
Biofumigation for the Management of Fusarium graminearum in a Wheat-Maize Rotation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Set Up
2.2. Semi-Artificial Infection of Wheat
2.3. Biofumigants-Biomass, Glucosinolate Analysis, Incorporation
2.4. Assessment of Fusarium graminearum Inoculum Buried in Sachets Post-Incorporation of Brassica Biofumigants
2.5. Maize Assessment
2.5.1. Disease Assessment and Fusarium graminearum Incidence
2.5.2. Mycotoxin and Molecular Analysis
2.6. Statistical Analysis
3. Results
3.1. Fusarium graminearum in Wheat
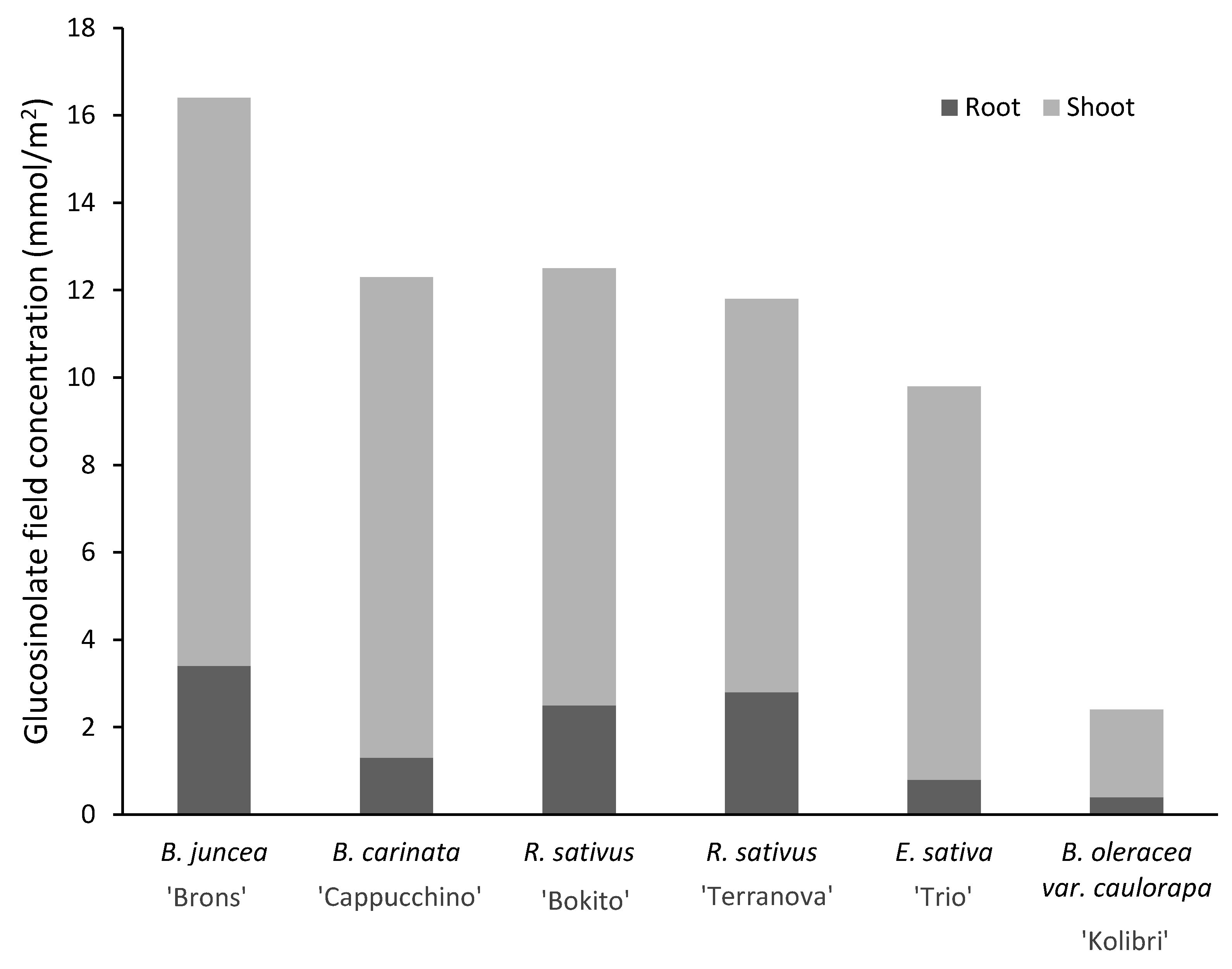
3.2. Biomass, Glucosinolate Concentrations of Brassicas
3.3. Effect of Brassica Incorporation on F. graminearum Inoculum in Sachets
3.4. Effect of Brassica Biofumigation on Fusarium graminearum Diseases in a Subsequent Maize Crop
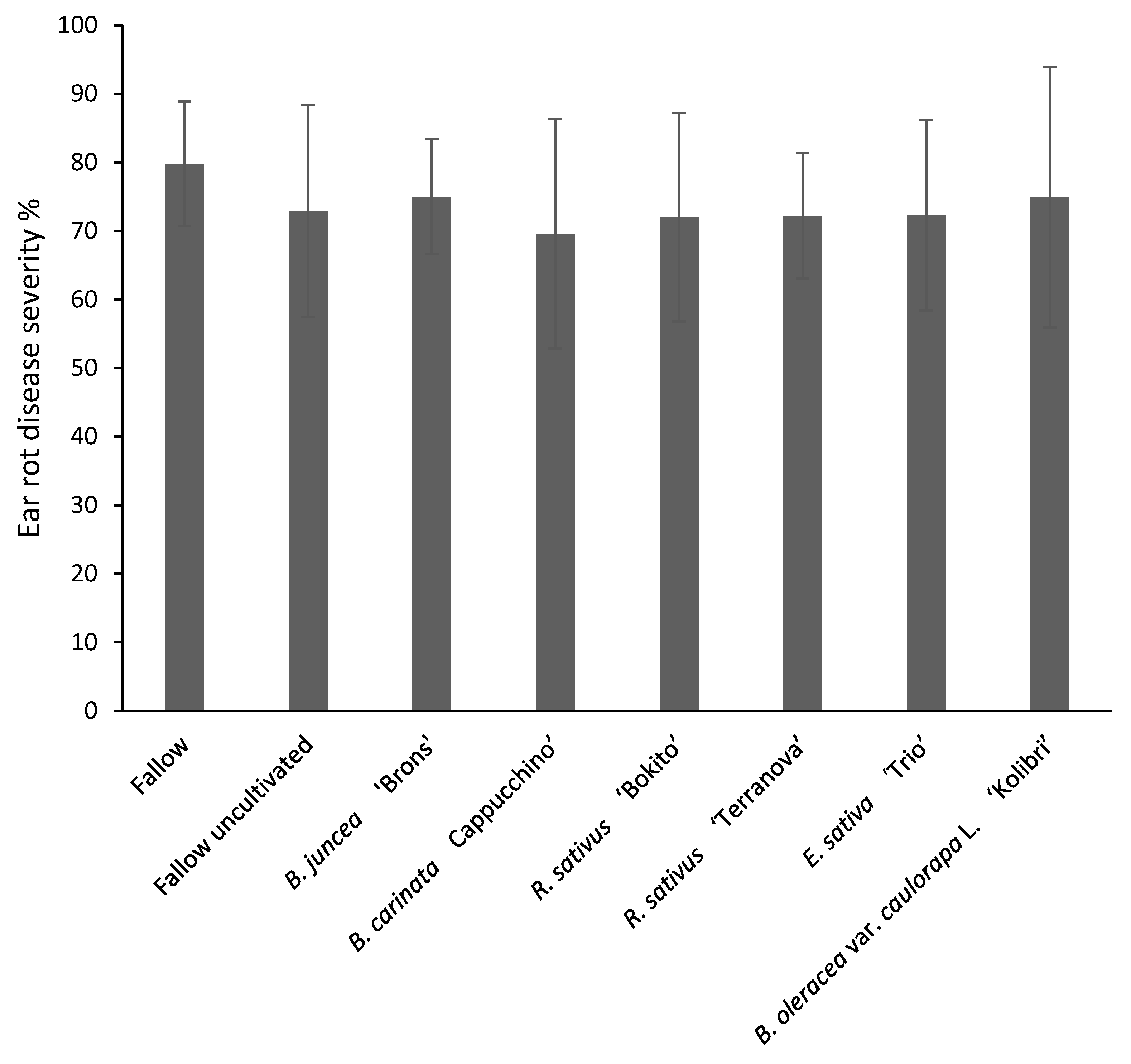
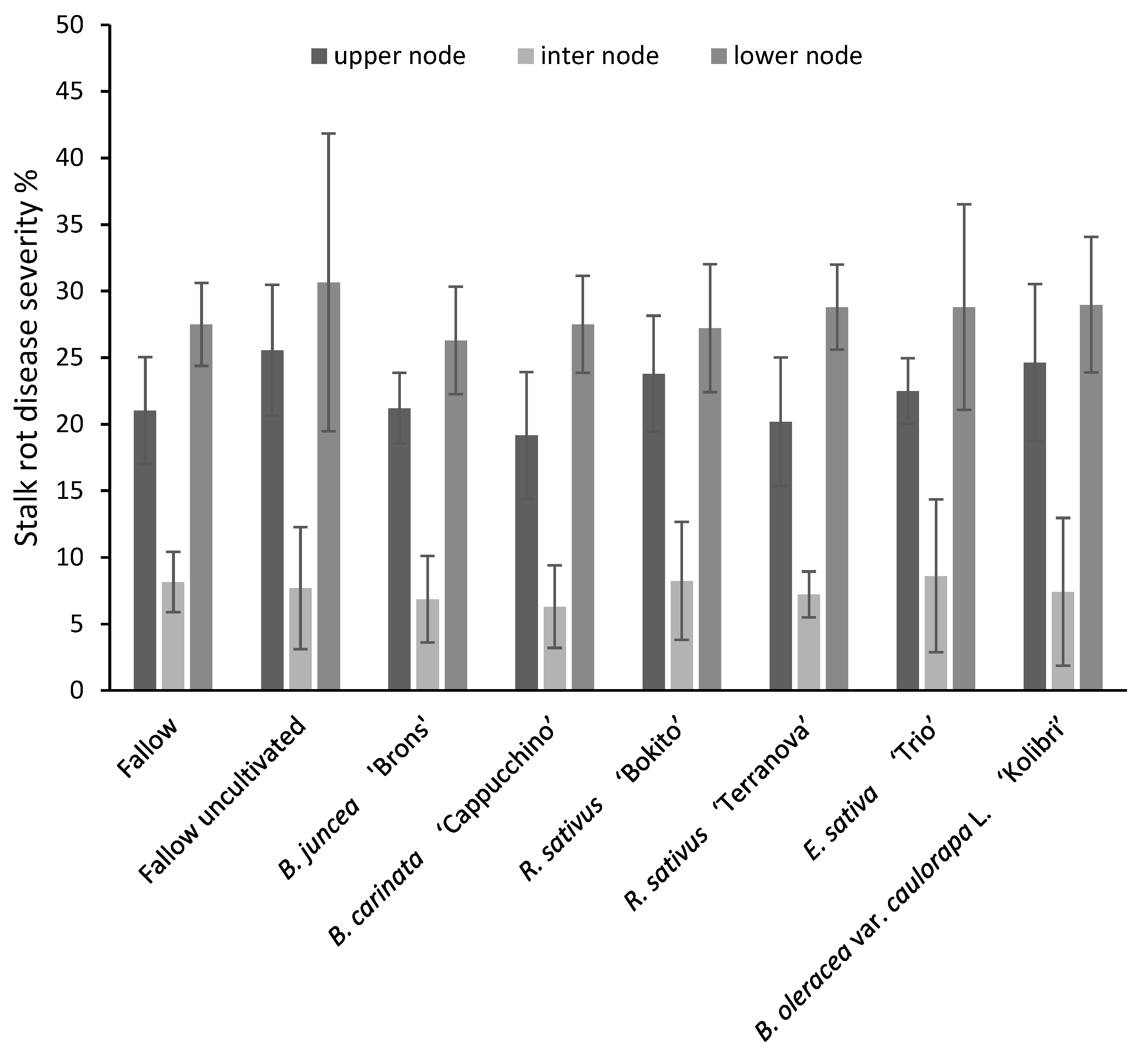
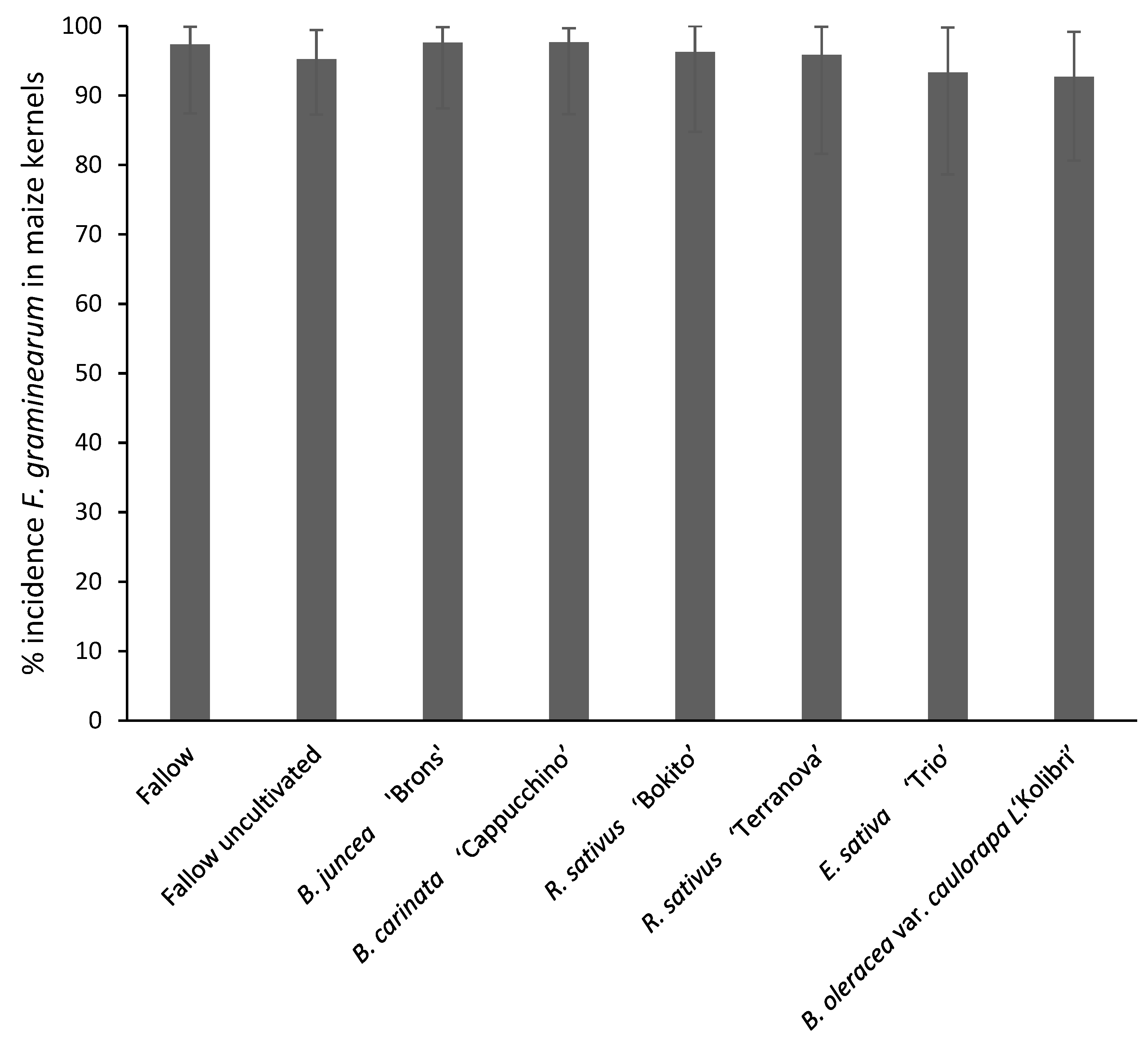
3.4.1. Disease Severity and Fusarium graminearum Incidence in Maize
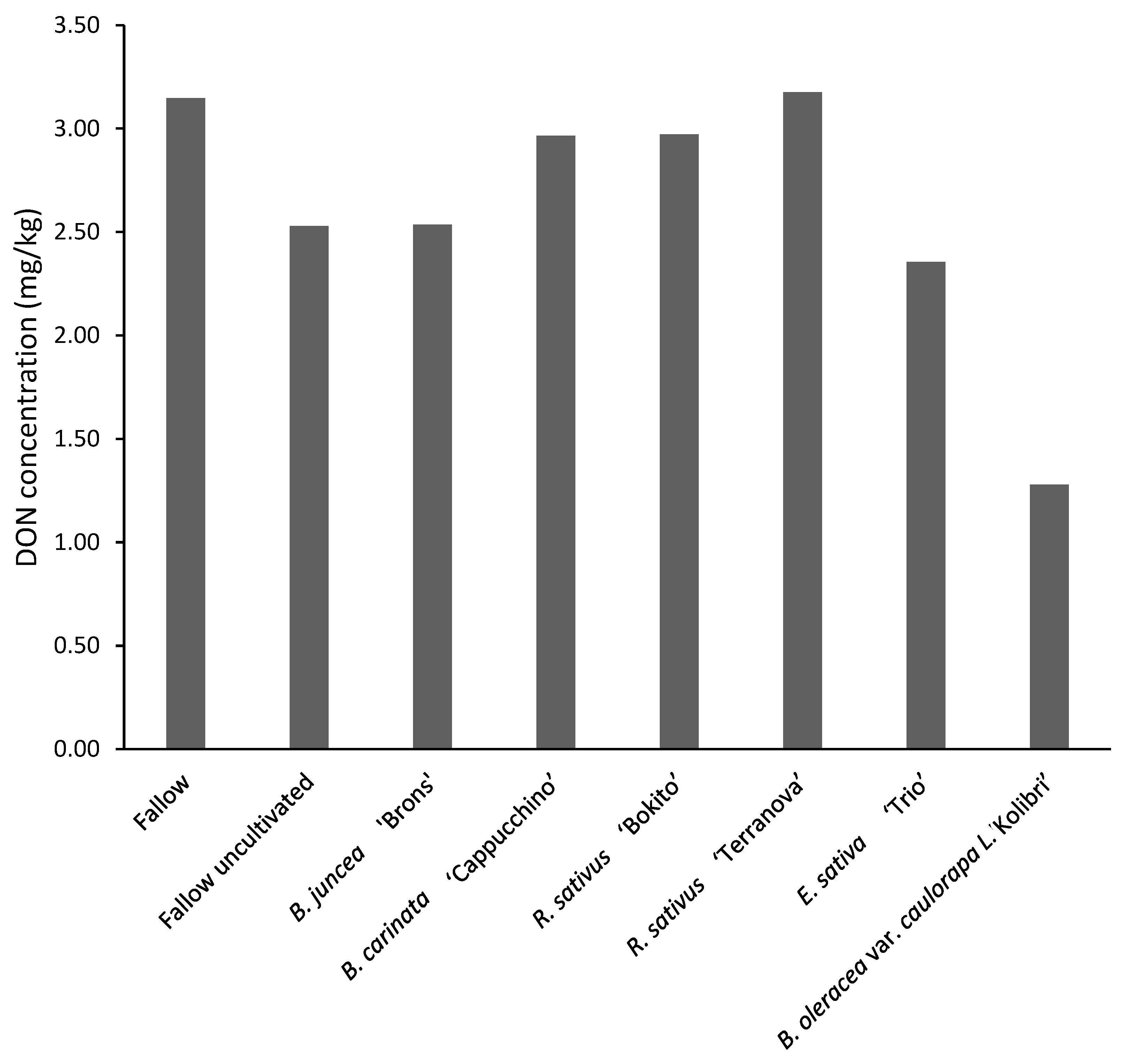
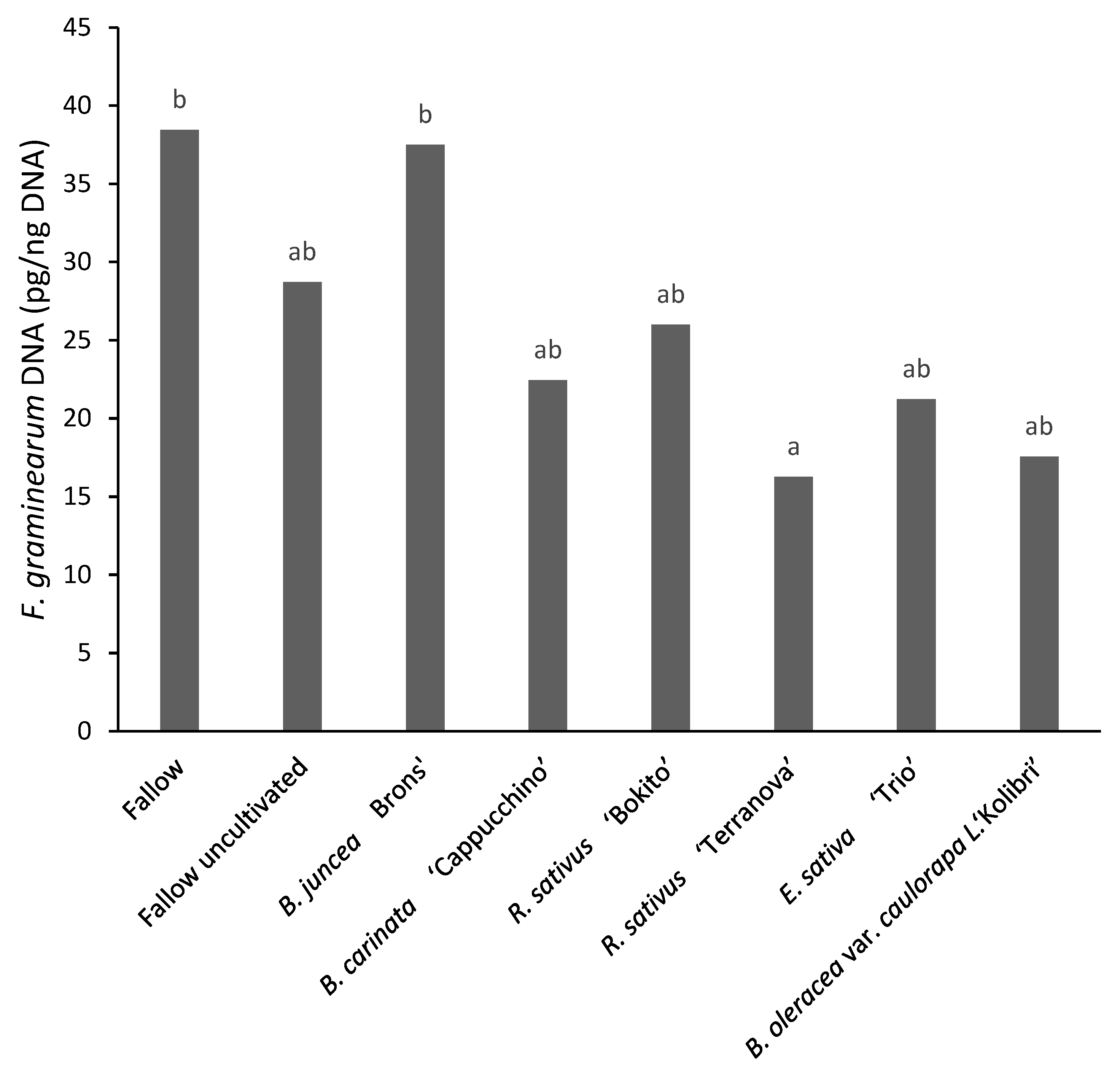
3.4.2. Assessment of Deoxynivalenol and Fusarium graminearum DNA in Maize
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- OECD-FAO Agricultural Outlook OECD Agriculture Statistics (Database). Available online: https://www.oecd-ilibrary.org/agriculture-and-food/data/oecd-agriculture-statistics_agr-data-en (accessed on 3 October 2021).
- FAOSTAT Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 14 November 2022).
- McMullen, M.; Bergstrom, G.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A unified effort to fight an enemy of wheat and barley: Fusarium head blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munkvold, G.P. Epidemiology of Fusarium diseases and their mycotoxins in maize ears. Eur. J. Plant Pathol. 2003, 109, 705–713. [Google Scholar] [CrossRef]
- CAST, Mycotoxins: Risks in Plant, Animal, and Human Systems; Task Force Report No. 139; Council for Agricultural Science and Technology: Ames, IA, USA, 2003.
- Guo, H.; Ji, J.; Wang, J.s.; Sun, X. Deoxynivalenol: Masked forms, fate during food processing, and potential biological remedies. Compr. Rev. Food Sci. Food Saf. 2020, 19, 895–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szuets, P.; Mesterhazy, A.; Falkay, G.Y.; Bartok, T. Early telarche symptoms in children and their relations to zearalenone contamination in foodstuffs. Cereal Res. Commun. 1997, 25, 429–436. [Google Scholar] [CrossRef]
- Ropejko, K.; Twarużek, M. Zearalenone and Its Metabolites—General Overview, Occurrence, and Toxicity. Toxins 2021, 13, 35. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.; Dahl, B.; Nganje, W. Economic costs of Fusarium Head Blight, scab and deoxynivalenol. World Mycotoxin J. 2018, 11, 291–302. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 1126/2007 of 28 September 2007 amending Regulation (EC) No 1881/2006 setting maximum levels for certain contaminants in foodstuffs as regards Fusarium toxins in maize and maize products. Off. J. Eur. Union 2007, L 255, 14–17.
- Edwards, S.G.; Jennings, P. Impact of agronomic factors on Fusarium mycotoxins in harvested wheat. Food Addit. Contam. Part A 2018, 35, 2443–2454. [Google Scholar] [CrossRef]
- Pleadin, J.; Vahčić, N.; Perši, N.; Ševelj, D.; Markov, K.; Frece, J. Fusarium mycotoxins’ occurrence in cereals harvested from Croatian fields. Food Control 2013, 32, 49–54. [Google Scholar] [CrossRef]
- Pallez-Barthel, M.; Cocco, E.; Vogelgsang, S.; Beyer, M. Frequency of Deoxynivalenol Concentrations above the Maximum Limit in Raw Winter Wheat Grain during a 12-Year Multi-Site Survey. Agronomy 2021, 11, 960. [Google Scholar] [CrossRef]
- Blandino, M.; Pilati, A.; Reyneri, A.; Scudellari, D. Effect of maize crop residue density on Fusarium head blight and on deoxynivalenol contamination of common wheat grains. Cereal Res. Commun. 2010, 38, 550–559. [Google Scholar] [CrossRef]
- Champeil, A.; Fourbet, J.-F.; Doré, T.; Rossignol, L. Influence of cropping system on Fusarium head blight and mycotoxin levels in winter wheat. Crop Prot. 2004, 23, 531–537. [Google Scholar] [CrossRef]
- Leplat, J.; Friberg, H.; Abid, M.; Steinberg, C. Survival of Fusarium graminearum, the causal agent of Fusarium head blight. A review. Agron. Sustain. Dev. 2013, 33, 97–111. [Google Scholar] [CrossRef]
- Shah, L.; Ali, A.; Yahya, M.; Zhu, Y.; Wang, S.; Si, H.; Rahman, H.; Ma, C. Integrated control of fusarium head blight and deoxynivalenol mycotoxin in wheat. Plant Pathol. 2018, 67, 532–548. [Google Scholar] [CrossRef]
- Yuen, G.Y.; Schoneweis, S.D. Strategies for managing Fusarium head blight and deoxynivalenol accumulation in wheat. Int. J. Food Microbiol. 2007, 119, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Pesticides and You: While France Bans a Common Endocrine Disrupting Pesticide, EPA Goes Silent. 2019, Volume 39, pp. 9–14. Available online: https://www.beyondpesticides.org/assets/media/documents/Full%20Summer%202019%20PAY.pdf (accessed on 26 November 2022).
- Becher, R.; Hettwer, U.; Karlovsky, P.; Deising, H.B.; Wirsel, S.G.R. Adaptation of Fusarium graminearum to tebuconazole yielded descendants diverging for levels of fitness, fungicide resistance, virulence, and mycotoxin production. Phytopathology 2010, 100, 444–453. [Google Scholar] [CrossRef] [Green Version]
- Kirkegaard, J.A.; Gardner, P.A.; Desmarchelier, J.M.; Angus, J.F. Biofumigation using Brassica species to control pests and diseases in horticulture and agriculture. In 9th Australian Research Assembly on Brassicas; Wratten, M., Mailer, R.J., Eds.; British Society for Plant Pathology, Agricultural Research Institute: Wagga Wagga, NSW, Australia, 1993; pp. 77–82. [Google Scholar]
- Kirkegaard, J.; Matthiessen, J. Developing and refining the biofumigation concept. Agroindustria 2004, 3, 233–239. [Google Scholar]
- Motisi, N.; Doré, T.; Lucas, P.; Montfort, F. Dealing with the variability in biofumigation efficacy through an epidemiological framework. Soil Biol. Biochem. 2010, 42, 2044–2057. [Google Scholar] [CrossRef]
- Tagele, S.B.; Kim, R.; Shin, J. Interactions between Brassica Biofumigants and Soil Microbiota: Causes and Impacts. J. Agric. Food Chem. 2021, 69, 11538–11553. [Google Scholar] [CrossRef]
- Larkin, R.P.; Griffin, T.S. Control of soilborne potato diseases using Brassica green manures. Crop Prot. 2007, 26, 1067–1077. [Google Scholar] [CrossRef]
- Drakopoulos, D.; Kägi, A.; Gimeno, A.; Six, J.; Jenny, E.; Forrer, H.-R.; Musa, T.; Meca, G.; Vogelgsang, S. Prevention of Fusarium head blight infection and mycotoxins in wheat with cut-and-carry biofumigation and botanicals. Field Crops Res. 2020, 246, 107681. [Google Scholar] [CrossRef]
- Ashiq, S.; Edwards, S.G.; Fatukasi, O.; Watson, A.; Back, M.A. In vitro activity of isothiocyanates against Fusarium graminearum. Plant Pathol. 2022, 71, 594–601. [Google Scholar] [CrossRef]
- Ashiq, S.; Edwards, S.; Watson, A.; Blundell, E.; Back, M. Antifungal effect of brassica tissues on the mycotoxigenic cereal pathogen Fusarium graminearum. Antibiotics 2022, 11, 1249. [Google Scholar] [CrossRef]
- Nicholson, P.; Simpson, D.R.; Weston, G.; Rezanoor, H.N.; Lees, A.K.; Parry, D.W.; Joyce, D. Detection and quantification of Fusarium culmorum and Fusarium graminearum in cereals using PCR assays. Physiol. Mol. Plant Pathol. 1998, 53, 17–37. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Hofgaard, I.S.; Seehusen, T.; Aamot, H.U.; Riley, H.; Razzaghian, J.; Le, V.H.; Hjelkrem, A.-G.R.; Dill-Macky, R.; Brodal, G. Inoculum potential of Fusarium spp. relates to tillage and straw management in Norwegian fields of spring oats. Front. Microbiol. 2016, 7, 556. [Google Scholar] [CrossRef] [Green Version]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Ames, IA, USA, 2006. [Google Scholar]
- Ngala, B.M.; Haydock, P.P.J.; Woods, S.; Back, M.A. Biofumigation with Brassica juncea, Raphanus sativus and Eruca sativa for the management of field populations of the potato cyst nematode Globodera pallida. Pest Manag. Sci. 2015, 71, 759–769. [Google Scholar] [CrossRef]
- ISO 9167; Rapeseed and Rapeseed Meals—Determination of Glucosinolates Content—Method Using HPLC. ISO: Geneva, Switzerland, 2019. Available online: https://www.iso.org/standard/72207.html (accessed on 21 March 2022).
- Masiello, M.; Somma, S.; Ghionna, V.; Logrieco, A.F.; Moretti, A. In vitro and in field response of different fungicides against Aspergillus flavus and Fusarium species causing ear rot disease of maize. Toxins 2019, 11, 11. [Google Scholar] [CrossRef] [Green Version]
- McKinney, H.H. Influence of soil temperature and moisture on infection of wheat seedlings by Helminthosporium sativum. J. Agric. Res. 1923, 192326, 195–217. [Google Scholar]
- Edwards, S.G.; Pirgozliev, S.R.; Hare, M.C.; Jenkinson, P. Quantification of trichothecene-producing Fusarium species in harvested grain by competitive PCR to determine efficacies of fungicides against Fusarium head blight of winter wheat. Appl. Environ. Microbiol. 2001, 67, 1575–1580. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Imathiu, S.M.; Ray, R.V.; Back, M.I.; Hare, M.C.; Edwards, S.G. A survey investigating the infection of Fusarium langsethiae and production of HT-2 and T-2 mycotoxins in UK oat fields. J. Phytopathol. 2013, 161, 553–561. [Google Scholar] [CrossRef]
- Subbarao, K.V.; Hubbard, J.C.; Koike, S.T. Evaluation of broccoli residue incorporation into field soil for Verticillium wilt control in cauliflower. Plant Dis. 1999, 83, 124–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blok, W.J.; Lamers, J.G.; Termorshuizen, A.J.; Bollen, G.J. Control of soilborne plant pathogens by incorporating fresh organic amendments followed by tarping. Phytopathology 2000, 90, 253–259. [Google Scholar] [CrossRef]
- Njoroge, S.M.C.; Riley, M.B.; Keinath, A.P. Effect of incorporation of Brassica spp. residues on population densities of soilborne microorganisms and on damping-off and Fusarium wilt of watermelon. Plant Dis. 2008, 92, 287–294. [Google Scholar] [CrossRef] [Green Version]
- Price, A.J.; Charron, C.S.; Saxton, A.M.; Sams, C.E. Allyl isothiocyanate and carbon dioxide produced during degradation of Brassica juncea tissue in different soil conditions. Hort. Sci. 2005, 40, 1734–1739. [Google Scholar] [CrossRef] [Green Version]
- Gamliel, A.; Stapleton, J.J. Improvement of soil solarization with volatile compounds generated from organic amendments. Phytoparasitica 1997, 25, S31–S38. [Google Scholar] [CrossRef]
- Met Office, Historic Station Data. Available online: https://www.metoffice.gov.uk/research/climate/maps-and-data/historic-station-data (accessed on 26 October 2021).
- Morra, M.J.; Kirkegaard, J.A. Isothiocyanate release from soil-incorporated Brassica tissues. Soil Biol. Biochem. 2002, 34, 1683–1690. [Google Scholar] [CrossRef]
- Matthiessen, J.N.; Warton, B.; Shackleton, M.A. The importance of plant maceration and water addition in achieving high Brassica-derived isothiocyanate levels in soil. Agroindustria 2004, 3, 277–280. [Google Scholar]
- Wang, L.; Mazzola, M. Effect of soil physical conditions on emission of allyl isothiocyanate and subsequent microbial inhibition in response to Brassicaceae seed meal amendment. Plant Dis. 2019, 103, 846–852. [Google Scholar] [CrossRef]
- Gimsing, A.L.; Strobel, B.W.; Hansen, H.C.B. Degradation and sorption of 2-propenyl and benzyl isothiocyanate in soil. Environ. Toxicol. Chem. 2009, 28, 1178–1184. [Google Scholar] [CrossRef]
- Weil, R.R.; Magdoff, F. Significance of soil organic matter to soil quality and health. In Soil Organic Matter in Sustainable Agriculture; Magdoff, F., Weil, R.R., Eds.; CRC Press: Boca Raton, FL, USA, 2004; p. 10. [Google Scholar]
- Doheny-Adams, T.; Lilley, C.J.; Barker, A.; Ellis, S.; Wade, R.; Atkinson, H.J.; Urwin, P.E.; Redeker, K.; Hartley, S.E. Constant isothiocyanate-release potentials across biofumigant seeding rates. J. Agric. Food Chem. 2018, 66, 5108–5116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngala, B.M.; Woods, S.R.; Back, M.A. In vitro assessment of the effects of Brassica juncea and Raphanus sativus leaf and root extracts on the viability of Globodera pallida encysted eggs. Nematology 2015, 17, 543–556. [Google Scholar] [CrossRef]
- Björkman, M.; Klingen, I.; Birch, A.N.E.; Bones, A.M.; Bruce, T.J.A.; Johansen, T.J.; Meadow, R.; Mølmann, J.; Seljåsen, R.; Smart, L.E. Phytochemicals of Brassicaceae in plant protection and human health–Influences of climate, environment and agronomic practice. Phytochemistry 2011, 72, 538–556. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.D.; Morra, M.J.; McCaffrey, J.P.; Auld, D.L.; Williams, L. Allelochemicals produced during glucosinolate degradation in soil. J. Chem. Ecol. 1991, 17, 2021–2034. [Google Scholar] [CrossRef]
- Fan, C.M.; Xiong, G.R.; Qi, P.; Ji, G.H.; He, Y.Q. Potential biofumigation effects of Brassica oleracea var. caulorapa on growth of fungi. J. Phytopathol. 2008, 156, 321–325. [Google Scholar] [CrossRef]
- El Nagdi, W.M.; Youssef, M.M.A. Brassica vegetable leaf residues as promising biofumigants for the control of root knot nematode, Meloidogyne incognita infecting cowpea. Agric. Eng. Int. CIGR J. 2019, 21, 134–139. [Google Scholar]
- Vandicke, J.; De Visschere, K.; Deconinck, S.; Leenknecht, D.; Vermeir, P.; Audenaert, K.; Haesaert, G. Uncovering the biofumigant capacity of allyl isothiocyanate from several Brassicaceae crops against Fusarium pathogens in maize. J. Sci. Food Agric. 2020, 100, 5476–5486. [Google Scholar] [CrossRef]
- Cheng, D.L.; Hashimoto, K.; Uda, Y. In vitro digestion of sinigrin and glucotropaeolin by single strains of Bifidobacterium and identification of the digestive products. Food Chem. Toxicol. 2004, 42, 351–357. [Google Scholar] [CrossRef]
- Rakariyatham, N.; Butrindr, B.; Niamsup, H.; Shank, L. Screening of filamentous fungi for production of myrosinase. Braz. J. Microbiol. 2005, 36, 242–245. [Google Scholar] [CrossRef] [Green Version]
- Ngala, B.M.; Woods, S.R.; Back, M.A. Sinigrin degradation and G. pallida suppression in soil cultivated with brassicas under controlled environmental conditions. App. Soil Ecol. 2015, 95, 9–14. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brassica Species | Common Name | Cultivar | Seed Rate | Supplier |
|---|---|---|---|---|
| Brassica juncea | Indian mustard | Brons | 8 kg ha−1 | RAGT Seeds UK |
| Brassica carinata | Ethiopian mustard | Cappucchino | 15 kg ha−1 | RAGT Seeds UK |
| Raphanus sativus | Oilseed radish | Bokito | 20 kg ha−1 | RAGT Seeds UK |
| Raphanus sativus | Oilseed radish | Terranova | 20 kg ha−1 | RAGT Seeds UK |
| Eruca sativa | Rocket | Trio | 10 kg ha−1 | RAGT Seeds UK |
| Brassica oleracea var. caulorapa L. | Kohlrabi | Kolibri | 0.5 kg ha−1 | Elsoms, UK |
| Brassica | Biomass (t ha−1) | |||||
|---|---|---|---|---|---|---|
| Fresh Weight | Dry Weight | |||||
| Shoot | Root | Total | Shoot | Root | Total | |
| Brassica juncea ‘Brons’ | 48.4 (5.6) a | 4.8 (0.8) | 53.3 (6.2) | 3.9 (0.7) | 0.7 (0.2) | 4.6 (0.8) |
| Brassica carinata ‘Cappucchino’ | 69.5 (16.5) | 4.3 (0.7) | 73.8 (17) | 6.4 (1.6) | 0.9 (0.2) | 7.3 (1.7) |
| Raphanus sativus ‘Bokito’ | 79.0 (14.7) | 10.4 (2.4) | 89.4 (15.3) | 6.3 (1.3) | 1.1 (0.2) | 7.4 (1.3) |
| Raphanus sativus ‘Terranova’ | 74.0 (22) | 10.5 (3.1) | 84.6 (23) | 5.6 (1.8) | 1.0 (0.3) | 6.5 (2.0) |
| Eruca sativa ‘Trio’ | 42.8 (14.4) | 2.4 (0.7) | 45.3 (15.0) | 3.6 (1.1) | 0.4 (0.1) | 4.0 (1.2) |
| Brassica oleracea var. caulorapa L. ‘Kolibri’ | 19.8 (8.0) | 0.9 (0.2) | 20.7 (8.2) | 2.6 (1.0) | 0.2 (0.1) | 2.9 (1.1) |
| Glucosinolate (µmol g−1 freeze-dried tissue) | B. juncea ‘Brons’ | B. carinata ‘Cappucchino’ | R. sativus ‘Bokito’ | R. sativus ‘Terranova’ | E. sativa ‘Trio’ | B. oleracea var. caulorapa L. ‘Kolibri’ | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | Shoot | Root | |
| Glucoberin | 2.25 (0.32) a | 2.17 (0.38) | 2.01 (0.63) | 1.89 (0.39) | 2.33 (0.74) | 2.22 (0.53) | 2.35 (0.41) | 2.25 (0.75) | 2.06 (0.58) | 2.18 (0.49) | 2.24 (0.21) | 1.83 (0.50) |
| Progoitrin | 0.42 (0.13) | 0.37 (0.08) | 0.34 (0.23) | 0.25 (0.17) | 0.42 (0.31) | 0.54 (0.39) | 0.33 (0.22) | 0.34 (0.27) | 2.08 (1.49) | 1.65 (1.02) | 0.39 (0.27) | 0.18 (0.13) |
| Sinigrin | 26.39 (12.42) | 19.97 (12.33) | 13.06 (6.21) | 4.81 (3.24) | - | - | - | - | - | - | - | - |
| Gluconapin | 0.20 (0.14) | 0.14 (0.17) | - | - | - | - | - | - | - | - | - | - |
| Glucobrassicin | 0.17 (0.07) | 0.32 (0.04) | 0.65 (0.30) | 0.43 (0.31) | 1.43 (0.79) | 1.54 (1.48) | 2.05 (1.29) | 0.78 (0.67) | 0.38 (0.38) | 0.05 (0.07) | 1.19 (0.76) | 0.91 (0.19) |
| Gluconasturtiin | 1.32 (0.70) | 18.04 (3.76) | 0.42 (0.29) | 6.17 (3.50) | 1.12 (0.49) | 2.47 (1.74) | 1.28 (0.61) | 2.38 (2.22) | 0.90 (0.42) | 2.96 (2.18) | 0.52 (0.37) | 5.42 (1.75) |
| Neoglucobrassicin | 0.07 (0.03) | 0.90 (0.17) | 0.11 (0.07) | 0.61 (0.44) | - | 0.42 (0.38) | 0.02 (0.03) | 0.12 (0.13) | 0.03 (0.02) | 0.00 | 0.53 (0.36) | 4.35 (1.82) |
| Glucoraphanin | - | - | - | - | 7.84 (4.79) | 1.39 (0.95) | 5.24 (4.43) | 1.47 (1.11) | 7.36 (3.96) | 3.94 (2.07) | 3.45 (2.72) | 1.30 (0.67) |
| Glucoraphenin | - | - | - | - | 0.45 (0.11) | - | 0.35 (0.19) | - | - | - | - | - |
| 4 hydroxy glucobrassicin | - | 0.04 (0.05) | - | - | - | - | - | 0.02 (0.03) | - | - | - | 0.27 (0.12) |
| Glucoraphasatin | - | - | - | - | 2.43 (1.69) | 14.90 (9.77) | 3.45 (2.60) | 17.60 (10.92) | - | - | - | - |
| Glucoalyssin | - | - | - | - | - | - | - | - | 0.50 (0.21) | 0.24 (0.21) | - | - |
| Glucoerucin | - | - | - | - | - | - | - | - | 7.03 (3.65) | 12.28 (7.30) | 0.14 (0.16) | 3.80 (1.71) |
| 4-mercaptobutyl | - | - | - | - | - | - | - | - | 4.89 (2.04) | 1.26 (1.94) | - | - |
| unknown | - | - | - | - | - | - | - | - | 0.53 (0.37) | - | - | - |
| unknown | - | - | - | - | - | - | - | - | 0.93 (0.70) | - | - | - |
| Total glucosinolates | 30.83 (13.76) | 41.91 (15.63) | 16.59 (7.62) | 14.19 (7.31) | 16.02 (8.66) | 23.47 (14.65) | 15.06 (7.33) | 24.96 (15.10) | 26.69 (12.33) | 24.57 (12.69) | 8.46 (4.55) | 18.06 (6.22) |
| Treatments | % Incidence Fusarium graminearum Recovered in Oat Inoculum | |
|---|---|---|
| 8 Weeks Post-Incorporation | 16 Weeks Post-Incorporation | |
| Fallow | 95 | 98 |
| Fallow uncultivated | 95 | 97 |
| Brassica juncea ‘Brons’ | 98 | 97 |
| Brassica carinata ‘Cappucchino’ | 96 | 95 |
| Raphanus sativus ‘Bokito’ | 96 | 94 |
| Raphanus sativus ‘Terranova’ | 95 | 95 |
| Eruca sativa ‘Trio’ | 94 | 94 |
| Brassica oleracea var. caulorapa L. ‘Kolibri’ | 97 | 97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashiq, S.; Edwards, S.; Watson, A.; Back, M. Biofumigation for the Management of Fusarium graminearum in a Wheat-Maize Rotation. Pathogens 2022, 11, 1427. https://doi.org/10.3390/pathogens11121427
Ashiq S, Edwards S, Watson A, Back M. Biofumigation for the Management of Fusarium graminearum in a Wheat-Maize Rotation. Pathogens. 2022; 11(12):1427. https://doi.org/10.3390/pathogens11121427
Chicago/Turabian StyleAshiq, Samina, Simon Edwards, Andrew Watson, and Matthew Back. 2022. "Biofumigation for the Management of Fusarium graminearum in a Wheat-Maize Rotation" Pathogens 11, no. 12: 1427. https://doi.org/10.3390/pathogens11121427