
H. pylori Infection and Virulence Factors cagA and vacA (s and m Regions) in Gastric Adenocarcinoma from Pará State, Brazil

 ,
, 
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takahashi-Kanemitsu, A.; Knight, C.T.; Hatakeyama, M. Molecular anatomy and pathogenic actions of Helicobacter pylori CagA that underpin gastric carcinogenesis. Cell. Mol. Immunol. 2020, 17, 50–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, I.; Araya, P.; Rojas, A. Helicobacter pylori Infection and Lung Cancer: New Insights and Future Challenges. Chin. J. Lung Cancer 2018, 21, 658–662. [Google Scholar] [CrossRef]
- Joo, M. Rare Gastric Lesions Associated with Helicobacter pylori Infection: A Histopathological Review. J. Pathol. Transl. Med. 2017, 51, 341–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poorolajal, J.; Moradi, L.; Mohammadi, Y.; Cheraghi, Z.; Gohari-Ensaf, F. Risk factors for stomach cancer: A systematic review and meta-analysis. Epidemiol. Health 2020, 42, e2020004. [Google Scholar] [CrossRef]
- Palamides, P.; Jolaiya, T.; Idowu, A.; Loell, E.; Onyekwere, C.; Ugiagbe, R.; Agbo, I.; Lesi, O.; Ndububa, D.; Adekanle, O.; et al. Helicobacter pylori patient isolates from South Africa and Nigeria differ in virulence factor pathogenicity profile and associated gastric disease outcome. Sci. Rep. 2020, 10, 11409. [Google Scholar] [CrossRef]
- Mucito-Varela, E.; Castillo-Rojas, G.; Calva, J.J.; López-Vidal, Y. Integrative and Conjugative Elements of Helicobacter pylori Are Hypothetical Virulence Factors Associated with Gastric Cancer. Front. Cell. Infect. Microbiol. 2020, 10, 525335. [Google Scholar] [CrossRef]
- Sharndama, H.C.; Mba, I.E. Helicobacter pylori: An up-to-date overview on the virulence and pathogenesis mechanisms. Braz. J. Microbiol. 2022, 53, 33–50. [Google Scholar] [CrossRef]
- Canzian, F.; Rizzato, C.; Obazee, O.; Stein, A.; Flores-Luna, L.; Camorlinga-Ponce, M.; Mendez-Tenorio, A.; Vivas, J.; Trujillo, E.; Jang, H.; et al. Genetic polymorphisms in the cag pathogenicity island of Helicobacter pylori and risk of stomach cancer and high-grade premalignant gastric lesions. Int. J. Cancer 2020, 147, 2437–2445. [Google Scholar] [CrossRef]
- Lima, V.P.; Silva-Fernandes, I.J.d.L.; Alves, M.K.S.; Rabenhorst, S.H.B. Prevalence of Helicobacter pylori genotypes (vacA, cagA, cagE and virB11) in gastric cancer in Brazilian’s patients: An association with histopathological parameters. Cancer Epidemiol. 2011, 35, e32–e37. [Google Scholar] [CrossRef] [Green Version]
- Silva, B.; Nunes, A.; Vale, F.F.; Rocha, R.; Gomes, J.P.; Dias, R.; Oleastro, M. The expression of Helicobacter pylori tfs plasticity zone cluster is regulated by pH and adherence, and its composition is associated with differential gastric IL-8 secretion. Helicobacter 2017, 22, e12390. [Google Scholar] [CrossRef]
- Foegeding, N.J.; Caston, R.R.; McClain, M.S.; Ohi, M.D.; Cover, T.L. An Overview of Helicobacter pylori VacA Toxin Biology. Toxins 2016, 8, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shetty, V.; Lingadakai, R.; Pai, G.C.; Ballal, M. Profile of Helicobacter pylori cagA & vacA genotypes and its association with the spectrum of gastroduodenal disease. Indian J. Med. Microbiol. 2021, 39, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Oikawa, R.; Kodaka, Y.; Sato, Y.; Ono, S.; Kenmochi, T.; Suzuki, H.; Futagami, S.; Kato, M.; Yamamoto, H.; et al. Cancer-related genetic variants of Helicobacter pylori strains determined using gastric wash-based whole-genome analysis with single-molecule real-time technology. Int. J. Cancer 2021, 148, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Boonyanugomol, W.; Kongkasame, W.; Palittapongarnpim, P.; Baik, S.-C.; Jung, M.-H.; Shin, M.-K.; Kang, H.-L.; Lee, W.-K. Genetic variation in the cag pathogenicity island of Helicobacter pylori strains detected from gastroduodenal patients in Thailand. Braz. J. Microbiol. 2020, 51, 1093–1101. [Google Scholar] [CrossRef]
- Tegtmeyer, N.; Neddermann, M.; Asche, C.I.; Backert, S. Subversion of host kinases: A key network in cellular signaling hijacked by Helicobacter pylori CagA. Mol. Microbiol. 2017, 105, 358–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagase, L.; Hayashi, T.; Senda, T.; Hatakeyama, M. Dramatic increase in SHP2 binding activity of Helicobacter pylori Western CagA by EPIYA-C duplication: Its implications in gastric carcinogenesis. Sci. Rep. 2015, 5, 15749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tagoe, E.A.; Awandare, G.A.; Quaye, O.; Asmah, R.H.; Archampong, T.N.; Osman, M.A.; Brown, C.A. Helicobacter pylori Variants with ABC-Type Tyrosine Phosphorylation Motif in Gastric Biopsies of Ghanaian Patients. BioMed Res. Int. 2021, 2021, 6616059. [Google Scholar] [CrossRef]
- Liu, B.; Li, X.; Sun, F.; Tong, X.; Bai, Y.; Jin, K.; Liu, L.; Dai, F.; Li, N. HP-CagA+ Regulates the Expression of CDK4/CyclinD1 via reg3 to Change Cell Cycle and Promote Cell Proliferation. Int. J. Mol. Sci. 2019, 21, 224. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Senda, M.; Suzuki, N.; Nishikawa, H.; Ben, C.; Tang, C.; Nagase, L.; Inoue, K.; Senda, T.; Hatakeyama, M. Differential Mechanisms for SHP2 Binding and Activation Are Exploited by Geographically Distinct Helicobacter pylori CagA Oncoproteins. Cell Rep. 2017, 20, 2876–2890. [Google Scholar] [CrossRef] [Green Version]
- Fazeli, Z.; Alebouyeh, M.; Tavirani, M.R.; Azimirad, M.; Yadegar, A. Helicobacter pylori CagA induced interleukin-8 secretion in gastric epithelial cells. Gastroenterol. Hepatol. Bed Bench 2016, 9, S42–S46. [Google Scholar]
- Ferreira, R.; Pinto-Ribeiro, I.; Wen, X.; Marcos-Pinto, R.; Dinis-Ribeiro, M.; Carneiro, F.; Figueiredo, C. Helicobacter pylori cagA Promoter Region Sequences Influence CagA Expression and Interleukin 8 Secretion. J. Infect. Dis. 2016, 213, 669–673. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Lim, J.W.; Kim, H. Astaxanthin Inhibits Mitochondrial Dysfunction and Interleukin-8 Expression in Helicobacter pylori-Infected Gastric Epithelial Cells. Nutrients 2018, 10, 1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, S.; Lim, J.; Kim, H. β-Carotene Inhibits Expression of Matrix Metalloproteinase-10 and Invasion in Helicobacter pylori-Infected Gastric Epithelial Cells. Molecules 2021, 26, 1567. [Google Scholar] [CrossRef]
- Dadashzadeh, K.; Peppelenbosch, M.; Adamu, A.I. Helicobacter pylori Pathogenicity Factors Related to Gastric Cancer. Can. J. Gastroenterol. Hepatol. 2017, 2017, e7942489. [Google Scholar] [CrossRef] [Green Version]
- Pyburn, T.M.; Foegeding, N.J.; González-Rivera, C.; McDonald, N.A.; Gould, K.L.; Cover, T.L.; Ohi, M.D. Structural organization of membrane-inserted hexamers formed by Helicobacter pylori VacA toxin. Mol. Microbiol. 2016, 102, 22–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricci, V. Relationship between VacA Toxin and Host Cell Autophagy in Helicobacter pylori Infection of the Human Stomach: A Few Answers, Many Questions. Toxins 2016, 8, 203. [Google Scholar] [CrossRef] [Green Version]
- McClain, M.S.; Czajkowsky, D.M.; Torres, V.J.; Szabo, G.; Shao, Z.; Cover, T.L. Random Mutagenesis of Helicobacter pylori vacA to Identify Amino Acids Essential for Vacuolating Cytotoxic Activity. Infect. Immun. 2006, 74, 6188–6195. [Google Scholar] [CrossRef] [Green Version]
- McClain, M.S.; Iwamoto, H.; Cao, P.; Vinion-Dubiel, A.D.; Li, Y.; Szabo, G.; Shao, Z.; Cover, T. Essential Role of a GXXXG Motif for Membrane Channel Formation by Helicobacter pylori Vacuolating Toxin. J. Biol. Chem. 2003, 278, 12101–12108. [Google Scholar] [CrossRef] [Green Version]
- Bakhti, S.Z.; Latifi-Navid, S.; Zahri, S. Unique constellations of five polymorphic sites of Helicobacter pylori vacA and cagA status associated with risk of gastric cancer. Infect. Genet. Evol. 2020, 79, 104167. [Google Scholar] [CrossRef]
- Pormohammad, A.; Ghotaslou, R.; Leylabadlo, H.E.; Nasiri, M.J.; Dabiri, H.; Hashemi, A. Risk of gastric cancer in association with Helicobacter pylori different virulence factors: A systematic review and meta-analysis. Microb. Pathog. 2018, 118, 214–219. [Google Scholar] [CrossRef]
- Lee, D.-H.; Ha, J.-H.; Shin, J.-I.; Kim, K.-M.; Choi, J.-G.; Park, S.; Park, J.-S.; Seo, J.-H.; Park, J.-S.; Shin, M.-K.; et al. Increased Risk of Severe Gastric Symptoms by Virulence Factors vacAs1c, alpA, babA2, and hopZ in Helicobacter pylori Infection. J. Microbiol. Biotechnol. 2021, 31, 368–379. [Google Scholar] [CrossRef]
- Nejati, S.; Karkhah, A.; Darvish, H.; Validi, M.; Ebrahimpour, S.; Nouri, H.R. Influence of Helicobacter pylori virulence factors CagA and VacA on pathogenesis of gastrointestinal disorders. Microb. Pathog. 2018, 117, 43–48. [Google Scholar] [CrossRef] [PubMed]
- de Martel, C.; Georges, D.; Bray, F.; Ferlay, J.; Clifford, G.M. Global burden of cancer attributable to infections in 2018: A worldwide incidence analysis. Lancet Glob. Health 2020, 8, e180–e190. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, V.P.; Koehne, A.L.; Dubrulle, J.; Rodriguez, A.E.; Leverich, C.K.; Kong, V.P.; Campbell, J.S.; Pierce, R.H.; Goldenring, J.R.; Choi, E.; et al. Sustained Helicobacter pylori infection accelerates gastric dysplasia in a mouse model. Life Sci. Alliance 2021, 4, e202000967. [Google Scholar] [CrossRef] [PubMed]
- Rahman, R.; Asombang, A.W.; Ibdah, J.A. Characteristics of gastric cancer in Asia. World J. Gastroenterol. 2014, 20, 4483–4490. [Google Scholar] [CrossRef]
- Wang, R.; Chen, X.-Z. High mortality from hepatic, gastric and esophageal cancers in mainland China: 40 years of experience and development. Clin. Res. Hepatol. Gastroenterol. 2014, 38, 751–756. [Google Scholar] [CrossRef]
- Kim, J.Y.; Lee, H.S.; Kim, N.; Shin, C.M.; Lee, S.H.; Park, Y.S.; Hwang, J.-H.; Kim, J.-W.; Jeong, S.-H.; Lee, D.H.; et al. Prevalence and Clinicopathologic Characteristics of Gastric Cardia Cancer in South Korea. Helicobacter 2012, 17, 358–368. [Google Scholar] [CrossRef]
- Zaterka, S.; Eisig, J.N.; Chinzon, D.; Rothstein, W. Factors Related to Helicobacter pylori Prevalence in an Adult Population in Brazil. Helicobacter 2007, 12, 82–88. [Google Scholar] [CrossRef]
- Coelho, L.G.V.; Marinho, J.R.; Genta, R.; Ribeiro, L.T.; Passos, M.D.C.F.; Zaterka, S.; Assumpção, P.P.; Barbosa, A.A.J.; Barbuti, R.; Braga, L.L.; et al. Ivth Brazilian Consensus Conference on Helicobacter pylori Infection. Arq. Gastroenterol. 2018, 55, 97–121. [Google Scholar] [CrossRef]
- De Souza, C.R.T.; De Oliveira, K.S.; Ferraz, J.J.S.; Leal, M.F.; Calcagno, D.Q.; Seabra, A.D.; Khayat, A.S.; Montenegro, R.C.; Alves, A.P.N.N.; Assumpção, P.P.; et al. Occurrence of Helicobacter pylori and Epstein-Barr virus infection in endoscopic and gastric cancer patients from Northern Brazil. BMC Gastroenterol. 2014, 14, 179. [Google Scholar] [CrossRef] [Green Version]
- Vinagre, I.D.F.; De Queiroz, A.L.; Silva Júnior, M.R.D.; Vinagre, R.M.D.F.; Martins, L.C. Helicobacter pylori Infection in Patients with Different Gastrointestinal Diseases from Northern Brazil. Arq. Gastroenterol. 2015, 52, 266–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Larrea-Baz, N.F.; Michel, A.; Romero, B.; Pérez-Gómez, B.; Moreno, V.; Martín, V.; Dierssen-Sotos, T.; Jimenez-Moleon, J.J.; Castilla, J.; Tardon, A.; et al. Helicobacter pylori Antibody Reactivities and Colorectal Cancer Risk in a Case-control Study in Spain. Front. Microbiol. 2017, 8, 888. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, Z.; Xia, X.; Chi, J.; Li, H.; Liu, X.; Li, R.; Li, Y.; Liu, D.; Tian, D.; et al. Helicobacter pylori infection selectively increases the risk for carotid atherosclerosis in young males. Atherosclerosis 2019, 291, 71–77. [Google Scholar] [CrossRef]
- Machlowska, J.; Baj, J.; Sitarz, M.; Maciejewski, R.; Sitarz, R. Gastric Cancer: Epidemiology, Risk Factors, Classification, Genomic Characteristics and Treatment Strategies. Int. J. Mol. Sci. 2020, 21, 4012. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.S.-H.; Wong, I.O.-L.; Leung, W.K. Helicobacter pylori associated gastric intestinal metaplasia: Treatment and surveillance. World J. Gastroenterol. 2016, 22, 1311–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.-N.; Wang, Z.; Li, X.; Zhou, Z.-G. Helicobacter pylori eradication cannot reduce the risk of gastric cancer in patients with intestinal metaplasia and dysplasia: Evidence from a meta-analysis. Gastric Cancer 2015, 19, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; El-Omar, E.M. Application of molecular techniques in Helicobacter pylori detection: Limitations and improvements. Helicobacter 2021, 26, e12841. [Google Scholar] [CrossRef]
- Jiang, H.; Zhou, Y.; Liao, Q.; Ouyang, H. Helicobacter pylori infection promotes the invasion and metastasis of gastric cancer through increasing the expression of matrix metalloproteinase-1 and matrix metalloproteinase-10. Exp. Ther. Med. 2014, 8, 769–774. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.P.; Sheng, X.P.; Shuai, T.K.; Zhao, Y.X.; Li, B.; Li, Y.M. Helicobacter pylori promotes invasion and metastasis of gastric cancer by enhancing heparanase expression. World J. Gastroenterol. 2018, 24, 4565–4577. [Google Scholar] [CrossRef]
- Xie, S.; Wang, S.; Xue, L.; Middleton, D.R.S.; Guan, C.; Hao, C.; Wang, J.; Li, B.; Chen, R.; Li, X.; et al. Helicobacter pylori Is Associated with Precancerous and Cancerous Lesions of the Gastric Cardia Mucosa: Results of a Large Population-Based Study in China. Front. Oncol. 2020, 10, 205. [Google Scholar] [CrossRef] [Green Version]
- Tsai, K.-F.; Liou, J.-M.; Chen, M.-J.; Chen, C.-C.; Kuo, S.-H.; Lai, I.-R.; Yeh, K.-H.; Lin, M.-T.; Wang, H.-P.; Cheng, A.-L.; et al. Distinct Clinicopathological Features and Prognosis of Helicobacter pylori Negative Gastric Cancer. PLoS ONE 2017, 12, e0170942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Y.; Ma, L.-Q.; Bai, P.-S.; Da, R.; Sun, H.; Qi, X.-G.; Ma, J.-Q.; Zhao, R.-M.; Chen, N.-Z.; Nan, K.-J. Helicobacter pylori promotes invasion and metastasis of gastric cancer cells through activation of AP-1 and up-regulation of CACUL1. Int. J. Biochem. Cell Biol. 2013, 45, 2666–2678. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, N.; Sadeghiani, M.; Rahimian, G.; Mahsa, M.; Shafigh, M.; Rafieian-Kopaei, M.; Shirzad, H. Correlation between expression of MMP-9 and MMP-3 in Helicobacter pylori infected patients with different gastroduodenal diseases. Arab J. Gastroenterol. 2018, 19, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Sadeghiani, M.; Bagheri, N.; Shahi, H.; Reiisi, S.; Rahimian, G.; Rashidi, R.; Mahsa, M.; Shafigh, M.; Salimi, E.; Rafieian-Kopaei, M.; et al. cag Pathogenicity island-dependent upregulation of matrix metalloproteinase-7 in infected patients with Helicobacter pylori. J. Immunoass. Immunochem. 2017, 38, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.S.; Dooyema, S.D.R.; Frick-Cheng, A.; Harvey, M.L.; Suarez, G.; Loh, J.T.; McDonald, W.H.; McClain, M.S.; Peek, R.M.; Cover, T.L. Bacterial Energetic Requirements for Helicobacter pylori Cag Type IV Secretion System-Dependent Alterations in Gastric Epithelial Cells. Infect. Immun. 2020, 88, e00790-19. [Google Scholar] [CrossRef] [PubMed]
- Mansour, K.B.; Fendri, C.; Battikh, H.; Garnier, M.; Zribi, M.; Jlizi, A.; Burucoa, C. Multiple and mixed Helicobacter pylori infections: Comparison of two epidemiological situations in Tunisia and France. Infect. Genet. Evol. 2016, 37, 43–48. [Google Scholar] [CrossRef]
- Matta, A.J.; Pazos, A.J.; Bustamante-Rengifo, J.A.; Bravo, L.E. Genomic Varia-bility of Helicobacter pylori Isolates of Gastric Regions from Two Colombian Populations. World J. Gastroenterol. 2017, 23, 800–809. [Google Scholar] [CrossRef]
- Dabiri, H.; Jafari, F.; Baghaei, K.; Shokrzadeh, L.; Abdi, S.; Pourhoseingholi, M.A.; Mohammadzadeh, A. Prevalence of Helicobacter pylori vacA, cagA, cagE, oipA, iceA, babA2 and babB genotypes in Iranian dyspeptic patients. Microb. Pathog. 2017, 105, 226–230. [Google Scholar] [CrossRef]
- Safaralizadeh, R.; Dastmalchi, N.; Hosseinpourfeizi, M.; Latifi-Navid, S. Helicobacter pylori virulence factors in relation to gastrointestinal diseases in Iran. Microb. Pathog. 2017, 105, 211–217. [Google Scholar] [CrossRef]
- Draper, J.L.; Hansen, L.M.; Bernick, D.L.; Abedrabbo, S.; Underwood, J.G.; Kong, N.; Huang, B.C.; Weis, A.M.; Weimer, B.C.; van Vliet, A.H.M.; et al. Fallacy of the Unique Genome: Sequence Diversity within Single Helicobacter pylori Strains. mBio 2017, 8, e02321-16. [Google Scholar] [CrossRef] [Green Version]
- Keikha, M.; Karbalaei, M. EPIYA motifs of Helicobacter pylori cagA genotypes and gastrointestinal diseases in the Iranian population: A systematic review and meta-analysis. New Microbes New Infect. 2021, 41, 100865. [Google Scholar] [CrossRef] [PubMed]
- Gastli, N.; Allain, M.; Lamarque, D.; Abitbol, V.; Billoët, A.; Collobert, G.; Coriat, R.; Terris, B.; Kalach, N.; Raymond, J. Diagnosis of Helicobacter pylori Infection in a Routine Testing Workflow: Effect of Bacterial Load and Virulence Factors. J. Clin. Med. 2021, 10, 2755. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.J.; Kim, C.G.; Lee, J.Y.; Kim, Y.-I.; Kook, M.-C.; Park, B.; Joo, J. Family History of Gastric Cancer and Helicobacter pylori Treatment. N. Engl. J. Med. 2020, 382, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Wang, T.C. Helicobacter pylori and Gastric Cancer. Gastrointest. Endosc. Clin. N. Am. 2021, 31, 451–465. [Google Scholar] [CrossRef]
- Tsuda, M.; Asaka, M.; Kato, M.; Matsushima, R.; Fujimori, K.; Akino, K.; Kikuchi, S.; Lin, Y.; Sakamoto, N. Effect on Helicobacter pylori eradication therapy against gastric cancer in Japan. Helicobacter 2017, 22, e12415. [Google Scholar] [CrossRef] [Green Version]
- Rota, C.A.; Pereira-Lima, J.C.; Blaya, C.; Nardi, N.B. Consensus and Variable Region PCR Analysis of Helicobacter pylori 3′ Region of cagA Gene in Isolates from Individuals with or without Peptic Ulcer. J. Clin. Microbiol. 2001, 39, 606–612. [Google Scholar] [CrossRef] [Green Version]
- Palau, M.; Piqué, N.; Ramírez-Lázaro, M.; Lario, S.; Calvet, X.; Miñana-Galbis, D. Whole-Genome Sequencing and Comparative Genomics of Three Helicobacter pylori Strains Isolated from the Stomach of a Patient with Adenocarcinoma. Pathogens 2021, 10, 331. [Google Scholar] [CrossRef]
- Fan, R.; Han, X.; Gong, Y.; He, L.; Xue, Z.; Yang, Y.; Sun, L.; Fan, D.; You, Y.; Meng, F.; et al. Alterations of Fucosyltransferase Genes and Fucosylated Glycans in Gastric Epithelial Cells Infected with Helicobacter pylori. Pathogens 2021, 10, 168. [Google Scholar] [CrossRef]
- Kumar, S.; Patel, G.K.; Ghoshal, U.C. Helicobacter pylori-Induced Inflammation: Possible Factors Modulating the Risk of Gastric Cancer. Pathogens 2021, 10, 1099. [Google Scholar] [CrossRef]
- Tharmalingam, N.; Park, M.; Lee, M.H.; Woo, H.J.; Kim, H.W.; Yang, J.Y.; Rhee, K.-J.; Kim, J.-B. Piperine treatment suppresses Helicobacter pylori toxin entry in to gastric epithelium and minimizes β-catenin mediated oncogenesis and IL-8 secretion in vitro. Am. J. Transl. Res. 2016, 8, 885–898. [Google Scholar]


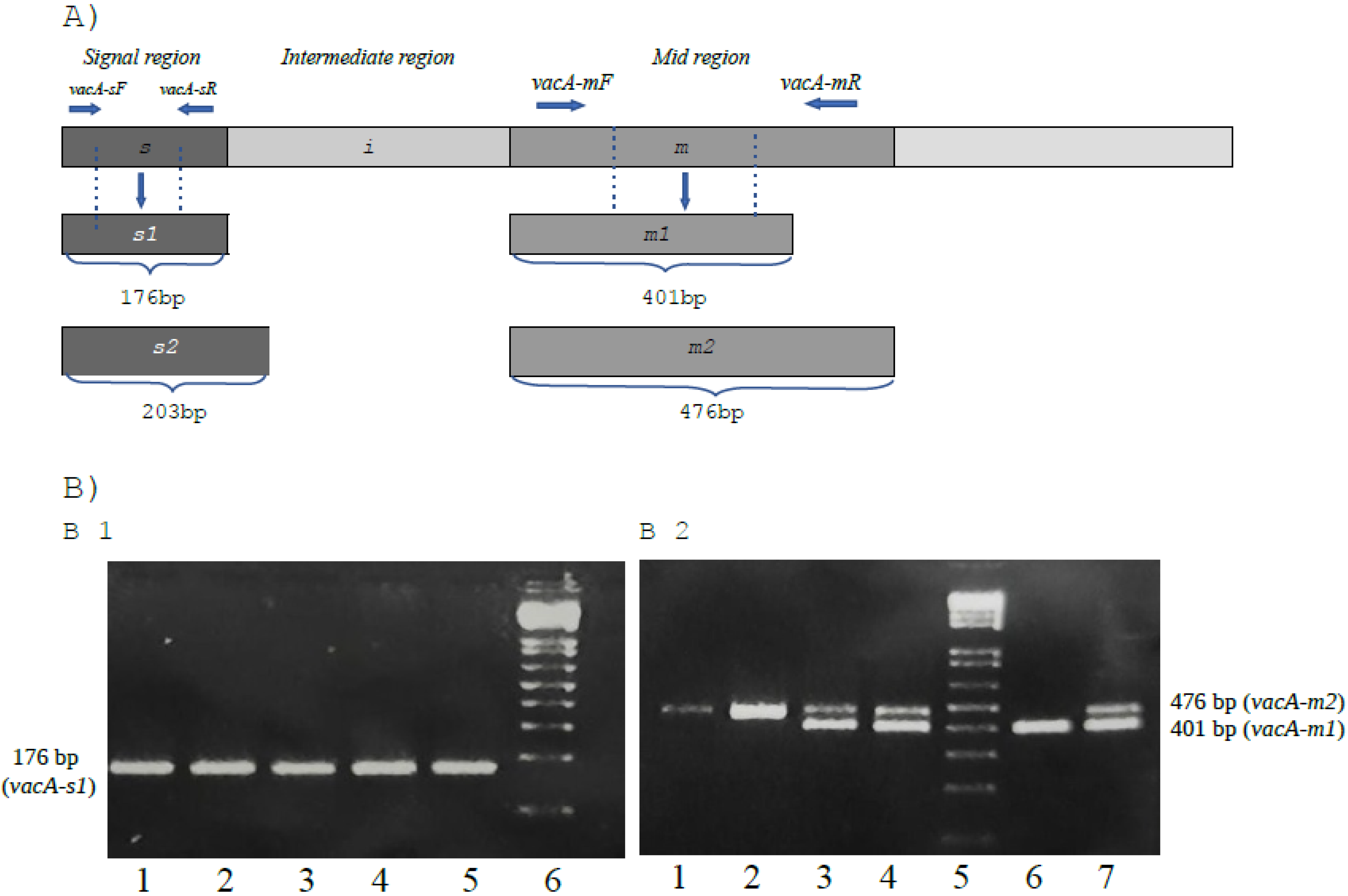{kind=link}
| Clinical and Epidemiological Variable | Positive H. pylori | Undetectable H. pylori | Value of p | |
|---|---|---|---|---|
| Age | ≥60 | 65.6% (59/90) | 48.2% (92/191) | 0.0093 |
| <60 | 34.4% (31/90) | 51.8% (99/191) | ||
| Gender | Man | 60% (54/90) | 72.8% (139/191) | 0.0437 |
| Woman | 40% (36/90) | 27.2% (52/191) | ||
| Location | Proximal | 37.8% (34/90) | 42.4% (81/191) | 0.4614 |
| Distal | 62.2% (56/90) | 57.6% (110/191) | ||
| Histological type | Intestinal | 53.3% (48/90) | 57.1% (109/191) | 0.5563 |
| Diffuse | 46.7% (42/90) | 42.9% (82/191) | ||
| Presence of metastases | presence | 59.6% (53/89) | 41.5% (78/188) | 0.0073 |
| Absent | 40.4% (36/89) | 58.5% (110/188) | ||
| Target | Primer PCR | Size (pb) | Reference |
|---|---|---|---|
| ureA | 5′-GCCAATGGTAAATTAGTT-3′ | 394 | [64] |
| 5′-CTCCTTAATTGTTTTTAC-3′ | |||
| 16SrRNA | 5′-CCCATTTGACTCAATGCGATG-3′ | 132 | [65] |
| 5′-TGGGATTAGCGAGTATGTCGG-3′ | |||
| cagA | 5′-GTGCCTGCTAGTTTGTCAGCG-3′ | 402 | [66] |
| 5′-TTGGAAACCACCTTTTGTATTAGC-3′ | |||
| vacA m1/m2 | 5′-CACAGCCACTTTCAATAACGA-3′ | 401/476 | [67] |
| 5′-CGTCAAAATAATTCCAAGGG-3′ | |||
| vacA s1/s2 | 5′-ATGGAAATACAACAAACACAC-3′ | 176/203 | [67] |
| 5′-CCTGARACCGTTCCTACAGC-3′ |
| Components/Concentrations | Volumes | |
|---|---|---|
| 16SrRNA | ureA, cagA, vacA | |
| Ultra-pure water | 14.55 μL | 16.55 μL |
| Reaction buffer 10X (Invitrogen) | 2.5 μL | 2.5 μL |
| Magnesium chloride-MgCl2 50 mM (Invitrogen) | 0.5 μL | 0.5 μL |
| Deoxynucleotides-dNTP 10 mM (Invitrogen) | 1.0 µL | 1.0 µL |
| Oligonucleotides-10 pmol/μL (Invitrogen) | 1.0 μL/1.0 μL | 1.0 μL/1.0 μL |
| Platinum-Taq DNA Polymerase 5 U/µL (Invitrogen) | 0.2 μL | 0.2 μL |
| DNA extracted from gastric biopsy | 4 μL | 2 μL |
| Final reaction volume | 25 μL | 25 μL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brasil-Costa, I.; Souza, C.d.O.; Monteiro, L.C.R.; Santos, M.E.S.; Oliveira, E.H.C.D.; Burbano, R.M.R. H. pylori Infection and Virulence Factors cagA and vacA (s and m Regions) in Gastric Adenocarcinoma from Pará State, Brazil. Pathogens 2022, 11, 414. https://doi.org/10.3390/pathogens11040414
Brasil-Costa I, Souza CdO, Monteiro LCR, Santos MES, Oliveira EHCD, Burbano RMR. H. pylori Infection and Virulence Factors cagA and vacA (s and m Regions) in Gastric Adenocarcinoma from Pará State, Brazil. Pathogens. 2022; 11(4):414. https://doi.org/10.3390/pathogens11040414
Chicago/Turabian StyleBrasil-Costa, Igor, Cintya de Oliveira Souza, Leni Célia Reis Monteiro, Maria Elisabete Silva Santos, Edivaldo Herculano Correa De Oliveira, and Rommel Mario Rodriguez Burbano. 2022. "H. pylori Infection and Virulence Factors cagA and vacA (s and m Regions) in Gastric Adenocarcinoma from Pará State, Brazil" Pathogens 11, no. 4: 414. https://doi.org/10.3390/pathogens11040414