
Pseudomonas syringae on Plants in Iceland Has Likely Evolved for Several Million Years Outside the Reach of Processes That Mix This Bacterial Complex across Earth’s Temperate Zones

 , , and
, , and
Abstract
:1. Introduction
2. Results
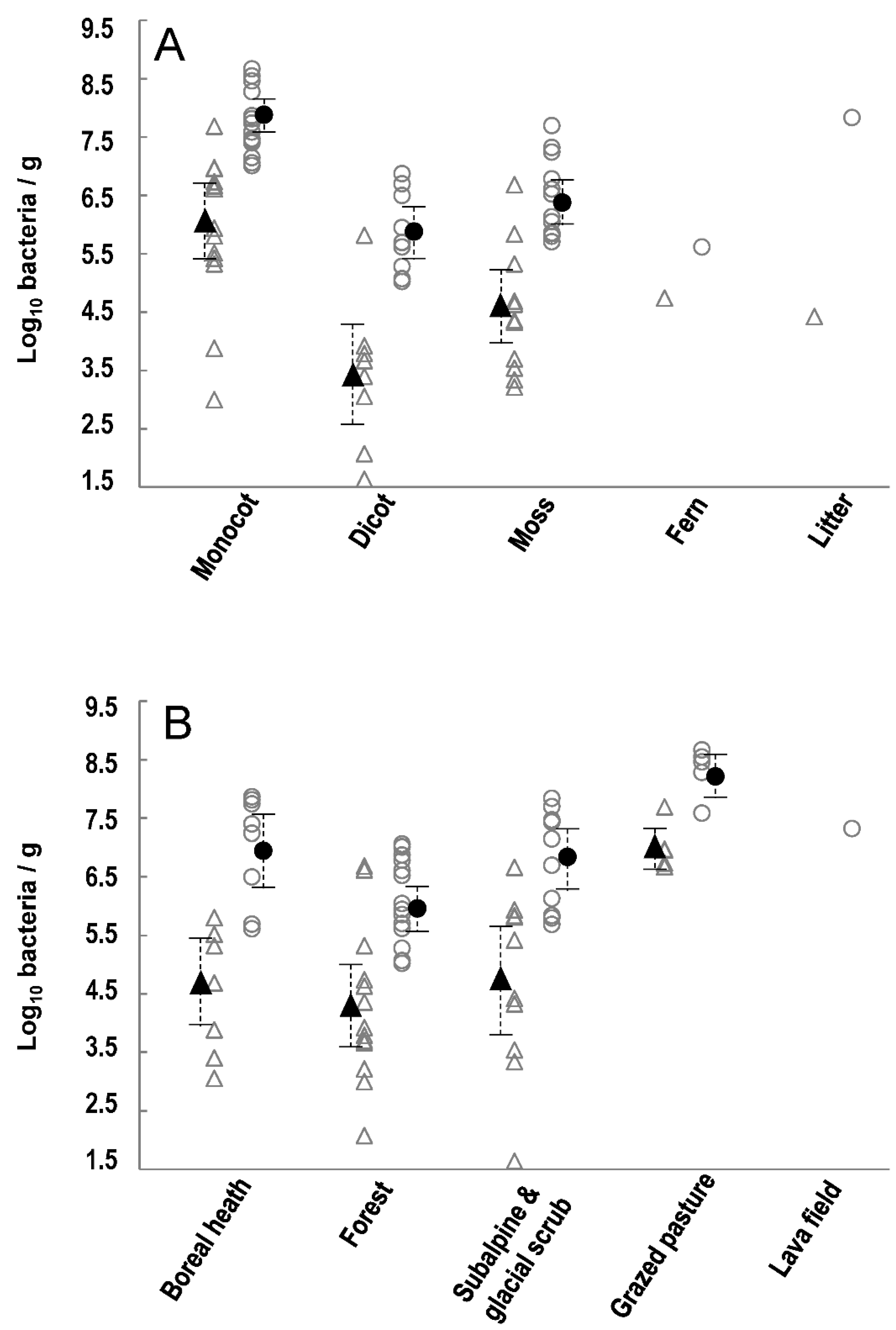
2.1. P. syringae Is Ubiquitous and Abundant on Vegetation across Iceland
2.2. P. syringae in Iceland Is Genetically Diverse but Distinct from P. syringae Elsewhere
2.3. The Size, Composition and Co-Occurrence of Certain Phylogroups of P. syringae Populations on Vegetation Depend on Habitat and Plant Type
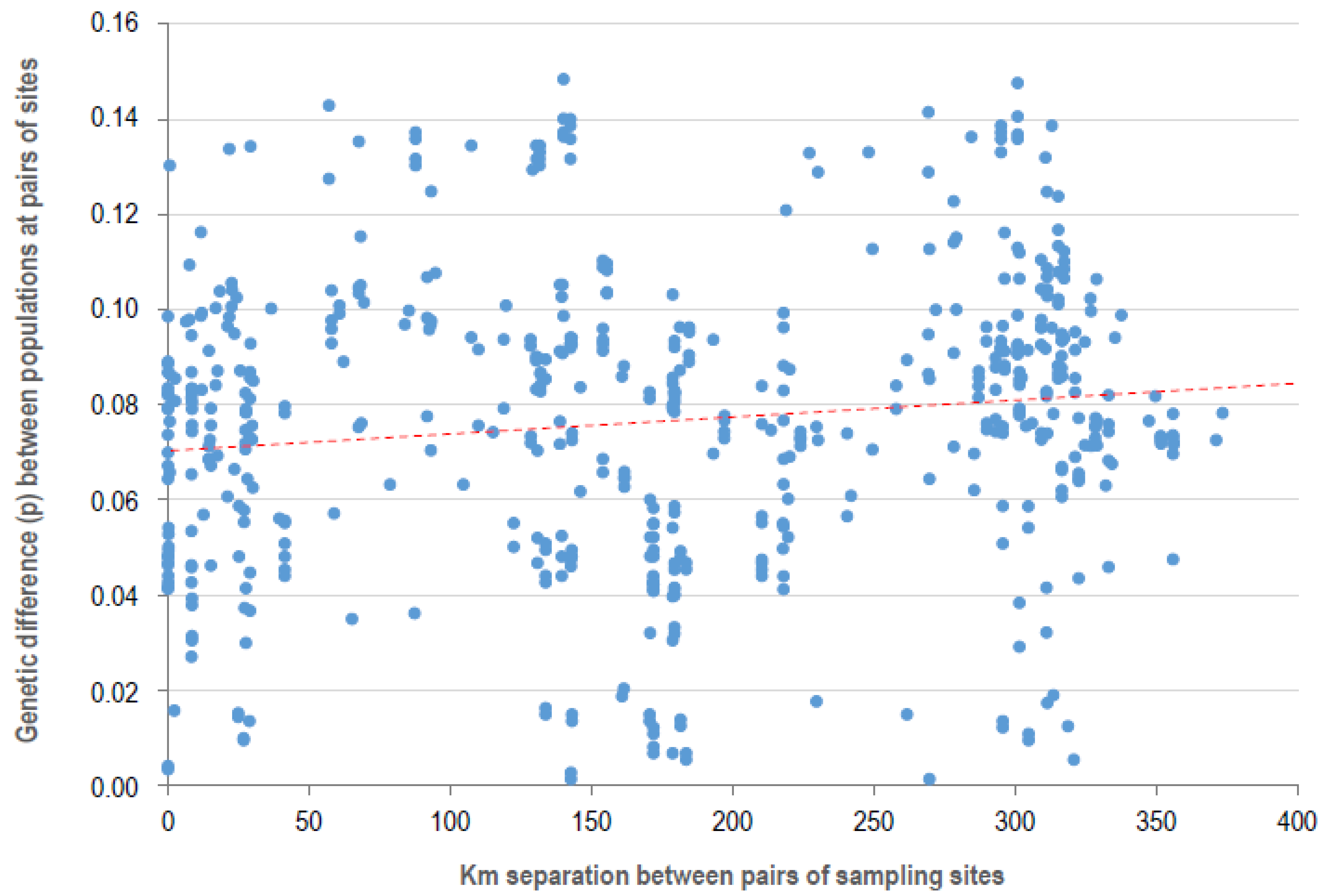
2.4. Haplotypes of P. syringae on Icelandic Vegetation Diverged near the Time of the First Fossil Evidence of Plants on Iceland as Well as Very Recently
3. Discussion
4. Materials and Methods
4.1. Samples and Sampling Sites

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Date | Latitude °N | Longitude °W | Habitat Type | Category of Plant Sample | Identity of Sampled Material | log10 cfu P. syringae/g Tissue | log10 cfu Total Bacteria/g Tissue |
|---|---|---|---|---|---|---|---|---|
| BR2003 | Augustust 2020 | 65.239038 | −20.851623 | Forest | monocot | mixed grasses | 6.62 | 7.01 |
| EG2002 | September 2020 | 65.262654 | −14.378452 | Forest | moss | Hylocomium splendens | 3.70 | 6.05 |
| EG2003 | August 2020 | 65.262654 | −14.378453 | Forest | monocot | Festuca sp. | 2.99 | 7.06 |
| EG2005 | September 2020 | 65.267478 | −14.331496 | Forest | dicot | Empetrum nigrum | 2.07 | 5.07 |
| EG2007 | September 2020 | 65.096122 | −14.732982 | Forest | moss | Hylocomium splendens | 3.21 | 5.86 |
| EG2008 | September 2020 | 65.096122 | −14.732983 | Forest | fern | Equisetum pratense | 4.74 | 5.62 |
| EG2011 | September 2020 | 65.094289 | −14.734456 | Forest | dicot | Rubus saxatilis | 3.79 | 5.28 |
| EG2012 | September 2020 | 65.094289 | −14.734457 | Forest | moss | Rhytidiadelphus loreus | 5.32 | 6.61 |
| EG2015 | August 2020 | 65.036883 | −14.620112 | Forest | dicot | Salix arctica | 3.67 | 5.95 |
| EG2016 | September 2020 | 65.036883 | −14.620113 | Forest | moss | Rhytidiadelphus squarrosus | 4.36 | 6.78 |
| EG2018 | September 2020 | 65.036937 | −14.620176 | Forest | dicot | Vaccinium uliginosum | 3.92 | 5.03 |
| EG2019 | September 2020 | 65.036937 | −14.620177 | Forest | moss | Rhytidiadelphus loreus | 4.63 | 6.52 |
| EG2021 | September 2020 | 65.036937 | −14.620176 | Forest | dicot | Alchemilla alpina | nd1 | 6.87 |
| EG2022 | September 2020 | 65.036937 | −14.620177 | Forest | moss | Hylocomium splendens | 6.68 | 5.70 |
| HV2005 | August 2020 | 65.402968 | −20.896696 | Boreal heath | dicot | Alchemilla alpina | 3.06 | 6.50 |
| HV2006 | August 2020 | 65.400877 | −20.884433 | Boreal heath | dicot | Alchemilla alpina | nd | 5.62 |
| HV2009 | August 2020 | 65.411194 | −21.215754 | Boreal heath | dicot | Salix herbacea | 3.40 | 5.69 |
| IS1801 | June 2018 | 65.904903 | −18.838611 | Subalpine & glacial scrub | monocot | sparse mixed grass | 5.94 | 7.43 |
| IS1802 | June 2018 | 65.925386 | −18.823611 | Subalpine & glacial scrub | monocot | sparse mixed grass | 6.66 | 7.47 |
| IS1803 | June 2018 | 66.024216 | −16.493893 | Subalpine & glacial scrub | litter | litter on forest floor | 4.42 | 7.84 |
| IS1804 | June 2018 | 66.5145298 | −16.132639 | Subalpine & glacial scrub | monocot | mixed grasses and mosses | 5.42 | 7.15 |
| IS1805 | June 2018 | 66.514528 | −16.132639 | Subalpine & glacial scrub | moss | mixed grasses and mosses | 4.33 | 6.13 |
| IS1806 | June 2018 | 65.163587 | −21.032756 | Boreal heath | monocot | mixed grasses and mosses | 5.52 | 7.81 |
| IS1807 | June 2018 | 65.163587 | −21.032756 | Boreal heath | moss | mixed grasses and mosses | 3.88 | 7.40 |
| IS1808 | June 2018 | 65.162114 | −21.019500 | Boreal heath | monocot | mixed grasses and mosses | 5.33 | 7.86 |
| IS1809 | June 2018 | 65.162114 | −21.019500 | Subalpine & glacial scrub | moss | mixed grasses and mosses | 5.84 | 7.69 |
| IS1810 | June 2018 | 65.205417 | −21.326132 | Boreal heath | monocot | mixed grasses and mosses | 5.80 | 7.74 |
| IS1811 | June 2018 | 65.205417 | −21.326132 | Boreal heath | moss | mixed grasses and mosses | 4.68 | 7.24 |
| IS1812 | June 2018 | 65.218333 | −21.370000 | Grazed pasture | monocot | farm grass | 6.97 | 8.54 |
| IS1813 | June 2018 | 65.184495 | −21.480230 | Grazed pasture | monocot | farm grass | 6.74 | 8.28 |
| IS1814 | June 2018 | 64.777373 | −21.520000 | Lava field | moss | moss on lava rock | nd | 7.32 |
| IS1901 | September 2019 | 64.660454 | −21.338979 | Grazed pasture | monocot | pasture grass and clover | 7.69 | 8.67 |
| IS1902 | September 2019 | 64.601361 | −21.563346 | Grazed pasture | monocot | pasture grass | 6.95 | 8.46 |
| IS1903 | September 2019 | 64.428018 | −21.959702 | Grazed pasture | monocot | pasture grass | 6.67 | 7.58 |
| SU2002 | August 2020 | 65.649893 | −18.183058 | Subalpine & glacial scrub | dicot | Empetrum nigrum | 5.82 | 6.70 |
| SU2003 | August 2020 | 65.649893 | −18.183058 | Subalpine & glacial scrub | moss | Hylocomium splendens | 3.54 | 5.84 |
| SU2005 | August 2020 | 65.650320 | −18.182976 | Subalpine & glacial scrub | dicot | Empetrum nigrum | 1.63 | 5.69 |
| SU2006 | August 2020 | 65.650320 | −18.182977 | Subalpine & glacial scrub | moss | Hylocomium splendens | 3.34 | 5.80 |
4.2. Isolation, Characterization and Quantification of Bacteria
4.3. Evaluation of Genetic Diversity and Divergence
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morris, C.E.; Monteil, C.L.; Berge, O. The life history of Pseudomonas syringae: Linking agriculture to Earth system processes. Annu. Rev. Phytopathol. 2013, 51, 85–104. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.E.; Sands, D.C.; Vanneste, J.L.; Montarry, J.; Oakley, B.; Guilbaud, C.; Glaux, C. Inferring the evolutionary history of the plant pathogen Pseudomonas syringae from its biogeography in headwaters of rivers in North America, Europe and New Zealand. mBio 2010, 1, e00107-10–e00107-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteil, C.L.; Yahara, K.; Studholme, D.J.; Mageiros, L.; Méric, G.; Swingle, B.; Morris, C.E.; Vinatzer, B.A.; Sheppard, S.K. Population genomic insights into the emergence, crop-adaptation, and dissemination of Pseudomonas syringae pathogens. Microb. Genom. 2016. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.E.; Conen, F.; Huffman, J.A.; Phillips, V.; Poeschl, U.; Sands, D.C. Bioprecipitation: A feedback cycle linking Earth history, ecosystem dynamics and land use through biological ice nucleators in the atmosphere. Glob. Chang. Biol. 2014, 20, 341–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berge, O.; Monteil, C.L.; Bartoli, C.; Chandeysson, C.; Guilbaud, C.; Sands, D.C.; Morris, C.E. A user’s guide to a data base of the diversity of Pseudomonas syringae and its application to classifying strains in this phylogenetic complex. PLoS ONE 2014, 9, e105547. [Google Scholar] [CrossRef] [PubMed]
- Denk, T.; Grímsson, F.; Zetter, R.; Símonarson, L.A. Introduction to the Nature and Geology of Iceland. In Late Cainozoic Floras of Iceland: 15 Million Years of Vegetation and Climate History in the Northern North Atlantic; Springer: Dordrecht, The Netherlands, 2011; pp. 1–29. [Google Scholar]
- Denk, T.; Grímsson, F.; Zetter, R.; Símonarson, L.A. The Archaic Floras. In Late Cainozoic Floras of Iceland: 15 Million Years of Vegetation and Climate History in the Northern North Atlantic; Springer: Dordrecht, The Netherlands, 2011; pp. 173–231. [Google Scholar]
- Foulger, G.R.; Doré, T.; Emeleus, C.H.; Franke, D.; Geoffroy, L.; Gernigon, L.; Hey, R.; Holdsworth, R.E.; Hole, M.; Höskuldsson, Á.; et al. The Iceland Microcontinent and a continental Greenland-Iceland-Faroe Ridge. Earth-Sci. Rev. 2020, 206, 102926. [Google Scholar] [CrossRef]
- Ehlers, J.; Gibbard, P.L. The extent and chronology of Cenozoic Global Glaciation. Quat. Int. 2007, 164–165, 6–20. [Google Scholar] [CrossRef]
- Buckland, P.C.; Perry, D.W.; Gklason, G.M.; Dugmore, A.J. The pre-Landnám fauna of Iceland: A palaeontological contribution. Boreas 1986, 15, 173–184. [Google Scholar] [CrossRef]
- Stewart, H.; Bradwell, T.; Bullard, J.; Davies, S.J.; Golledge, N.; McCulloch, R.D. 8000 years of North Atlantic storminess reconstructed from a Scottish peat record: Implications for Holocene atmospheric circulation patterns in Western Europe. J. Quat. Sci. 2017, 32, 1075–1084. [Google Scholar] [CrossRef]
- Brooks, D.R.; Hoberg, E.P.; Boeger, W.A. The Stockholm Paradigm: Climate Change and Emerging Disease; The University of Chicago Press: Chicago, IL, USA, 2019; p. 409. [Google Scholar]
- Brooks, D.R.; Hoberg, E.P.; Boeger, W.A.; Gardner, S.L.; Galbreath, K.E.; Herczeg, D.; Mejía-Madrid, H.H.; Rácz, S.E.; Dursahinhan, A.T. Finding them before they find us: Informatics, parasites, and environments in accelerating climate change. Comp. Parasitol. 2014, 81, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.E.; Bardin, M.; Kinkel, L.L.; Moury, B.; Nicot, P.C.; Sands, D.C. Expanding the paradigms of plant pathogen life history and evolution of parasitic fitness beyond agricultural boundaries. PLoS Pathog. 2009, 5, e1000693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, C.E.; Géniaux, G.; Nédellec, C.; Sauvion, N.; Soubeyrand, S. One Health concepts and challenges for surveillance, forecasting, and mitigation of plant disease beyond the traditional scope of crop production. Plant Pathol. 2022, 71, 86–97. [Google Scholar] [CrossRef]
- Prins, H.H.T. Why are mosses eaten in cold environments only? Oikos 1981, 38, 374–380. [Google Scholar] [CrossRef]
- Gornall, J.L.; Woodin, S.J.; Jónsdóttir, I.S.; Van der Wal, R. Herbivore impacts to the moss layer determine tundra ecosystem response to grazing and warming. Oecologia 2009, 161, 747–758. [Google Scholar] [CrossRef]
- Kornobis, E.; Pállsson, S.; Kristjánsson, B.K.; Svarsson, J. Molecular evidence of the survival of subterranean amphipods (Arthropoda) during Ice Age underneath glaciers in Iceland. Mol. Ecol. 2010, 19, 2516–2530. [Google Scholar] [CrossRef]
- Gaidos, E.; Lanoil, B.; Thorsteinsson, T.; Graham, A.; Skidmore, M.; Han, S.K.; Rust, T.; Popp, B. A viable microbial community in a subglacial volcanic crater lake, Iceland. Astrobiology 2004, 4, 327–344. [Google Scholar] [CrossRef]
- Clark, P.U.; Dyke, A.S.; Shakun, J.D.; Carlson, A.E.; Clark, J.; Wohlfarth, B.; Mitrovica, J.X.; Hostetler, S.W.; McCabe, A.M. The Last Glacial Maximum. Science 2009, 325, 710–714. [Google Scholar] [CrossRef] [Green Version]
- Christner, B.C.; Mosley-Thompson, E.; Thompson, L.G.; Zagorodnov, V.; Sandman, K.; Reeve, J.N. Recovery and identification of viable bacteria immured in glacial ice. Icarus 2000, 144, 479–485. [Google Scholar] [CrossRef]
- Karl, D.M.; Bird, D.F.; Björkman, K.; Houlihan, T.; Shackelford, R.; Tupas, L. Microorganisms in the accreted ice of Lake Vostok, Antarctica. Science 1999, 286, 2144–2147. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, A.; Kamel, M. Future threat from the past. Environ. Sci. Pollut. Res. 2021, 28, 1287–1291. [Google Scholar] [CrossRef]
- Monteil, C.L.; Guilbaud, C.; Glaux, C.; Lafolie, F.; Soubeyrand, S.; Morris, C.E. Emigration of the plant pathogen Pseudomonas syringae from leaf litter contributes to its population dynamics in alpine snowpack. Environ. Microbiol. 2012, 14, 2099–2112. [Google Scholar] [CrossRef] [PubMed]
- Monteil, C.L.; Lafolie, F.; Laurent, J.; Clement, J.-C.; Simler, R.; Travi, Y.; Morris, C.E. Soil water flow is a source of the plant pathogen Pseudomonas syringae in subalpine headwaters. Environ. Microbiol. 2013, 16, 2038–2052. [Google Scholar] [CrossRef] [PubMed]
- Bardin, M.; Leyronas, C.; Troulet, C.; Morris, C.E. Striking similarities between Botrytis cinerea from non-agricultural and from agricultural habitats. Front. Plant Sci. 2018, 9, 1820. [Google Scholar] [CrossRef] [PubMed]
- Monteil, C.L.; Bardin, M.; Morris, C.E. Features of air masses associated with the deposition of Pseudomonas syringae and Botrytis cinerea by rain and snowfall. ISME J. 2014, 8, 2290–2304. [Google Scholar] [CrossRef] [Green Version]
- Mohan, S.K.; Schaad, N.W. An improved agar plating assay for detecting Pseudomonas syringae pv. syringae and P. s. pv. phaseolicola in contaminated bean seed. Phytopathology 1987, 77, 1390–1395. [Google Scholar] [CrossRef]
- Morris, C.E.; Sands, D.C.; Vinatzer, B.A.; Glaux, C.; Guilbaud, C.; Buffière, A.; Yan, S.; Dominguez, H.; Thompson, B.M. The life history of the plant pathogen Pseudomonas syringae is linked to the water cycle. ISME J. 2008, 2, 321–334. [Google Scholar] [CrossRef] [Green Version]
- Mello, B. Estimating TimeTrees with MEGA and the TimeTree resource. Mol. Biol. Evol. 2018, 35, 2334–2342. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, H.E.; Thakur, S.; Gong, Y.; Fung, P.; Zhang, J.; Yuan, L.; Wang, P.W.; Yong, C.; Scortichini, M.; Guttman, D.S. Extensive remodeling of the Pseudomonas syringae pv. avellanae type III secretome associated with two independent host shifts onto hazelnut. BMC Microbiol. 2012, 12, 141. [Google Scholar] [CrossRef] [Green Version]
- Karasov, T.L.; Almario, J.; Friedemann, C.; Ding, W.; Giolai, M.; Heavens, D.; Kersten, S.; Lundberg, D.S.; Neumann, M.; Regalado, J.; et al. Arabidopsis thaliana and Pseudomonas pathogens exhibit stable associations over evolutionary timescales. Cell Host Microbe 2018, 24, 168–179.e164. [Google Scholar] [CrossRef] [Green Version]
- Chapman, J.R.; Taylor, R.K.; Weir, B.S.; Romberg, M.K.; Vanneste, J.L.; Luck, J.; Alexander, B.J.R. Phylogenetic relationships among global populations of Pseudomonas syringae pv. actinidiae. Phytopathology 2012, 102, 1034–1044. [Google Scholar] [CrossRef] [Green Version]
- Joly, M.; Attard, E.; Sancelme, M.; Deguillaume, L.; Guilbaud, C.; Morris, C.E.; Amato, P.; Delort, A.M. Ice nucleation activity of bacteria isolated from cloud water. Atmos. Environ. 2013, 70, 392–400. [Google Scholar] [CrossRef]
- Ruinelli, M.; Blom, J.; Smits, T.H.M.; Pothier, J.F. Comparative genomics and pathogenicity potential of members of the Pseudomonas syringae species complex on Prunus spp. BMC Genom. 2019, 20, 172. [Google Scholar] [CrossRef] [PubMed]
- Stopelli, E.; Conen, F.; Guilbaud, C.; Zopfi, J.; Alewell, C.; Morris, C.E. Ice nucleators, bacterial cells and Pseudomonas syringae in precipitation at Jungfraujoch. Biogeosciences 2017, 14, 1189–1196. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morris, C.E.; Ramirez, N.; Berge, O.; Lacroix, C.; Monteil, C.; Chandeysson, C.; Guilbaud, C.; Blischke, A.; Sigurbjörnsdóttir, M.A.; Vilhelmsson, O.Þ. Pseudomonas syringae on Plants in Iceland Has Likely Evolved for Several Million Years Outside the Reach of Processes That Mix This Bacterial Complex across Earth’s Temperate Zones. Pathogens 2022, 11, 357. https://doi.org/10.3390/pathogens11030357
Morris CE, Ramirez N, Berge O, Lacroix C, Monteil C, Chandeysson C, Guilbaud C, Blischke A, Sigurbjörnsdóttir MA, Vilhelmsson OÞ. Pseudomonas syringae on Plants in Iceland Has Likely Evolved for Several Million Years Outside the Reach of Processes That Mix This Bacterial Complex across Earth’s Temperate Zones. Pathogens. 2022; 11(3):357. https://doi.org/10.3390/pathogens11030357
Chicago/Turabian StyleMorris, Cindy E., Natalia Ramirez, Odile Berge, Christelle Lacroix, Cécile Monteil, Charlotte Chandeysson, Caroline Guilbaud, Anett Blischke, Margrét Auður Sigurbjörnsdóttir, and Oddur Þ. Vilhelmsson. 2022. "Pseudomonas syringae on Plants in Iceland Has Likely Evolved for Several Million Years Outside the Reach of Processes That Mix This Bacterial Complex across Earth’s Temperate Zones" Pathogens 11, no. 3: 357. https://doi.org/10.3390/pathogens11030357