 | T | A | T | ||||||||||||||||||||||
| Hap_31 | T | T | T | ||||||||||||||||||||||
| Hap_32 | A | C | A | G | T | G | |||||||||||||||||||
| Hap_33 | A | ||||||||||||||||||||||||
| Nucleotide Positions | 479 | 480 | 482 | 484 | 486 | 493 | 497 | 498 | 499 | 500 | 502 | 503 | 504 | 505 | 506 | 507 | 511 | 513 | 515 | 518 | 522 | 523 | 525 | 527 | 528 |
| NC_044548 (Ref. sequence) | G | T | T | T | T | C | G | T | T | T | G | G | G | T | C | A | T | T | T | A | T | T | G | T | T |
| Hap_01 | |||||||||||||||||||||||||
| Hap_02 | C | C | |||||||||||||||||||||||
| Hap_03 | |||||||||||||||||||||||||
| Hap_04 | |||||||||||||||||||||||||
| Hap_05 | A | A | A | C | A | A | C | ||||||||||||||||||
| Hap_06 | |||||||||||||||||||||||||
| Hap_07 | A | ||||||||||||||||||||||||
| Hap_08 | C | T | |||||||||||||||||||||||
| Hap_09 | |||||||||||||||||||||||||
| Hap_10 | T | A | A | A | A | A | |||||||||||||||||||
| Hap_11 | |||||||||||||||||||||||||
| Hap_12 | A | A | A | A | C | A | |||||||||||||||||||
| Hap_13 | |||||||||||||||||||||||||
| Hap_14 | T | G | |||||||||||||||||||||||
| Hap_15 | |||||||||||||||||||||||||
| Hap_16 | |||||||||||||||||||||||||
| Hap_17 | |||||||||||||||||||||||||
| Hap_18 | |||||||||||||||||||||||||
| Hap_19 | |||||||||||||||||||||||||
| Hap_20 | C | A | A | A | T | ||||||||||||||||||||
| Hap_21 | A | ||||||||||||||||||||||||
| Hap_22 | T | ||||||||||||||||||||||||
| Hap_23 | T | ||||||||||||||||||||||||
| Hap_24 | |||||||||||||||||||||||||
| Hap_25 | C | ||||||||||||||||||||||||
| Hap_26 | A | A | A | A | C | A | |||||||||||||||||||
| Hap_27 | C | A | A | A | A | ||||||||||||||||||||
| Hap_28 | A | ||||||||||||||||||||||||
| Hap_29 | C | G | A | A | A | ||||||||||||||||||||
| Hap_30 | |||||||||||||||||||||||||
| Hap_31 | C | A | |||||||||||||||||||||||
| Hap_32 | A | A | |||||||||||||||||||||||
| Hap_33 | |||||||||||||||||||||||||
| No | Haplotype Name | Number of Isolates | Isolate Codes (Accession Numbers) |
|---|---|---|---|
| 1 | Hap_01 | 1 | BNG41(MW020975) |
| 2 | Hap_02 | 1 | BNG42(MW020976) |
| 3 | Hap_03 | 2 | BNG43(MW020977), BNG96(MW021030) |
| 4 | Hap_04 | 1 | BNG44(MW020978) |
| 5 | Hap_05 | 1 | BNG45(MW020979) |
| 6 | Hap_06 | 17 | BNG46(MW020980), BNG55(MW020989), BNG59(MW020993), BNG61(MW020995), BNG62(MW020996), BNG73(MW021007), BNG76(MW021010), BNG81(MW021015), BNG84(MW021018), BNG85(MW021019), BNG91(MW021025), BNG93(MW021027), BNG94(MW021028), BNG95(MW021029), BNG97(MW021031), BNG98(MW021032), BNG99(MW021033) |
| 7 | Hap_07 | 1 | BNG47 (MW020981) |
| 8 | Hap_08 | 1 | BNG48 (MW020982) |
| 9 | Hap_09 | 3 | BNG49(MW020983), BNG87(MW021021), BNG89(MW021023) |
| 10 | Hap_10 | 1 | BNG50(MW020984) |
| 11 | Hap_11 | 1 | BNG51(MW020985) |
| 12 | Hap_12 | 1 | BNG52(MW020986) |
| 13 | Hap_13 | 1 | BNG53(MW020987) |
| 14 | Hap_14 | 1 | BNG54(MW020988) |
| 15 | Hap_15 | 1 | BNG56(MW020990) |
| 16 | Hap_16 | 1 | BNG57(MW020991) |
| 17 | Hap_17 | 2 | BNG58(MW020992), BNG60 (MW020994) |
| 18 | Hap_18 | 7 | BNG63(MW020997), BNG64(MW020998), BNG66(MW021000), BNG70(MW021004), BNG74(MW021008), BNG82(MW021016), BNG83(MW021017) |
| 19 | Hap_19 | 1 | BNG65(MW020999) |
| 20 | Hap_20 | 1 | BNG67(MW021001) |
| 21 | Hap_21 | 1 | BNG68(MW021002) |
| 22 | Hap_22 | 1 | BNG69(MW021003) |
| 23 | Hap_23 | 1 | BNG71(MW021005) |
| 24 | Hap_24 | 1 | BNG72(MW021006) |
| 25 | Hap_25 | 2 | BNG75(MW021009), BNG100(MW021034) |
| 26 | Hap_26 | 1 | BNG77(MW021011) |
| 27 | Hap_27 | 1 | BNG78(MW021012) |
| 28 | Hap_28 | 1 | BNG79(MW021013) |
| 29 | Hap_29 | 1 | BNG80(MW021014) |
| 30 | Hap_30 | 1 | BNG86(MW021020) |
| 31 | Hap_31 | 1 | BNG88(MW021022) |
| 32 | Hap_32 | 1 | BNG90(MW021024) |
| 33 | Hap_33 | 1 | BNG92(MW021026) |
| mtDNA | n | hn | hd ± SD | πd ± SD | Tajima’s D | p Value | Fu’s Fs | p Value | FLD | p Value | FLF | p Value |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 530 bp | 60 | 33 | 0.908 ± 0.030 | 0.00692 ± 0.00101 | −2.35657 | p < 0.01 | −27.276 | 0.0000 | −3.51234 | p < 0.02 | −3.67712 | p < 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kesik, H.K.; Celik, F.; Kilinc, S.G.; Simsek, S.; Ahmed, H.; Shen, Y.; Cao, J. Genetic Diversity and Haplotype Analysis of Cattle Hydatid Cyst Isolates Using Mitochondrial Markers in Turkey. Pathogens 2022, 11, 519. https://doi.org/10.3390/pathogens11050519
Kesik HK, Celik F, Kilinc SG, Simsek S, Ahmed H, Shen Y, Cao J. Genetic Diversity and Haplotype Analysis of Cattle Hydatid Cyst Isolates Using Mitochondrial Markers in Turkey. Pathogens. 2022; 11(5):519. https://doi.org/10.3390/pathogens11050519
Chicago/Turabian StyleKesik, Harun Kaya, Figen Celik, Seyma Gunyakti Kilinc, Sami Simsek, Haroon Ahmed, Yujuan Shen, and Jianping Cao. 2022. "Genetic Diversity and Haplotype Analysis of Cattle Hydatid Cyst Isolates Using Mitochondrial Markers in Turkey" Pathogens 11, no. 5: 519. https://doi.org/10.3390/pathogens11050519