
Therapeutic Efficacy of Orally Administered Nitrofurantoin against Animal African Trypanosomosis Caused by Trypanosoma congolense Infection

 ,
,  ,
,
Abstract
:1. Introduction
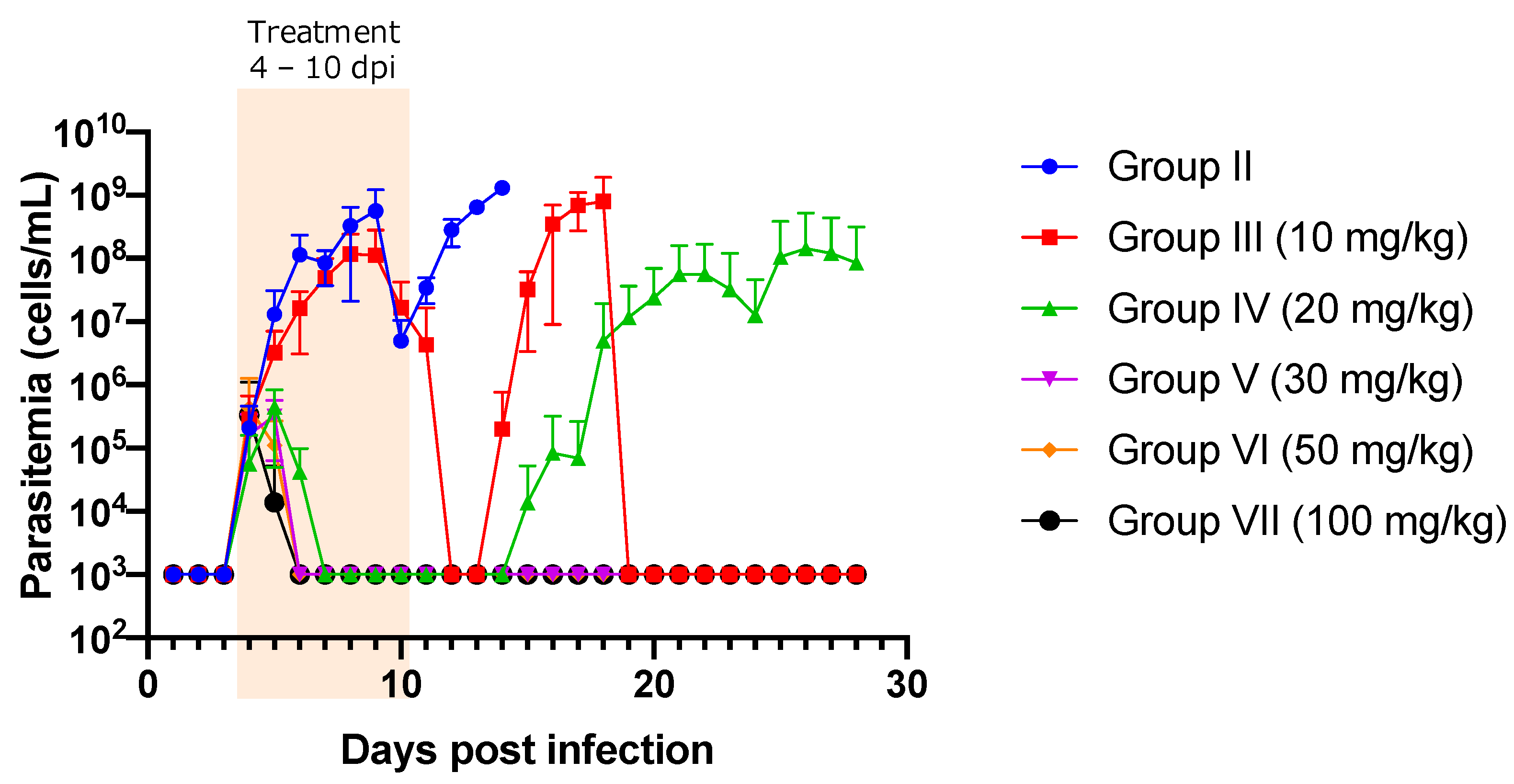
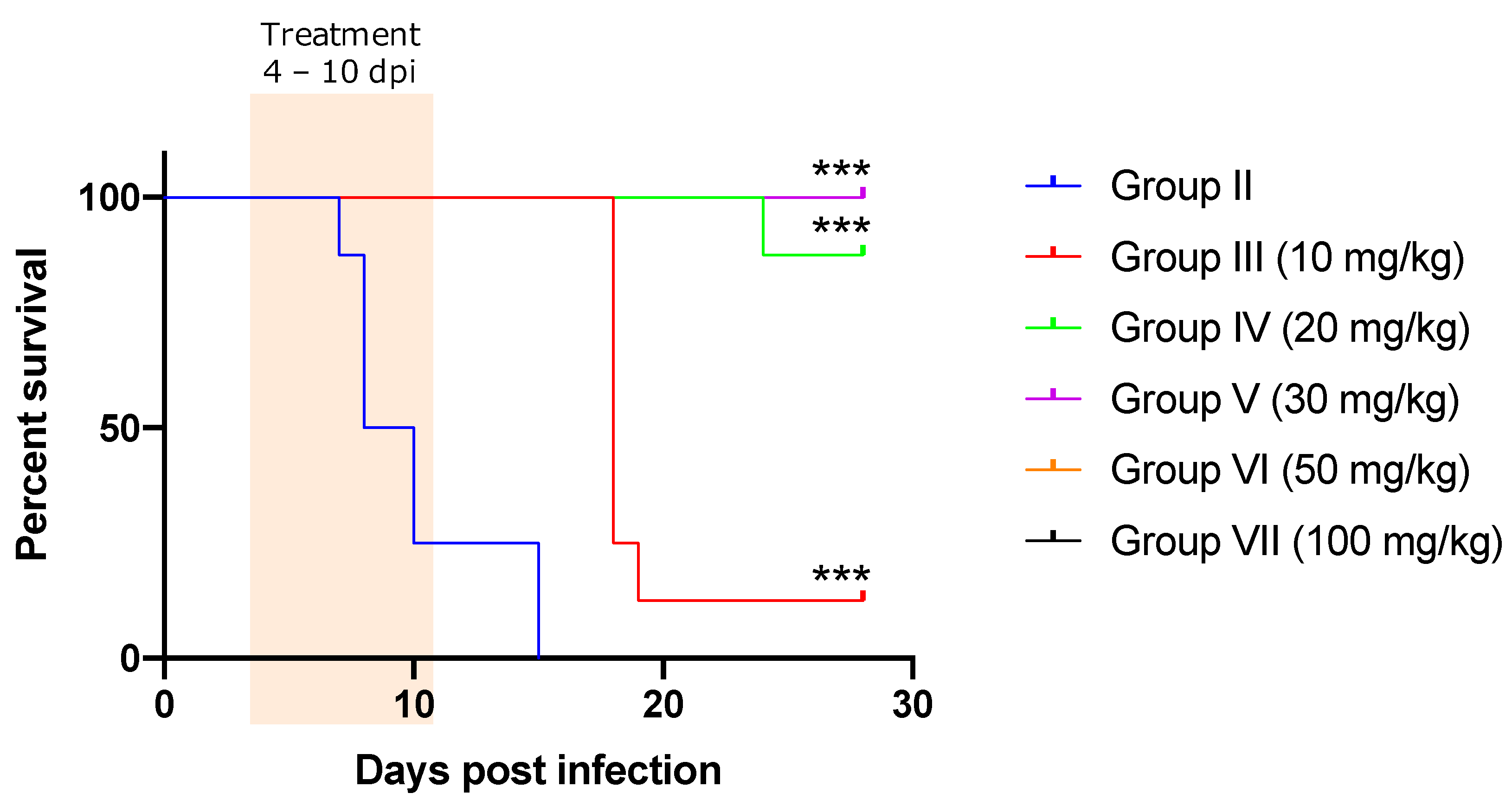
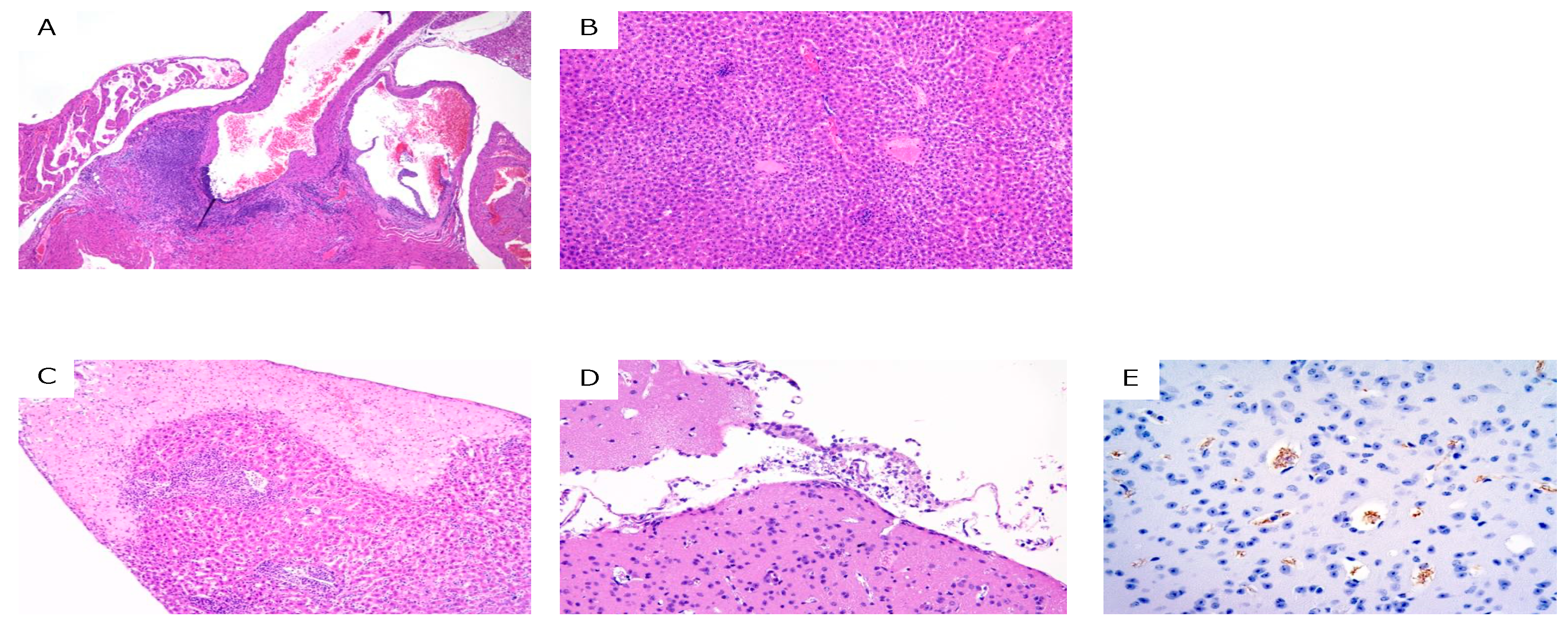
2. Results
Treatment Efficacy of NF against T. congolense-Infected Mice
3. Discussion
4. Materials and Methods
4.1. Animal Experiments
4.2. Evaluation of Parasitemia
4.3. Hematological Parameters
4.4. Histopathological Analyses
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giordani, F.; Morrison, L.J.; Rowan, T.G.; De Koning, H.P.; Barrett, M.P. The Animal Trypanosomiases and Their Chemotherapy: A Review. Parasitology 2016, 143, 1862–1889. [Google Scholar] [CrossRef] [PubMed]
- Morrison, L.J.; Vezza, L.; Rowan, T.; Hope, J.C. Animal African Trypanosomiasis: Time to Increase Focus on Clinically Relevant Parasite and Host Species. Trends Parasitol. 2016, 32, 599–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gizaw, Y.; Megersa, M.; Fayera, T. Dourine: A Neglected Disease of Equids. Trop. Anim. Health Prod. 2017, 49, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Aregawi, W.G.; Agga, G.E.; Abdi, R.D.; Büscher, P. Systematic Review and Meta-Analysis on the Global Distribution, Host Range, and Prevalence of Trypanosoma evansi. Parasit. Vectors 2019, 12, 67. [Google Scholar] [CrossRef] [Green Version]
- Desquesnes, M.; Holzmuller, P.; Lai, D.H.; Dargantes, A.; Lun, Z.R.; Jittaplapong, S. Trypanosoma evansi and Surra: A Review and Perspectives on Origin, History, Distribution, Taxonomy, Morphology, Hosts, and Pathogenic Effects. BioMed Res. Int. 2013, 2013, 194176. [Google Scholar] [CrossRef] [Green Version]
- Adams, E.R.; Hamilton, P.B.; Gibson, W.C. African Trypanosomes: Celebrating Diversity. Trends Parasitol. 2010, 26, 324–328. [Google Scholar] [CrossRef]
- Magez, S.; Caljon, G.; Tran, T.; Stijlemans, B.; Radwanska, M. Current Status of Vaccination Against African Trypanosomiasis. Parasitology 2010, 137, 2017–2027. [Google Scholar] [CrossRef] [Green Version]
- Autheman, D.; Crosnier, C.; Clare, S.; Goulding, D.; Brandt, C.; Harcourt, K.; Tolley, C.; Galaway, F.; Khushu, M.; Ong, H.; et al. An invariant Trypanosoma vivax vaccine antigen induces protective immunity. Nature 2021, 595, 96–100. [Google Scholar] [CrossRef]
- Schofield, C.J.; Kabayo, J.P. Trypanosomiasis Vector Control in Africa and Latin America. Parasit. Vectors 2008, 1, 24. [Google Scholar] [CrossRef] [Green Version]
- Courtin, F.; Kaba, D.; Rayaisse, J.-B.; Solano, P.; Torr, S.; Shaw, A. The cost of tsetse control using ‘Tiny Targets’ in the sleeping sickness endemic forest area of Bonon in Côte d’Ivoire: Implications for comparing costs across different settings. PLoS Negl. Trop. Dis. 2007, 16, e0010033. [Google Scholar] [CrossRef]
- Rayaisse, J.-B.; Courtin, F.; Mahamat, M.; Chérif, M.; Yoni, W.; Gadjibet, N.; Peka, M.; Solano, P.; Torr, S.; Shaw, A. Delivering ‘tiny targets’ in a remote region of southern Chad: A cost analysis of tsetse control in the Mandoul sleeping sickness focus. Parasit. Vectors 2020, 13, 419. [Google Scholar] [CrossRef] [PubMed]
- Muyobela, J.; Pirk, C.; Yusuf, A.; Mbewe, N.; Sole, C. A novel vehicle-mounted sticky trap; an effective sampling tool for savannah tsetse flies Glossina morsitans Westwood and Glossina morsitans centralis Machado. PLoS Negl. Trop. Dis. 2021, 15, e0009620. [Google Scholar] [CrossRef] [PubMed]
- Percoma, L.; Sow, A.; Pagabeleguen, S.; Dicko, A.; Serdebéogo, O.; Ouédraogo, M.; Rayaissé, J.-B.; Bouyer, J.; Belem, A.; Sidibé, I. Impact of an integrated control campaign on tsetse populations in Burkina Faso. Parasit. Vectors 2018, 11, 270. [Google Scholar] [CrossRef] [PubMed]
- Diall, O.; Cecchi, G.; Wanda, G.; Argilés-Herrero, R.; Vreysen, M.J.B.; Cattoli, G.; Viljoen, G.J.; Mattioli, R.; Bouyer, J. Developing a Progressive Control Pathway for African Animal Trypanosomosis. Trends Parasitol. 2017, 33, 499–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, A.; Workineh, S. Drug Resistance in African Animal Trypanosomes: A Review. Afr. J. Microbiol. Res. 2018, 12, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Mungube, E.O.; Vitouley, H.S.; Allegye-Cudjoe, E.; Diall, O.; Boucoum, Z.; Diarra, B.; Sanogo, Y.; Randolph, T.; Bauer, B.; Zessin, K.H.; et al. Detection of Multiple Drug-Resistant Trypanosoma congolense Populations in Village Cattle of South-East Mali. Parasit. Vectors 2012, 5, 155. [Google Scholar] [CrossRef] [Green Version]
- Jamal, S.; Sigauque, I.; Macuamule, C.; Neves, L.; Penzhorn, B.L.; Marcotty, T.; Van Den Bossche, P. The Susceptibility of Trypanosoma congolense Isolated in Zambézia Province, Mozambique, to Isometamidium Chloride, Diminazene Aceturate and Homidium Chloride. Onderstepoort J. Vet. Res. 2005, 72, 333–338. [Google Scholar] [CrossRef] [Green Version]
- Mamoudou, A.; Delespaux, V.; Chepnda, V.; Hachimou, Z.; Andrikaye, J.P.; Zoli, A.; Geerts, S. Assessment of the Occurrence of Trypanocidal Drug Resistance in Trypanosomes of Naturally Infected Cattle in the Adamaoua Region of Cameroon Using the Standard Mouse Test and Molecular Tools. Acta Trop. 2008, 106, 115–118. [Google Scholar] [CrossRef]
- Clausen, P.H.; Sidibe, I.; Kaboré, I.; Bauer, B. Development of Multiple Drug Resistance of Trypanosoma congolense in Zebu Cattle Under High Natural Tsetse Fly Challenge in the Pastoral Zone of Samorogouan, Burkina Faso. Acta Trop. 1992, 51, 229–236. [Google Scholar] [CrossRef]
- Sow, A.; Sidibé, I.; Bengaly, Z.; Marcotty, T.; Séré, M.; Diallo, A.; Vitouley, H.S.; Nebié, R.L.; Ouédraogo, M.; Akoda, G.K.; et al. Field Detection of Resistance to Isometamidium Chloride and Diminazene Aceturate in Trypanosoma vivax From the Region of the Boucle du Mouhoun in Burkina Faso. Vet. Parasitol. 2012, 187, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Mulugeta, W.; Wilkes, J.; Mulatu, W.; Majiwa, P.A.; Masake, R.; Peregrine, A.S. Long-Term Occurrence of Trypanosoma congolense Resistant to Diminazene, Isometamidium and Homidium in Cattle at Ghibe, Ethiopia. Acta Trop. 1997, 64, 205–217. [Google Scholar] [CrossRef]
- Ainanshe, O.A.; Jennings, F.W.; Holmes, P.H. Isolation of Drug-Resistant Strains of Trypanosoma congolense From the Lower Shabelle Region of Southern Somalia. Trop. Anim. Health Prod. 1992, 24, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Sinyangwe, L.; Delespaux, V.; Brandt, J.; Geerts, S.; Mubanga, J.; Machila, N.; Holmes, P.H.; Eisler, M.C. Trypanocidal Drug Resistance in Eastern Province of Zambia. Vet. Parasitol. 2004, 119, 125–135. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Interim Guidelines for the Treatment of Gambiense Human African Trypanosomiasis; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Mesu, V.K.B.K.; Kalonji, W.M.; Bardonneau, C.; Mordt, O.V.; Blesson, S.; Simon, F.; Delhomme, S.; Bernhard, S.; Kuziena, W.; Lubaki, J.-P.F.; et al. Oral Fexinidazole for Late-Stage African Trypanosoma brucei gambiense Trypanosomiasis: A Pivotal Multicentre, Randomised, Non-Inferiority Trial. Lancet 2018, 391, 144–154. [Google Scholar] [CrossRef]
- World Health Organization. World Health Organization Model List of Essential Medicines: 21st List 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Priotto, G.; Kasparian, S.; Ngouama, D.; Ghorashian, S.; Arnold, U.; Ghabri, S.; Karunakara, U. Nifurtimox-Eflornithine Combination Therapy for Second-Stage Trypanosoma Brucei Gambiense Sleeping Sickness: A Randomized Clinical Trial in Congo. Clin. Infect. Dis. 2007, 45, 1435–1442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Checchi, F.; Piola, P.; Ayikoru, H.; Thomas, F.; Legros, D.; Priotto, G. Nifurtimox Plus Eflornithine for Late-Stage Sleeping Sickness in Uganda: A Case Series. PLoS Negl. Trop. Dis. 2007, 1, e64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steverding, D. The Development of Drugs for Treatment of Sleeping Sickness: A Historical Review. Parasit. Vectors 2010, 3, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leuin, A.S.; Hartmann, F.; Viviano, K. Administration of nitrofurantoin in dogs with lower urinary tract infections: 14 cases (2013–2019). J. Small Anim. Pract. 2020, 62, 42–48. [Google Scholar] [CrossRef] [PubMed]
- MSD Manual Veterinaty Manual. Available online: https://www.msdvetmanual.com/pharmacology/antibacterial-agents/nitrofurans/ (accessed on 29 December 2021).
- Nitrofuran Ban in Effect. Available online: https://www.avma.org/javma-news/2002-09-15/nitrofuran-ban-effect (accessed on 29 December 2021).
- McCracken, R.J.; Kennedy, D.G. Detection, accumulation and distribution of nitrofuran residues in egg yolk, albumen and shell. Food Addit. Contam. 2007, 24, 26–33. [Google Scholar] [CrossRef]
- Munsimbwe, L.; Seetsi, A.; Namangala, B.; N’Da, D.D.; Inoue, N.; Suganuma, K. In vitro and in vivo Trypanocidal Efficacy of Synthesized Nitrofurantoin Analogs. Molecules 2021, 26, 3372. [Google Scholar] [CrossRef]
- Merino, G.; Perez, M.; Real, R.; Egido, E.; Prieto, J.G.; Alvarez, A.I. In vivo Inhibition of BCRP/ABCG2 Mediated Transport of Nitrofurantoin by the Isoflavones Genistein and Daidzein: A Comparative Study in Bcrp1 (−/−) Mice. Pharm. Res. 2010, 27, 2098–2105. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Morris, M.E. Effects of the Flavonoid Chrysin on Nitrofurantoin Pharmacokinetics in Rats: Potential Involvement of ABCG2. Drug Metab. Dispos. 2007, 35, 268–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijma, R.A.; Fransen, F.; Muller, A.E.; Mouton, J.W. Optimizing Dosing of Nitrofurantoin From a PK/PD Point of View: What Do We Need to Know? Drug Resist. Updat. 2019, 43, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wijma, R.A.; Huttner, A.; Koch, B.C.P.; Mouton, J.W.; Muller, A.E. Review of the Pharmacokinetic Properties of Nitrofurantoin and Nitroxoline. J. Antimicrob. Chemother. 2018, 73, 2916–2926. [Google Scholar] [CrossRef]
- Barour, D.; Berghiche, A.; Boulebda, N. Antimicrobial Resistance of Escherichia coli Isolates from Cattle in Eastern Algeria. Vet. World 2019, 12, 1195–1203. [Google Scholar] [CrossRef]
- Mubita, C.; Syakalima, M.; Chisenga, C.; Munyeme, M.; Bwalya, M.; Chifumpa, G.; Hang’ombe, B.M.; Sinkala, P.; Simuunza, M.; Fukushi, F.; et al. Antibiograms of Faecal Escherichia coli and Enterococci Species Isolated from Pastoralist Cattle in the Interface Areas of the Kafue Basin in Zambia. Vet. Arh. 2008, 78, 179–185. [Google Scholar]
- Giske, C.G. Contemporary Resistance Trends and Mechanisms for the Old Antibiotics Colistin, Temocillin, Fosfomycin, Mecillinam and Nitrofurantoin. Clin. Microbiol. Infect. 2015, 21, 899–905. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.S.; Lee, S.J.; Choe, H.S. Community-Acquired Urinary Tract Infection by Escherichia coli in the Era of Antibiotic Resistance. BioMed Res. Int. 2018, 2018, 7656752. [Google Scholar] [CrossRef] [Green Version]
- Zuma, N.H.; Aucamp, J.; N’Da, D.D. An Update on Derivatisation and Repurposing of Clinical Nitrofuran Drugs. Eur. J. Pharm. Sci. 2019, 140, 105092. [Google Scholar] [CrossRef]
- Erasmus, C.; Aucamp, J.; Smit, F.J.; Seldom, R.; Jordaan, A.; Warner, D.F.; N’Da, D.D. Synthesis and Comparison of in vitro Dual Anti-infective Activities of Novel Naphthoquinone Hybrids and Atovaquone. Biorg. Chem. 2021, 114, 10518. [Google Scholar] [CrossRef]
- Bachman, J.E.; Curtiss, L.A.; Assary, R.S. Investigation of the Redox Chemistry of Anthraquinone Derivatives Using Density Functional Theory. J. Phys. Chem. A 2014, 118, 8852–8860. [Google Scholar] [CrossRef] [PubMed]
- Moser, D.R.; Cook, G.A.; Ochs, D.E.; Bailey, C.P.; McKane, M.R.; Donelson, J.E. Detection of Trypanosoma congolense and Trypanosoma brucei subspecies by DNA Amplification Using the Polymerase Chain Reaction. Parasitology 1989, 99, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Sarwono, A.E.Y.; Suganuma, K.; Mitsuhashi, S.; Okada, T.; Musinguzi, S.P.; Shigetomi, K.; Inoue, N.; Ubukata, M. Identification and Characterization of Guanosine 5′-Monophosphate Reductase of Trypanosoma congolense as a Drug Target. Parasitol. Int. 2017, 66, 537–544. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Dose (mg/kg) | Total Mice Number | Survived Mice (%) | Euthanized Mice (%) | Mean Day of Survival in Euthanized Mice ± SD | Relapsed Mice (%) | Mean Day of Relapse ± SD | Re-infected Mice Numbers | Infected Mice in Re-Infection | % Infected in Re-Infected Mice |
|---|---|---|---|---|---|---|---|---|---|
| Group II (Non treated) | 8 | 0 (0.00) | 8 (100) | 9.15 ± 3.18 | 0 (0.00) | 14.85 ± 0.37 | 0 | NA | NA |
| Group III (10 mg/kg) | 8 | 1 (12.50) | 7 (87.50) | 17.14 ± 0.38 | 7 (87.5) | 14.85 ± 0.38 | 1 | 1 | 100 |
| Group IV (20 mg/kg) | 8 | 7 (87.50) | 1 (12.50) | 23.00 | 2 (25.00) | 17.00 ± 2.83 | 7 | 1 | 14.29 |
| Group V (30 mg/kg) | 8 | 8 (100) | 0 (0.00) | ND | 0 (0.00) | ND | 4 * | 0 | 0.00 |
| Group VI (50 mg/kg) | 8 | 8 (100) | 0 (0.00) | ND | 0 (0.00) | ND | 4 * | 0 | 0.00 |
| Group VII (100 mg/kg) | 8 | 8 (100) | 0 (0.00) | ND | 0 (0.00) | ND | 4 * | 0 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suganuma, K.; N’Da, D.D.; Watanabe, K.-i.; Tanaka, Y.; Mossaad, E.; Elata, A.; Inoue, N.; Kawazu, S.-i. Therapeutic Efficacy of Orally Administered Nitrofurantoin against Animal African Trypanosomosis Caused by Trypanosoma congolense Infection. Pathogens 2022, 11, 331. https://doi.org/10.3390/pathogens11030331
Suganuma K, N’Da DD, Watanabe K-i, Tanaka Y, Mossaad E, Elata A, Inoue N, Kawazu S-i. Therapeutic Efficacy of Orally Administered Nitrofurantoin against Animal African Trypanosomosis Caused by Trypanosoma congolense Infection. Pathogens. 2022; 11(3):331. https://doi.org/10.3390/pathogens11030331
Chicago/Turabian StyleSuganuma, Keisuke, David D. N’Da, Ken-ichi Watanabe, Yusuke Tanaka, Ehab Mossaad, Afraa Elata, Noboru Inoue, and Shin-ichiro Kawazu. 2022. "Therapeutic Efficacy of Orally Administered Nitrofurantoin against Animal African Trypanosomosis Caused by Trypanosoma congolense Infection" Pathogens 11, no. 3: 331. https://doi.org/10.3390/pathogens11030331