
The Relevance of TLR8 in Viral Infections


Abstract
:1. Introduction
2. Activation and Regulation of TLR8
3. TLR8 and Viral Infections

{kind=link}
| TLR | Name | Structure | Class | Ref. |
|---|---|---|---|---|
| TLR7 | Imiquimod |  | Imidazoquinoline amine analog to guanosine | [34,35] |
| Gardiquimod |  | Imidazoquinoline compound | [13,36,37] | |
| Loxoribine |  | Guanosine analogue | [38,39,40,41] | |
| Isatoribine |  | Nucleoside analogue | [42] | |
| TLR8 | TL8-506 |  | Benzoazepine compound | [43] |
| Selgantolimod |  | Nucleotide analogue | [44,45] | |
| TLR7/TLR8 | Resiquimod |  | Imidazoquinoline compound derivative | [24,38,40] |
| CL075 |  | Thiazoloquinolone derivative | [29,46,47,48] | |
| CL097 |  | Imidazoquinoline compound derivative | [48] |
| DNA Virus | ||
|---|---|---|
| Virus | Effect on TLR8 Activation | Cells |
| DNA Viruses | ||
| Vaccinia virus | Induces the expression of IFN-α, IFN-β | pDCs, HEK cells [21] |
| Hepatitis B virus | HBV infection inhibits innate immunity by decreasing TLR8 levels | PBMC [52] |
| RNA Viruses | ||
| Influenza A virus (H3N2) | Induces the expression of IL-8 | HEK cells, neutrophils [33] |
| Coxsackie B virus | Induces the expression of IFN-β, IL-6 | Human cardiac cells [4] |
| HIV-1 | Induces the expression of IL-6 and IL-1β | Human primary CD4+ T cells [50] |
| Colocalization of HIV and TLR8 is decreased | Dendritic cells [32] | |
| West Nile virus | TLR8 favors the infection in infected mice | Bone-marrow-derived dendritic cells (BMDCs) [27] |
| Zika virus | TLR8 and MyD88 expression decreases | Peripheral blood [53] |
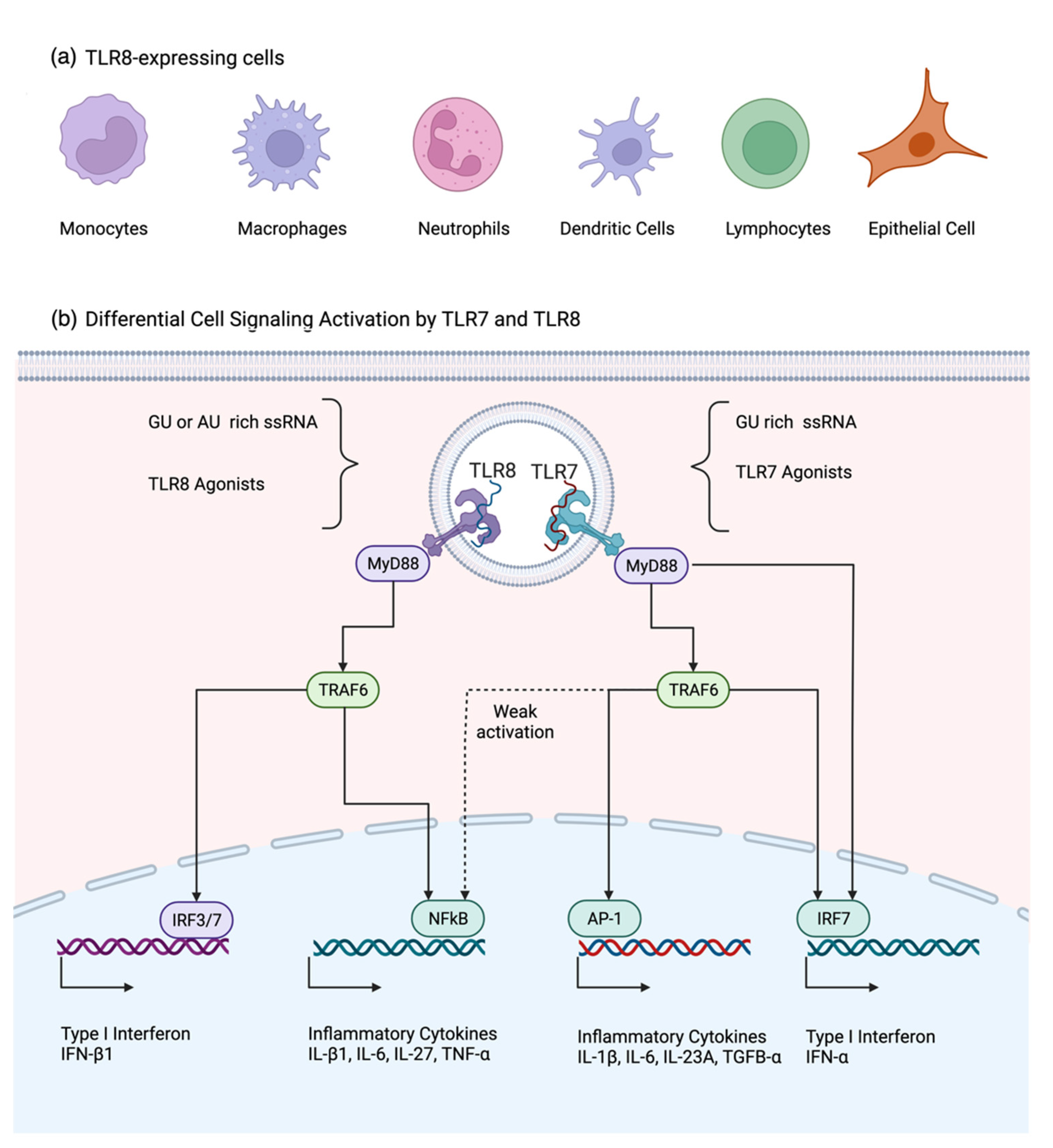
4. Signaling Pathways Associated with TLR8 and Its Differences with TLR7

5. Future Perspectives for Therapeutics with TLR8 Agonist for Viral Infections
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bendelja, K.; Vojvoda, V.; Aberle, N.; Cepin-Bogovic, J.; Gagro, A.; Mlinaric-Galinovic, G.; Rabatic, S. Decreased Toll-like Receptor 8 Expression and Lower TNF-α Synthesis in Infants with Acute RSV Infection. Respir. Res. 2010, 11, 143. [Google Scholar] [CrossRef] [Green Version]
- Leulier, F.; Lemaitre, B. Toll-like Receptors--Taking an Evolutionary Approach. Nat. Rev. Genet. 2008, 9, 165–178. [Google Scholar] [CrossRef] [Green Version]
- Kolli, D.; Velayutham, T.S.; Casola, A. Host-Viral Interactions: Role of Pattern Recognition Receptors (PRRs) in Human Pneumovirus Infections. Pathogens 2013, 2, 232–263. [Google Scholar] [CrossRef] [Green Version]
- Triantafilou, K.; Orthopoulos, G.; Vakakis, E.; Ahmed, M.A.E.; Golenbock, D.T.; Lepper, P.M.; Triantafilou, M. Human Cardiac Inflammatory Responses Triggered by Coxsackie B Viruses Are Mainly Toll-like Receptor (TLR) 8-Dependent. Cell. Microbiol. 2005, 7, 1117–1126. [Google Scholar] [CrossRef]
- Lin, Y.-T.; Verma, A.; Hodgkinson, C.P. Toll-Like Receptors and Human Disease: Lessons from Single Nucleotide Polymorphisms. Curr. Genom. 2012, 13, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Tomai, M.A.; Vasilakos, J.P. Chapter 8 - Toll-Like Receptor 7 and 8 Agonists for Vaccine Adjuvant Use. In Immunopotentiators in Modern Vaccines, 2nd ed.; Schijns, V.E.J.C., O’Hagan, D.T., Eds.; Academic Press: Cambridge, MA, USA, 2017; Chapter 8; pp. 149–162. ISBN 978-0-12-804019-5. [Google Scholar]
- Fore, F.; Indriputri, C.; Mamutse, J.; Nugraha, J. TLR10 and Its Unique Anti-Inflammatory Properties and Potential Use as a Target in Therapeutics. Immune Netw. 2020, 20, e21. [Google Scholar] [CrossRef]
- Bender, A.T.; Tzvetkov, E.; Pereira, A.; Wu, Y.; Kasar, S.; Przetak, M.M.; Vlach, J.; Niewold, T.B.; Jensen, M.A.; Okitsu, S.L. TLR7 and TLR8 Differentially Activate the IRF and NF-ΚB Pathways in Specific Cell Types to Promote Inflammation. ImmunoHorizons 2020, 4, 93–107. [Google Scholar] [CrossRef] [Green Version]
- Eng, H.-L.; Hsu, Y.-Y.; Lin, T.-M. Differences in TLR7/8 Activation between Monocytes and Macrophages. Biochem. Biophys. Res. Commun. 2018, 497, 319–325. [Google Scholar] [CrossRef]
- Maris, N.A.; Dessing, M.C.; de Vos, A.F.; Bresser, P.; van der Zee, J.S.; Jansen, H.M.; Spek, C.A.; Poll, T. van der Toll-like Receptor MRNA Levels in Alveolar Macrophages after Inhalation of Endotoxin. Eur. Respir. J. 2006, 28, 622–626. [Google Scholar] [CrossRef]
- Peng, G.; Guo, Z.; Kiniwa, Y.; Voo, K.S.; Peng, W.; Fu, T.; Wang, D.Y.; Li, Y.; Wang, H.Y.; Wang, R.-F. Toll-Like Receptor 8-Mediated Reversal of CD4+ Regulatory T Cell Function. Science 2005, 309, 1380–1384. [Google Scholar] [CrossRef]
- Ohto, U.; Tanji, H.; Shimizu, T. Structure and Function of Toll-like Receptor 8. Microbes Infect. 2014, 16, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Brownlie, R.; Liu, Q.; Babiuk, L.A.; Potter, A.; Mutwiri, G.K. Characterization of Bovine Toll-like Receptor 8: Ligand Specificity, Signaling Essential Sites and Dimerization. Mol. Immunol. 2009, 46, 978–990. [Google Scholar] [CrossRef]
- Tanji, H.; Ohto, U.; Motoi, Y.; Shibata, T.; Miyake, K.; Shimizu, T. Autoinhibition and Relief Mechanism by the Proteolytic Processing of Toll-like Receptor 8. Proc. Natl. Acad. Sci. USA 2016, 113, 3012–3017. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Xu, C.; Hsu, L.-C.; Luo, Y.; Xiang, R.; Chuang, T.-H. A Five-Amino-Acid Motif in the Undefined Region of the TLR8 Ectodomain Is Required for Species-Specific Ligand Recognition. Mol. Immunol. 2010, 47, 1083. [Google Scholar] [CrossRef] [Green Version]
- Pluta, L.; Yousefi, B.; Damania, B.; Khan, A.A. Endosomal TLR-8 Senses MicroRNA-1294 Resulting in the Production of NFḱB Dependent Cytokines. Front. Immunol. 2019, 10, 2860. [Google Scholar] [CrossRef]
- Zhang, Y.; El-Far, M.; Dupuy, F.P.; Abdel-Hakeem, M.S.; He, Z.; Procopio, F.A.; Shi, Y.; Haddad, E.K.; Ancuta, P.; Sekaly, R.-P.; et al. HCV RNA Activates APCs via TLR7/TLR8 While Virus Selectively Stimulates Macrophages Without Inducing Antiviral Responses. Sci. Rep. 2016, 6, 29447. [Google Scholar] [CrossRef] [Green Version]
- Forsbach, A.; Nemorin, J.-G.; Montino, C.; Müller, C.; Samulowitz, U.; Vicari, A.P.; Jurk, M.; Mutwiri, G.K.; Krieg, A.M.; Lipford, G.B.; et al. Identification of RNA Sequence Motifs Stimulating Sequence-Specific TLR8-Dependent Immune Responses. J Immunol. 2008, 180, 3729–3738. [Google Scholar] [CrossRef]
- Karikó, K.; Buckstein, M.; Ni, H.; Weissman, D. Suppression of RNA Recognition by Toll-like Receptors: The Impact of Nucleoside Modification and the Evolutionary Origin of RNA. Immunity 2005, 23, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Ugolini, M.; Gerhard, J.; Burkert, S.; Jensen, K.J.; Georg, P.; Ebner, F.; Volkers, S.M.; Thada, S.; Dietert, K.; Bauer, L.; et al. Recognition of Microbial Viability via TLR8 Drives TFH Cell Differentiation and Vaccine Responses. Nat. Immunol. 2018, 19, 386–396. [Google Scholar] [CrossRef]
- Martinez, J.; Huang, X.; Yang, Y. Toll-like Receptor 8-Mediated Activation of Murine Plasmacytoid Dendritic Cells by Vaccinia Viral DNA. Proc. Natl. Acad. Sci. USA 2010, 107, 6442–6447. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.; et al. MicroRNAs Bind to Toll-like Receptors to Induce Prometastatic Inflammatory Response. Proc Natl Acad Sci USA 2012, 109, E2110–E2116. [Google Scholar] [CrossRef] [Green Version]
- Bernard, M.A.; Han, X.; Inderbitzin, S.; Agbim, I.; Zhao, H.; Koziel, H.; Tachado, S.D. HIV-Derived SsRNA Binds to TLR8 to Induce Inflammation-Driven Macrophage Foam Cell Formation. PLoS ONE 2014, 9, e104039. [Google Scholar] [CrossRef] [Green Version]
- Hackstein, H.; Knoche, A.; Nockher, A.; Poeling, J.; Kubin, T.; Jurk, M.; Vollmer, J.; Bein, G. The TLR7/8 Ligand Resiquimod Targets Monocyte-Derived Dendritic Cell Differentiation via TLR8 and Augments Functional Dendritic Cell Generation. Cell. Immunol. 2011, 271, 401–412. [Google Scholar] [CrossRef] [PubMed]
- de Marcken, M.; Dhaliwal, K.; Danielsen, A.C.; Gautron, A.S.; Dominguez-Villar, M. TLR7 and TLR8 Activate Distinct Pathways in Monocytes during RNA Virus Infection. Sci. Signal. 2019, 12. [Google Scholar] [CrossRef]
- Wang, J.; Shao, Y.; Bennett, T.A.; Shankar, R.A.; Wightman, P.D.; Reddy, L.G. The Functional Effects of Physical Interactions among Toll-like Receptors 7, 8, and 9*. J. Biol. Chem. 2006, 281, 37427–37434. [Google Scholar] [CrossRef] [Green Version]
- Paul, A.M.; Acharya, D.; Le, L.; Wang, P.; Stokic, D.S.; Leis, A.A.; Alexopoulou, L.; Town, T.; Flavell, R.A.; Fikrig, E.; et al. TLR8 Couples SOCS-1 and Restrains TLR7-Mediated Antiviral Immunity, Exacerbating West Nile Virus Infection in Mice. J. Immunol. 2016, 197, 4425–4435. [Google Scholar] [CrossRef] [Green Version]
- Odoardi, N.; Kourko, O.; Petes, C.; Basta, S.; Gee, K. TLR7 Ligation Inhibits TLR8 Responsiveness in IL-27-Primed Human THP-1 Monocytes and Macrophages. J. Innate Immun. 2021, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-H.; Eng, H.-L.; Lin, K.-H.; Chang, C.-H.; Hsieh, C.-A.; Lin, Y.-L.; Lin, T.-M. TLR7 and TLR8 Gene Variations and Susceptibility to Hepatitis C Virus Infection. PLOS ONE 2011, 6, e26235. [Google Scholar] [CrossRef] [Green Version]
- Awais, M.; Wang, K.; Lin, X.; Qian, W.; Zhang, N.; Wang, C.; Wang, K.; Zhao, L.; Fu, Z.F.; Cui, M. TLR7 Deficiency Leads to TLR8 Compensative Regulation of Immune Response against JEV in Mice. Front. Immunol. 2017, 8, 160. [Google Scholar] [CrossRef] [Green Version]
- Triantafilou, K.; Vakakis, E.; Orthopoulos, G.; Ahmed, M.A.E.; Schumann, C.; Lepper, P.M.; Triantafilou, M. TLR8 and TLR7 Are Involved in the Host’s Immune Response to Human Parechovirus 1. Eur. J. Immunol. 2005, 35, 2416–2423. [Google Scholar] [CrossRef]
- Khatamzas, E.; Hipp, M.M.; Gaughan, D.; Pichulik, T.; Leslie, A.; Fernandes, R.A.; Muraro, D.; Booth, S.; Zausmer, K.; Sun, M.; et al. Snapin Promotes HIV-1 Transmission from Dendritic Cells by Dampening TLR8 Signaling. EMBO J. 2017, 36, 2998–3011. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.P.; Bowen, G.N.; Padden, C.; Cerny, A.; Finberg, R.W.; Newburger, P.E.; Kurt-Jones, E.A. Toll-like Receptor–Mediated Activation of Neutrophils by Influenza A Virus. Blood 2008, 112, 2028–2034. [Google Scholar] [CrossRef] [PubMed]
- Larangé, A.; Antonios, D.; Pallardy, M.; Kerdine-Römer, S. TLR7 and TLR8 Agonists Trigger Different Signaling Pathways for Human Dendritic Cell Maturation. J. of Leukoc. Biol. 2009, 85, 673–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craft, N.; Bruhn, K.W.; Nguyen, B.D.; Prins, R.; Lin, J.W.; Liau, L.M.; Miller, J.F. The TLR7 Agonist Imiquimod Enhances the Anti-Melanoma Effects of a Recombinant Listeria Monocytogenes Vaccine. J. Immunol. 2005, 175, 1983–1990. [Google Scholar] [CrossRef] [Green Version]
- Buitendijk, M.; Eszterhas, S.K.; Howell, A.L. Gardiquimod: A Toll-Like Receptor-7 Agonist That Inhibits HIV Type 1 Infection of Human Macrophages and Activated T Cells. AIDS Res. Hum. Retrovir. 2013, 29, 907–918. [Google Scholar] [CrossRef] [Green Version]
- Enosi Tuipulotu, D.; Netzler, N.E.; Lun, J.H.; Mackenzie, J.M.; White, P.A. TLR7 Agonists Display Potent Antiviral Effects against Norovirus Infection via Innate Stimulation. Antimicrob. Agents and Chemother. 2018, 62, e02417-17. [Google Scholar] [CrossRef] [Green Version]
- Jurk, M.; Kritzler, A.; Schulte, B.; Tluk, S.; Schetter, C.; Krieg, A.M.; Vollmer, J. Modulating Responsiveness of Human TLR7 and 8 to Small Molecule Ligands with T-Rich Phosphorothiate Oligodeoxynucleotides. Eur. J. Immunol. 2006, 36, 1815–1826. [Google Scholar] [CrossRef]
- Dzopalic, T.; Dragicevic, A.; Vasilijic, S.; Vucevic, D.; Majstorovic, I.; Bozic, B.; Balint, B.; Colic, M. Loxoribine, a Selective Toll-like Receptor 7 Agonist, Induces Maturation of Human Monocyte-Derived Dendritic Cells and Stimulates Their Th-1- and Th-17-Polarizing Capability. Int. Immunopharmacol. 2010, 10, 1428–1433. [Google Scholar] [CrossRef] [PubMed]
- Heil, F.; Ahmad-Nejad, P.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Gellert, T.; Dietrich, H.; Lipford, G.; Takeda, K.; Akira, S.; et al. The Toll-like Receptor 7 (TLR7)-Specific Stimulus Loxoribine Uncovers a Strong Relationship within the TLR7, 8 and 9 Subfamily. Eur. J. Immunol. 2003, 33, 2987–2997. [Google Scholar] [CrossRef]
- Xu, N.; Yao, H.-P.; Lv, G.-C.; Chen, Z. Downregulation of TLR7/9 Leads to Deficient Production of IFN-α from Plasmacytoid Dendritic Cells in Chronic Hepatitis B. Inflamm. Res. 2012, 61, 997–1004. [Google Scholar] [CrossRef]
- Horsmans, Y.; Berg, T.; Desager, J.-P.; Mueller, T.; Schott, E.; Fletcher, S.P.; Steffy, K.R.; Bauman, L.A.; Kerr, B.M.; Averett, D.R. Isatoribine, an Agonist of TLR7, Reduces Plasma Virus Concentration in Chronic Hepatitis C Infection. Hepatology 2005, 42, 724–731. [Google Scholar] [CrossRef]
- Serrano, R.; Wesch, D.; Kabelitz, D. Activation of Human Γδ T Cells: Modulation by Toll-Like Receptor 8 Ligands and Role of Monocytes. Cells 2020, 9, 713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackman, R.L.; Mish, M.; Chin, G.; Perry, J.K.; Appleby, T.; Aktoudianakis, V.; Metobo, S.; Pyun, P.; Niu, C.; Daffis, S.; et al. Discovery of GS-9688 (Selgantolimod) as a Potent and Selective Oral Toll-Like Receptor 8 Agonist for the Treatment of Chronic Hepatitis B. J. Med. Chem. 2020, 63, 10188–10203. [Google Scholar] [CrossRef] [PubMed]
- Amin, O.E.; Colbeck, E.J.; Daffis, S.; Khan, S.; Ramakrishnan, D.; Pattabiraman, D.; Chu, R.; Micolochick Steuer, H.; Lehar, S.; Peiser, L.; et al. Therapeutic Potential of TLR8 Agonist GS-9688 (Selgantolimod) in Chronic Hepatitis B: Remodeling of Antiviral and Regulatory Mediators. Hepatology 2021, 74, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Wu, Z.; Ren, S.; Wei, Y.; Gao, M.; Randolph, G.J.; Qu, C. TLR8 Agonists Stimulate Newly Recruited Monocyte-Derived Cells into Potent APCs That Enhance HBsAg Immunogenicity. Vaccine 2010, 28, 6273–6281. [Google Scholar] [CrossRef] [Green Version]
- Chatillon, J.-F.; Hamieh, M.; Bayeux, F.; Abasq, C.; Fauquembergue, E.; Drouet, A.; Guisier, F.; Latouche, J.-B.; Musette, P. Direct Toll-Like Receptor 8 Signaling Increases the Functional Avidity of Human CD8+ T Lymphocytes Generated for Adoptive T Cell Therapy Strategies. Immun. Inflamm. Dis. 2015, 3, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, S.; Li, T.; Lv, X.; Zhang, M.; Zang, G.; Qi, C.; Liu, Y.-J.; Xu, L.; Chen, J. PDC Activation by TLR7/8 Ligand CL097 Compared to TLR7 Ligand IMQ or TLR9 Ligand CpG. J. Immunol. Res. 2019, 2019, 1749803. [Google Scholar] [CrossRef] [Green Version]
- Song, B.M.; Kang, Y.M.; Kim, H.S.; Seo, S.H. Induction of Inflammatory Cytokines and Toll-like Receptors in Human Normal Respiratory Epithelial Cells Infected with Seasonal H1N1, 2009 Pandemic H1N1, Seasonal H3N2, and Highly Pathogenic H5N1 Influenza Virus. Viral Immunol. 2011, 24, 179–187. [Google Scholar] [CrossRef]
- Meås, H.Z.; Haug, M.; Beckwith, M.S.; Louet, C.; Ryan, L.; Hu, Z.; Landskron, J.; Nordbø, S.A.; Taskén, K.; Yin, H.; et al. Sensing of HIV-1 by TLR8 Activates Human T Cells and Reverses Latency. Nat. Commun. 2020, 11, 147. [Google Scholar] [CrossRef]
- Campbell, G.R.; Rawat, P.; Bruckman, R.S.; Spector, S.A. Human Immunodeficiency Virus Type 1 Nef Inhibits Autophagy through Transcription Factor EB Sequestration. PLoS Pathog. 2015, 11, e1005018. [Google Scholar] [CrossRef] [Green Version]
- Deng, G.; Ge, J.; Liu, C.; Pang, J.; Huang, Z.; Peng, J.; Sun, J.; Hou, J.; Zhang, X. Impaired Expression and Function of TLR8 in Chronic HBV Infection and Its Association with Treatment Responses during Peg-IFN-α-2a Antiviral Therapy. Clin. Res. Hepatol. Gastroenterol. 2017, 41, 386–398. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.H.M.; Moises, R.N.C.; Alves, B.E.B.; Pereira, H.W.B.; de Paiva, A.A.P.; Morais, I.C.; Nascimento, Y.M.; Monteiro, J.D.; de Souto, J.T.; Nascimento, M.S.L.; et al. Innate Immune Response in Patients with Acute Zika Virus Infection. Med. Microbiol. Immunol. 2019, 208, 703–714. [Google Scholar] [CrossRef]
- Oh, D.-Y.; Taube, S.; Hamouda, O.; Kücherer, C.; Poggensee, G.; Jessen, H.; Eckert, J.K.; Neumann, K.; Storek, A.; Pouliot, M.; et al. A Functional Toll-Like Receptor 8 Variant Is Associated with HIV Disease Restriction. J. Infect. Dis. 2008, 198, 701–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahallawi, W.H.; Suliman, B.A. TLR8 Is Highly Conserved among the Saudi Population and Its Mutations Have No Effect on the Severity of COVID-19 Symptoms. Am. J. Clin. Exp. Immunol. 2021, 10, 71–76. [Google Scholar] [PubMed]
- Gorden, K.B.; Gorski, K.S.; Gibson, S.J.; Kedl, R.M.; Kieper, W.C.; Qiu, X.; Tomai, M.A.; Alkan, S.S.; Vasilakos, J.P. Synthetic TLR Agonists Reveal Functional Differences between Human TLR7 and TLR8. J. Immunol. 2005, 174, 1259–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornung, V.; Guenthner-Biller, M.; Bourquin, C.; Ablasser, A.; Schlee, M.; Uematsu, S.; Noronha, A.; Manoharan, M.; Akira, S.; de Fougerolles, A.; et al. Sequence-Specific Potent Induction of IFN-α by Short Interfering RNA in Plasmacytoid Dendritic Cells through TLR7. Nat. Med. 2005, 11, 263–270. [Google Scholar] [CrossRef]
- Kawai, T.; Sato, S.; Ishii, K.J.; Coban, C.; Hemmi, H.; Yamamoto, M.; Terai, K.; Matsuda, M.; Inoue, J.; Uematsu, S.; et al. Interferon-α Induction through Toll-like Receptors Involves a Direct Interaction of IRF7 with MyD88 and TRAF6. Nat. Immunol. 2004, 5, 1061–1068. [Google Scholar] [CrossRef]
- Diebold, S.S.; Kaisho, T.; Hemmi, H.; Akira, S.; Sousa, C.R. e Innate Antiviral Responses by Means of TLR7-Mediated Recognition of Single-Stranded RNA. Science 2004, 303, 1529–1531. [Google Scholar] [CrossRef]
- Aryan, Z.; Holgate, S.T.; Radzioch, D.; Rezaei, N. A New Era of Targeting the Ancient Gatekeepers of the Immune System: Toll-Like Agonists in the Treatment of Allergic Rhinitis and Asthma. Int. Arch. Allergy Immunol. 2014, 164, 46–63. [Google Scholar] [CrossRef]
- Wille-Reece, U.; Flynn, B.J.; Loré, K.; Koup, R.A.; Kedl, R.M.; Mattapallil, J.J.; Weiss, W.R.; Roederer, M.; Seder, R.A. HIV Gag Protein Conjugated to a Toll-like Receptor 7/8 Agonist Improves the Magnitude and Quality of Th1 and CD8+ T Cell Responses in Nonhuman Primates. Proc. Natl. Acad. Sci. USA 2005, 102, 15190–15194. [Google Scholar] [CrossRef] [Green Version]
- Jangra, S.; De Vrieze, J.; Choi, A.; Rathnasinghe, R.; Laghlali, G.; Uvyn, A.; Van Herck, S.; Nuhn, L.; Deswarte, K.; Zhong, Z.; et al. Sterilizing Immunity against SARS-CoV-2 Infection in Mice by a Single-Shot and Lipid Amphiphile Imidazoquinoline TLR7/8 Agonist-Adjuvanted Recombinant Spike Protein Vaccine**. Angew. Chem. Int. Ed. 2021, 60, 9467–9473. [Google Scholar] [CrossRef] [PubMed]
- Lissina, A.; Briceño, O.; Afonso, G.; Larsen, M.; Gostick, E.; Price, D.A.; Mallone, R.; Appay, V. Priming of Qualitatively Superior Human Effector CD8+ T Cells Using TLR8 Ligand Combined with FLT3 Ligand. J. Immunol. 2016, 196, 256–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Espinoza, I.; Guerrero-Plata, A. The Relevance of TLR8 in Viral Infections. Pathogens 2022, 11, 134. https://doi.org/10.3390/pathogens11020134
Martínez-Espinoza I, Guerrero-Plata A. The Relevance of TLR8 in Viral Infections. Pathogens. 2022; 11(2):134. https://doi.org/10.3390/pathogens11020134
Chicago/Turabian StyleMartínez-Espinoza, Iván, and Antonieta Guerrero-Plata. 2022. "The Relevance of TLR8 in Viral Infections" Pathogens 11, no. 2: 134. https://doi.org/10.3390/pathogens11020134