
Ex Vivo Infection of Human Placental Explants by Trypanosoma cruzi Reveals a microRNA Profile Similar to That Seen in Trophoblast Differentiation


Abstract
:
1. Introduction
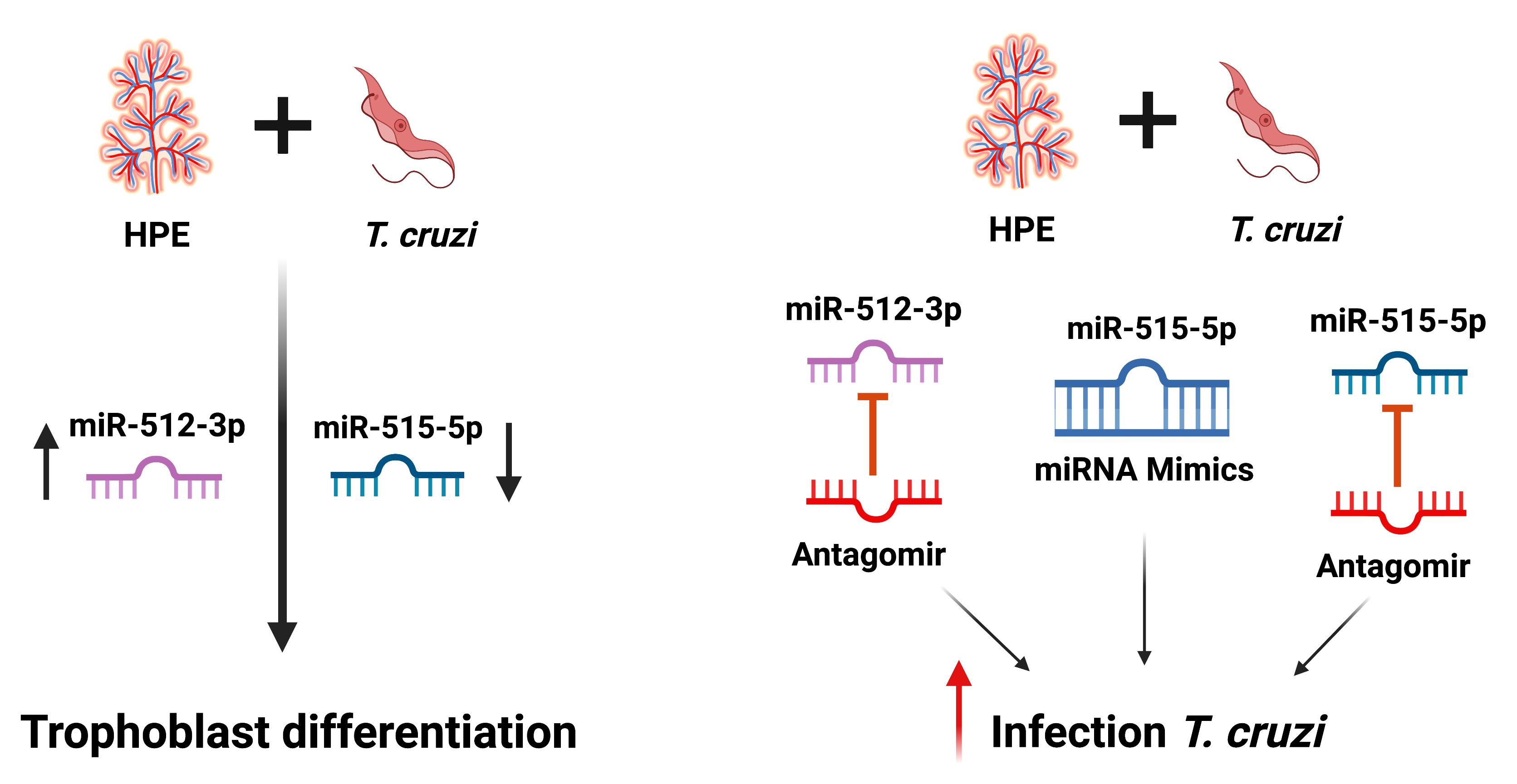
2. Results
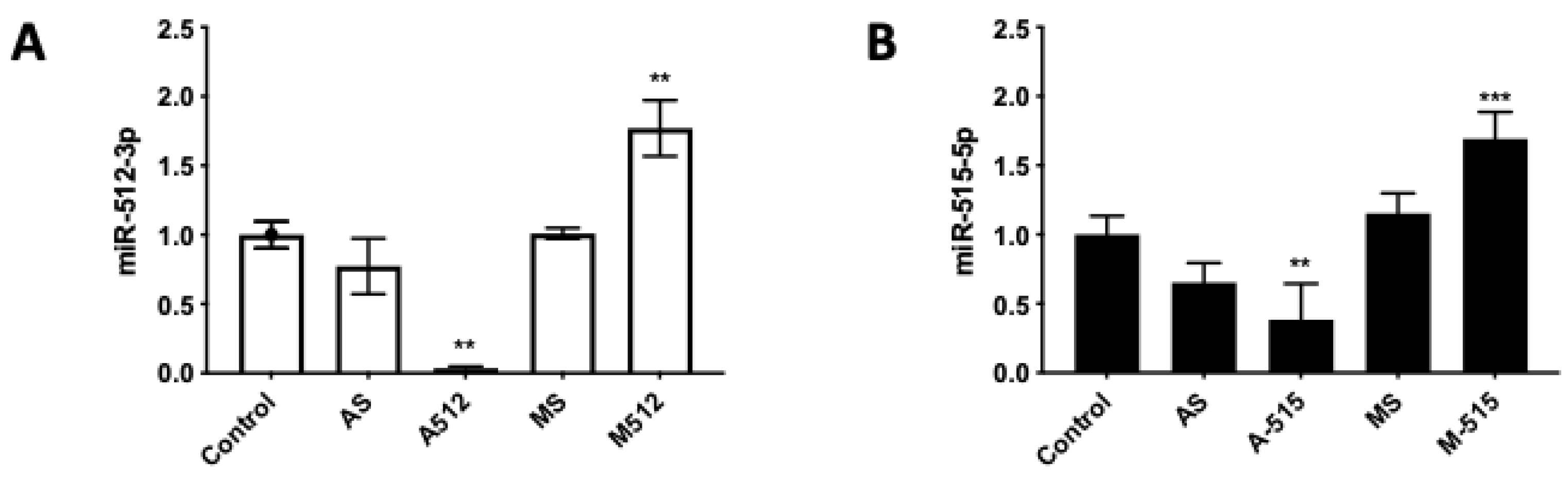
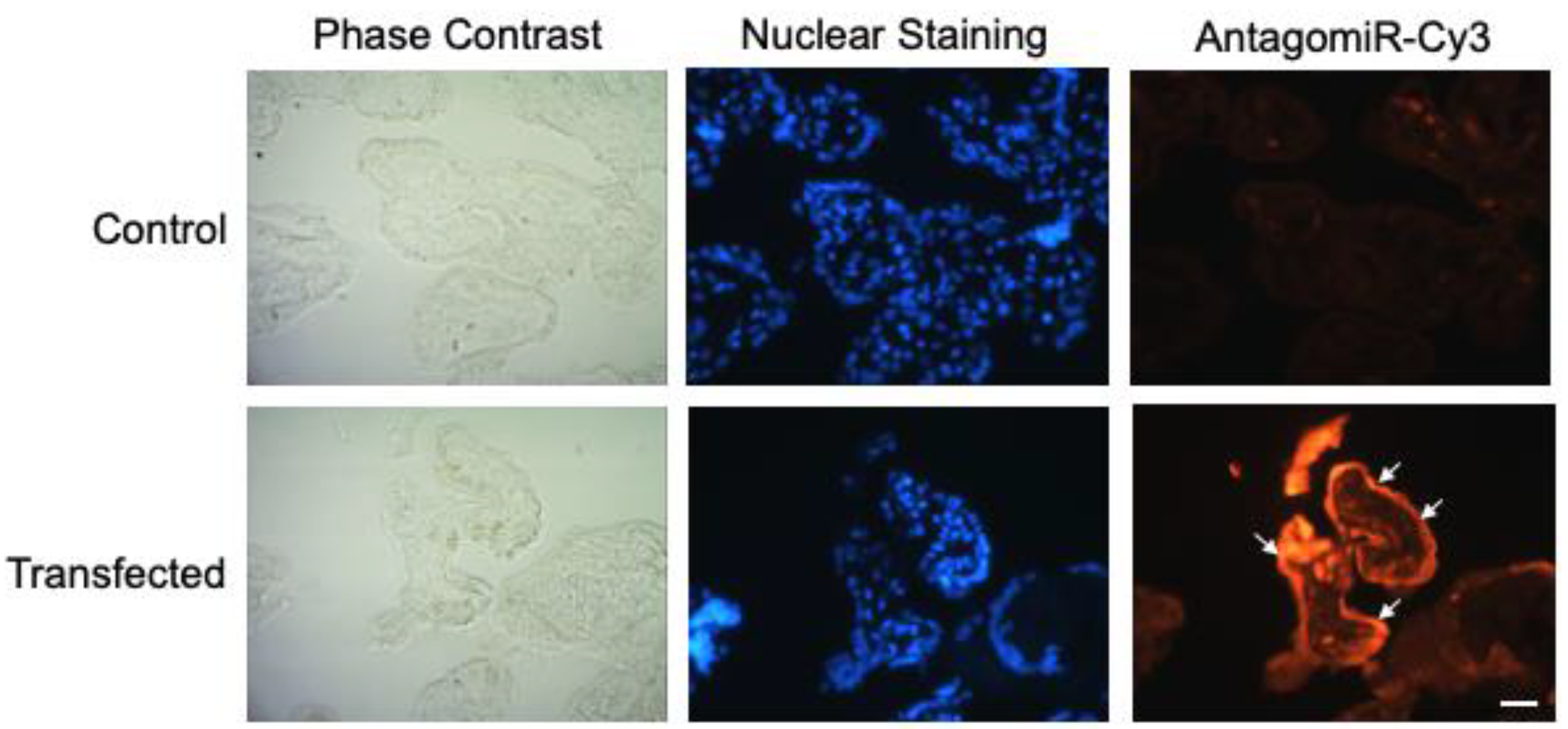
2.1. HPE Can Be Effectively Transfected with miR-512-3p and miR-515-5p Mimics and Antagomirs without Causing Tissue Damage
2.2. T. cruzi Increases and Decreases miR-512-3p and miR-515-5p Levels in HPE, Respectively
2.3. T. cruzi Does Not Affect c-FLIP Expression
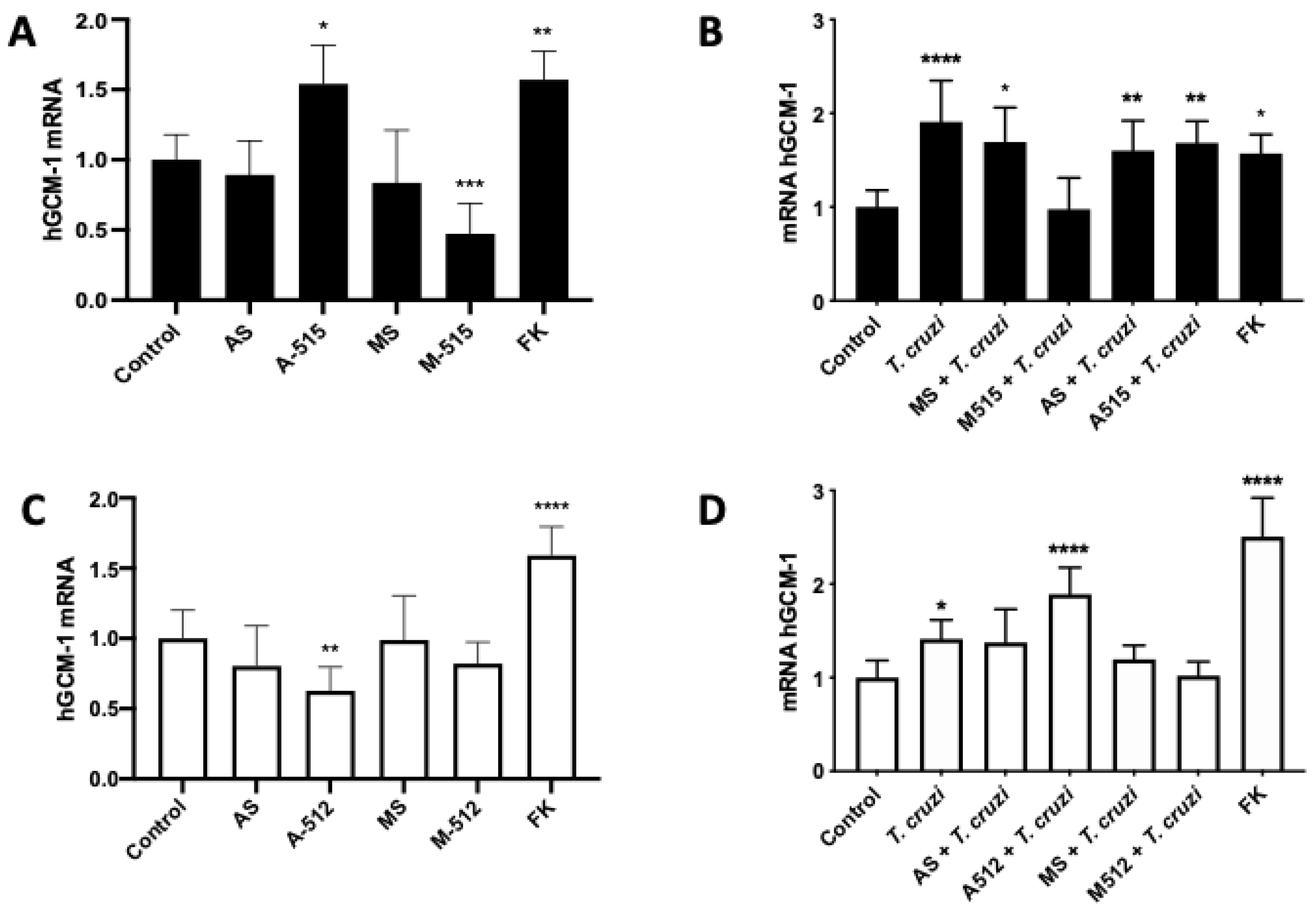
2.4. miR-515-5p and miR-512-3p Regulate T. cruzi-Induced Increase in hGCM-1
2.5. miR-512-3p and miR-515-5p Regulate T. cruzi-Induced Increase in hCG
2.6. miR-512-3p and miR-515-5p Levels Determine HPE Susceptibility to T. cruzi Infection
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Parasite Culture and Harvesting
4.3. Human Placental Explant (HPE) Culture and Parasite Infection
4.4. HPE Transfection with miRNAs Mimics and Antagomirs
4.5. Lactate Deshydrogenase Activity
4.6. Histology
4.7. miRNA and mRNA Expression Analysis via RT-qPCR
4.8. DNA Amplification via Real-Time PCR
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lenk, E.J.; Redekop, W.K.; Luyendijk, M.; Fitzpatrick, C.; Niessen, L.; Stolk, W.A.; Tediosi, F.; Rijnsburger, A.J.; Bakker, R.; Hontelez, J.A.C.; et al. Socioeconomic Benefit to Individuals of Achieving 2020 Targets for Four Neglected Tropical Diseases Controlled/Eliminated by Innovative and Intensified Disease Management: Human African Trypanosomiasis, Leprosy, Visceral Leishmaniasis, Chagas Disease. PLOS Negl. Trop. Dis. 2018, 12, e0006250. [Google Scholar] [CrossRef] [Green Version]
- Kemmerling, U.; Osuna, A.; Schijman, A.G.A.G.; Truyens, C. Congenital Transmission of Trypanosoma cruzi: A Review About the Interactions Between the Parasite, the Placenta, the Maternal and the Fetal/Neonatal Immune Responses. Front. Microbiol. 2019, 10, 1854. [Google Scholar] [CrossRef] [Green Version]
- WHO Chagas Disease (American Trypanosomiasis). Available online: https://www.who.int/health-topics/chagas-disease#tab=tab_1 (accessed on 12 December 2021).
- WHO. Chagas Disease in Latin America: An Epidemiological Update Based on 2010 Estimates. Relev. Epidemiol. Hebd. 2015, 90, 33–43. [Google Scholar]
- Picado, A.; Cruz, I.; Redard-Jacot, M.; Schijman, A.G.; Torrico, F.; Sosa-Estani, S.; Katz, Z.; Ndung’u, J.M.; Ndung’u, J.M. The Burden of Congenital Chagas Disease and Implementation of Molecular Diagnostic Tools in Latin America. BMJ Glob. Health 2018, 3, e001069. [Google Scholar] [CrossRef] [Green Version]
- Carlier, Y.; Altcheh, J.; Angheben, A.; Freilij, H.; Luquetti, A.O.; Schijman, A.G.; Segovia, M.; Wagner, N.; Vinas, P.A. Congenital Chagas Disease: Updated Recommendations for Prevention, Diagnosis, Treatment, and Follow-up of Newborns and Siblings, Girls, Women of Childbearing Age, and Pregnant Women. PLoS Negl. Trop. Dis. 2019, 13, e0007694. [Google Scholar] [CrossRef]
- Basile, L.; Jansà, J.M.; Carlier, Y.; Salamanca, D.D.; Angheben, A.; Bartoloni, A.; Seixas, J.; van Gool, T.; Cañavate, C.; Flores-Chávez, M.; et al. Chagas Disease in European Countries: The Challenge of a Surveillance System. Eurosurveillance 2011, 16, 3. [Google Scholar] [CrossRef] [Green Version]
- Liempi, A.; Castillo, C.; Carrillo, I.; Muñoz, L.; Droguett, D.; Galanti, N.; Maya, J.D.J.D.; Kemmerling, U. A Local Innate Immune Response against Trypanosoma cruzi in the Human Placenta: The Epithelial Turnover of the Trophoblast. Microb. Pathog. 2016, 99, 123–129. [Google Scholar] [CrossRef]
- Liempi, A.; Castillo, C.; Duaso, J.; Droguett, D.; Sandoval, A.; Barahona, K.; Hernández, A.; Galanti, N.; Maya, J.D.J.D.; Kemmerling, U. Trypanosoma cruzi Induces Trophoblast Differentiation: A Potential Local Antiparasitic Mechanism of the Human Placenta? Placenta 2014, 35, 1035–1042. [Google Scholar] [CrossRef]
- Mayhew, T.M. Turnover of Human Villous Trophoblast in Normal Pregnancy: What Do We Know and What Do We Need to Know? Placenta 2014, 35, 229–240. [Google Scholar] [CrossRef]
- Huppertz, B.; Gauster, M. Trophoblast Fusion. Adv. Exp. Med. Biol. 2011, 713, 81–95. [Google Scholar] [CrossRef]
- Duval, M.; Cossart, P.; Lebreton, A. Mammalian MicroRNAs and Long Noncoding RNAs in the Host-Bacterial Pathogen Crosstalk. Semin. Cell Dev. Biol. 2017, 65, 11–19. [Google Scholar] [CrossRef]
- Hayder, H.; O’Brien, J.; Nadeem, U.; Peng, C. MicroRNAs: Crucial Regulators of Placental Development. Reproduction 2018, 155, R259–R271. [Google Scholar] [CrossRef] [Green Version]
- Saliminejad, K.; Khorram Khorshid, H.; Soleymani Fard, S.; Ghaffari, S. An Overview of MicroRNAs: Biology, Functions, Therapeutics, and Analysis Methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Medina, L.; Castillo, C.; Liempi, A.; Guerrero-Muñoz, J.; Rojas-Pirela, M.; Maya, J.D.; Prieto, H.; Kemmerling, U. Trypanosoma cruzi and Toxoplasma Gondii Induce a Differential MicroRNA Profile in Human Placental Explants. Front. Immunol. 2020, 11, 595250. [Google Scholar] [CrossRef]
- Sadovsky, Y.; Mouillet, J.F.; Ouyang, Y.; Bayer, A.; Coyne, C.B. The Function of Trophomirs and Other Micrornas in the Human Placenta. Cold Spring Harb. Perspect. Med. 2015, 5, a023036. [Google Scholar] [CrossRef] [Green Version]
- Donker, R.B.; Mouillet, J.F.; Chu, T.; Hubel, C.A.; Stolz, D.B.; Morelli, A.E.; Sadovsky, Y. The Expression Profile of C19MC MicroRNAs in Primary Human Trophoblast Cells and Exosomes. Mol. Hum. Reprod. 2012, 18, 417–424. [Google Scholar] [CrossRef] [Green Version]
- Arora, N.; Sadovsky, Y.; Dermody, T.S.; Coyne, C.B. Microbial Vertical Transmission during Human Pregnancy. Cell Host Microbe 2017, 21, 561–567. [Google Scholar] [CrossRef]
- Chu, T.; Mouillet, J.F.; Cao, Z.; Barak, O.; Ouyang, Y.; Sadovsky, Y. RNA Network Interactions During Differentiation of Human Trophoblasts. Front. Cell Dev. Biol. 2021, 9, 677981. [Google Scholar] [CrossRef]
- Chen, F.; Zhu, H.-H.; Zhou, L.-F.; Wu, S.-S.; Wang, J.; Chen, Z. Inhibition of C-FLIP Expression by MiR-512-3p Contributes to Taxol-Induced Apoptosis in Hepatocellular Carcinoma Cells. Oncol. Rep. 2010, 23, 1457–1462. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, I.; Droguett, D.; Castillo, C.; Liempi, A.; Muñoz, L.; Maya, J.D.; Galanti, N.; Kemmerling, U. Caspase-8 Activity Is Part of the BeWo Trophoblast Cell Defense Mechanisms against Trypanosoma cruzi Infection. Exp. Parasitol. 2016, 168, 9–15. [Google Scholar] [CrossRef]
- Zhang, M.; Muralimanoharan, S.; Wortman, A.C.; Mendelson, C.R. Primate-Specific MiR-515 Family Members Inhibit Key Genes in Human Trophoblast Differentiation and Are Upregulated in Preeclampsia. Proc. Natl. Acad. Sci. USA 2016, 113, E7069–E7076. [Google Scholar] [CrossRef] [Green Version]
- Benirschke, K.; Kaufmann, P.; Baergen, P. Pathology of the Human Placenta, 6th ed.; SpringerLink: New York, NY, USA, 2012. [Google Scholar]
- Zheng, Y.; Cai, X.; Bradley, J.E. MicroRNAs in Parasites and Parasite Infection. RNA Biol. 2013, 10, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Acuña, S.M.; Floeter-Winter, L.M.; Muxel, S.M. MicroRNAs: Biological Regulators in Pathogen–Host Interactions. Cells 2020, 9, 113. [Google Scholar] [CrossRef] [Green Version]
- Villares, M.; Berthelet, J.; Weitzman, J.B. The Clever Strategies Used by Intracellular Parasites to Hijack Host Gene Expression. Semin. Immunopathol. 2020, 42, 215–226. [Google Scholar] [CrossRef]
- Castillo, C.; Carrillo, I.; Libisch, G.; Juiz, N.; Schijman, A.G.; Robello, C.; Kemmerling, U. Host-Parasite Interaction: Changes in Human Placental Gene Expression Induced by Trypanosoma cruzi. Parasites Vectors 2018, 11, 479. [Google Scholar] [CrossRef]
- Mouillet, J.F.; Ouyang, Y.; Coyne, C.B.; Sadovsky, Y. MicroRNAs in Placental Health and Disease. Am. J. Obstet. Gynecol. 2015, 213, S163–S172. [Google Scholar] [CrossRef] [Green Version]
- Hromadnikova, I.; Kotlabova, K.; Ondrackova, M.; Pirkova, P.; Kestlerova, A.; Novotna, V.; Hympanova, L.; Krofta, L. Expression Profile of C19MC MicroRNAs in Placental Tissue in Pregnancy-Related Complications. DNA Cell Biol. 2015, 34, 437–457. [Google Scholar] [CrossRef]
- Carlier, Y.; Schijman, A.G.; Kemmerling, U. Placenta, Trypanosoma cruzi, and Congenital Chagas Disease. Curr. Trop. Med. Rep. 2020, 7, 172–182. [Google Scholar] [CrossRef]
- Chang, S.; Lee, S.; Yang, L.; Kim, D.; Yoon, J.; Ko, H.; Ogawa, M.; Sasakawa, C.; Kweon, M. Autophagy Controls an Intrinsic Host Defense to Bacteria by Promoting Epithelial Cell Survival: A Murine Model. PLoS ONE 2013, 8, e81095. [Google Scholar] [CrossRef] [Green Version]
- Kasper, C.A.; Sorg, I.; Schmutz, C.; Tschon, T.; Wischnewski, H.; Kim, M.L.; Arrieumerlou, C. Cell-Cell Propagation of NF-ΚB Transcription Factor and MAP Kinase Activation Amplifies Innate Immunity against Bacterial Infection. Immunity 2010, 33, 804–816. [Google Scholar] [CrossRef] [Green Version]
- Liempi, A.; Castillo, C.; Medina, L.; Rojas, M.; Maya, J.D.J.D.; Parraguez, V.H.V.H.; Kemmerling, U. Ex Vivo Infection of Human Placental Explants with Trypanosoma cruzi and Toxoplasma Gondii: Differential Activation of NF Kappa B Signaling Pathways. Acta Trop. 2019, 199, 105153. [Google Scholar] [CrossRef]
- Castillo, C.; Villarroel, A.; Duaso, J.; Galanti, N.; Cabrera, G.; Maya, J.D.; Kemmerling, U. Phospholipase C Gamma and ERK1/2 Mitogen Activated Kinase Pathways Are Differentially Modulated by Trypanosoma cruzi during Tissue Invasion in Human Placenta. Exp. Parasitol. 2013, 133, 12–17. [Google Scholar] [CrossRef]
- Morales-Prieto, D.M.; Ospina-Prieto, S.; Chaiwangyen, W.; Schoenleben, M.; Markert, U.R. Pregnancy-Associated MiRNA-Clusters. J. Reprod. Immunol. 2013, 97, 51–61. [Google Scholar] [CrossRef]
- Yang, Q.; Lu, J.; Wang, S.; Li, H.; Ge, Q.; Lu, Z. Application of Next-Generation Sequencing Technology to Profile the Circulating MicroRNAs in the Serum of Preeclampsia versus Normal Pregnant Women. Clin. Chim. Acta 2011, 412, 2167–2173. [Google Scholar] [CrossRef]
- Kumar, P.; Luo, Y.; Tudela, C.; Alexander, J.M.; Mendelson, C.R. The C-Myc-Regulated MicroRNA-17 92 (MiR-17 92) and MiR-106a 363 Clusters Target HCYP19A1 and HGCM1 To Inhibit Human Trophoblast Differentiation. Mol. Cell. Biol. 2013, 33, 1782–1796. [Google Scholar] [CrossRef] [Green Version]
- Meinhardt, G.; Saleh, L.; Otti, G.R.; Haider, S.; Velicky, P.; Fiala, C.; Pollheimer, J.; Knöfler, M. Wingless Ligand 5a Is a Critical Regulator of Placental Growth and Survival. Sci. Rep. 2016, 6, 28127. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.W.; Yan, T.Q.; Tong, H. Effect of MiR-515-5p on Proliferation and Drug Sensitivity of Retinoblastoma Cells. Cancer Manag. Res. 2020, 12, 12087–12098. [Google Scholar] [CrossRef]
- Li, M.; Chen, W.D.; Wang, Y.D. The Roles of the Gut Microbiota-MiRNA Interaction in the Host Pathophysiology. Mol. Med. 2020, 26, 101. [Google Scholar] [CrossRef]
- Villalta, F.; Kierszenbaum, F. Growth of Isolated Amastigotes of Trypanosoma cruzi in Cell-Free Medium. J. Protozool. 1982, 29, 570–576. [Google Scholar] [CrossRef]
- Duaso, J.; Rojo, G.; Cabrera, G.; Galanti, N.; Bosco, C.; Maya, J.D.; Morello, A.; Kemmerling, U. Trypanosoma cruzi Induces Tissue Disorganization and Destruction of Chorionic Villi in an Ex Vivo Infection Model of Human Placenta. Placenta 2010, 31, 705–711. [Google Scholar] [CrossRef]
- Rebustini, I.T. A Functional MicroRNA Screening Method for Organ Morphogenesis. Curr. Protoc. Cell Biol. 2017, 74, 19.19.1–19.19.17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponce, I.; Aldunate, C.; Valenzuela, L.; Sepúlveda, S.; Garrido, G.; Kemmerling, U.; Cabrera, G.; Galanti, N. A Flap Endonuclease (TcFEN1) Is Involved in Trypanosoma cruzi Cell Proliferation, DNA Repair, and Parasite Survival. J. Cell. Biochem. 2017, 118, 1722–1732. [Google Scholar] [CrossRef] [PubMed]
- Liempi, A.; Castillo, C.; Medina, L.; Galanti, N.; Maya, J.D.; Parraguez, V.H.; Kemmerling, U. Comparative Ex Vivo Infection with Trypanosoma cruzi and Toxoplasma Gondii of Human, Canine and Ovine Placenta: Analysis of Tissue Damage and Infection Efficiency. Parasitol. Int. 2020, 76, 102065. [Google Scholar] [CrossRef]
- Androvic, P.; Valihrach, L.; Elling, J.; Sjoback, R.; Kubista, M. Two-Tailed RT-QPCR: A Novel Method for Highly Accurate MiRNA Quantification. Nucleic Acids Res. 2017, 45, e144. [Google Scholar] [CrossRef]
- Castillo, C.; López-Muñoz, R.A.; Duaso, J.; Galanti, N.; Jaña, F.; Ferreira, J.; Cabrera, G.; Maya, J.D.; Kemmerling, U. Role of Matrix Metalloproteinases 2 and 9 in Ex Vivo Trypanosoma cruzi Infection of Human Placental Chorionic Villi. Placenta 2012, 33, 991–997. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| qPCR Primers | Sequence |
|---|---|
| miR-512-3p | AAGUGCUGUCAUAGCUGAGGUC |
| miR-515-5p | UUCUCCAAAAGAAAGCACUUUCUG |
| RNU6-1 | GUGCUCGCUUCGGCAGCACAUAUACUAAAAUUGGAA CGAUACAGAGAAGAUUAGCAUGGCCCCUGCGCAAGG AUGACACGCAAAUUCGUGAAGCGUUCCAUAUUUU |
| qPCR Primers | Primer Forward | Primer Reverse | Product Length |
|---|---|---|---|
| c-FLIP | CAGGAACCCTCACCTTGTT | CGGCCCATGTAATCCTTCAT | 114 |
| hGCM1 | AAGCCCTAGAAAACAATCTC | AGGTTCCATGATAAGGTCAG | 148 |
| hCG | CCCCTTGACCTGTGATGACC | TATTGTGGGAGGATCGGGGT | 120 |
| hGAPDH | AACAGCGACACCCACTCCTC | GGAGGGGAGATTCAGTGTGGT | 258 |
| qPCR Primers | Primer Forward | Primer Reverse | Product Length |
|---|---|---|---|
| T. cruzi | GCTCTTGCCCACAMGGGTGC | CAAGCAGCGGATAGTTCAGG | 181 |
| hGAPDH | TGATGCGTGTACAAGCGTTTT | ACATGGTATTCACCACCCCACTAT | 97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medina, L.; Guerrero-Muñoz, J.A.; Liempi, A.I.; Castillo, C.; Ortega, Y.; Sepúlveda, A.; Salomó, F.; Maya, J.D.; Kemmerling, U. Ex Vivo Infection of Human Placental Explants by Trypanosoma cruzi Reveals a microRNA Profile Similar to That Seen in Trophoblast Differentiation. Pathogens 2022, 11, 361. https://doi.org/10.3390/pathogens11030361
Medina L, Guerrero-Muñoz JA, Liempi AI, Castillo C, Ortega Y, Sepúlveda A, Salomó F, Maya JD, Kemmerling U. Ex Vivo Infection of Human Placental Explants by Trypanosoma cruzi Reveals a microRNA Profile Similar to That Seen in Trophoblast Differentiation. Pathogens. 2022; 11(3):361. https://doi.org/10.3390/pathogens11030361
Chicago/Turabian StyleMedina, Lisvaneth, Jesús Alejandro Guerrero-Muñoz, Ana Isabel Liempi, Christian Castillo, Yessica Ortega, Alfredo Sepúlveda, Fernando Salomó, Juan Diego Maya, and Ulrike Kemmerling. 2022. "Ex Vivo Infection of Human Placental Explants by Trypanosoma cruzi Reveals a microRNA Profile Similar to That Seen in Trophoblast Differentiation" Pathogens 11, no. 3: 361. https://doi.org/10.3390/pathogens11030361