
Outbreak of Parasitic Dinoflagellate Piscinoodinium sp. Infection in an Endangered Fish from India: Arulius Barb (Dawkinsia arulius)

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Case History
2.2. Examination of Affected Fish and Collection of Parasites
2.3. Polymerase Chain Reaction and Sequencing of rDNA
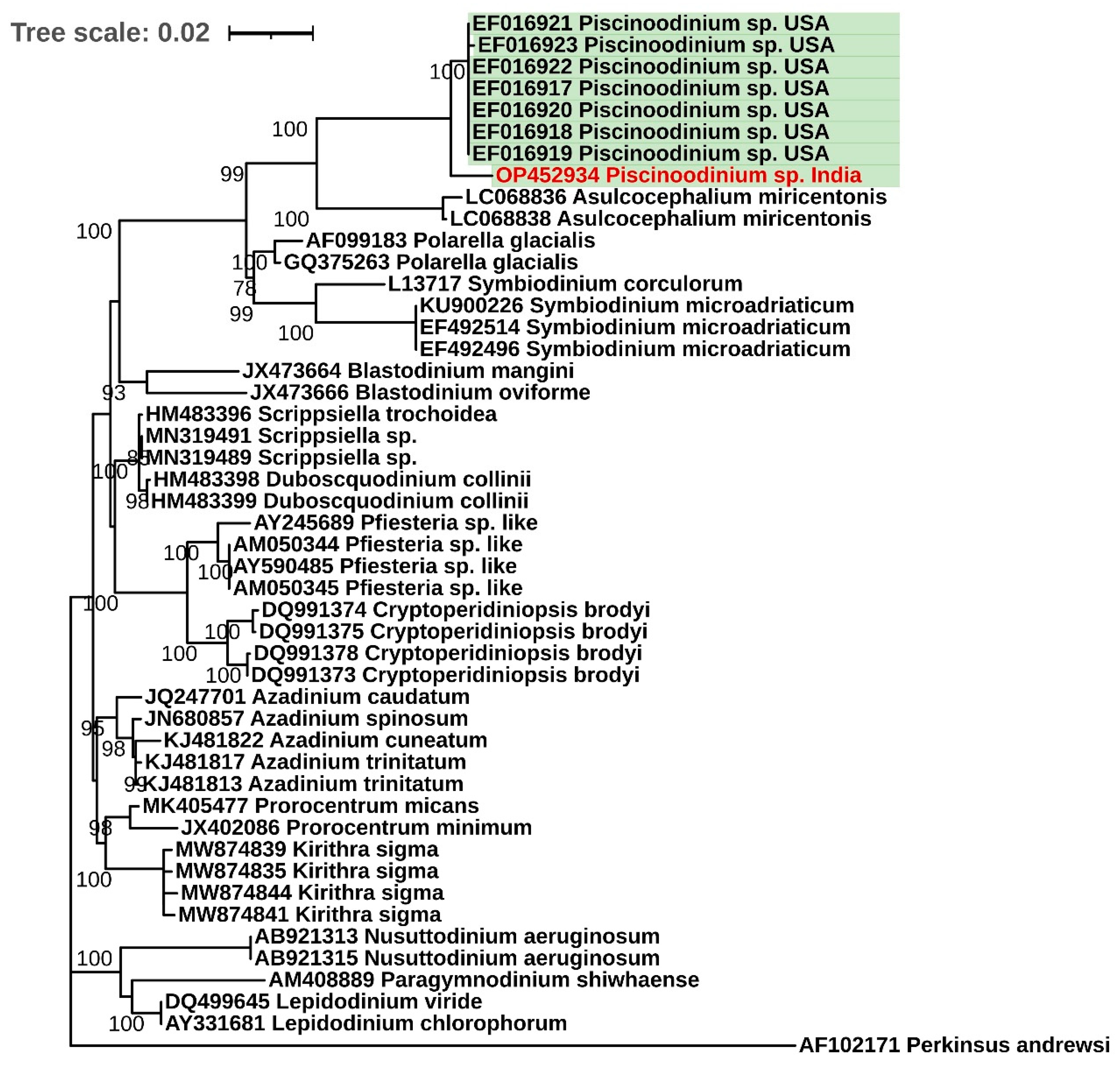
2.4. Phylogenetic Analysis
3. Results
3.1. Clinical Signs and Parasitological Examination
3.2. Polymerase Chain Reaction and Sequence Analysis
3.3. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noga, E.J. Fish Disease; Blackwell Publishing, Inc.: Ames, IA, USA, 2010; ISBN 9781118786758. [Google Scholar]
- Martins, M.L.; Moraes, J.R.; Andrade, P.M.; Schalch, S.H.; Moraes, F.R. Piscinoodinium pillulare (Schäperclaus, 1954) Lom, 1981 (Dinoflagellida) Infection in Cultivated Freshwater Fish from the Northeast Region of Sao Paulo State, Brazil. Parasitological and Pathological Aspects. Braz J. Biol. 2001, 61, 639–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zago, A.C.; Franceschini, L.; Garcia, F.; Schalch, S.H.C.; Gozi, K.S.; Silva, R.J. da Ectoparasites of Nile Tilapia (Oreochromis Niloticus) in Cage Farming in a Hydroelectric Reservoir in Brazil. Rev. Bras. Parasitol. Vet. 2014, 23, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Neves, L.R.; Pereira, F.B.; Tavares-Dias, M.; Luque, J.L. Seasonal Influence on the Parasite Fauna of a Wild Population of Astronotus ocellatus (Perciformes: Cichlidae) from the Brazilian Amazon. J. Parasitol. 2013, 99, 718–721. [Google Scholar] [CrossRef] [PubMed]
- Shaharom-Harrison, F.M.; Anderson, I.G.; Siti, A.Z.; Shazili, N.A.M.; Ang, K.J.; Azmi, T.I. Epizootics of Malaysian Cultured Freshwater Pond Fishes by Piscinoodinium pillulare (Schaperclaus 1954) Lom 1981. Aquaculture 1990, 86, 127–138. [Google Scholar] [CrossRef]
- Wilson, J.R.; Saunders, R.J.; Hutson, K.S. Parasites of the Invasive Tilapia Oreochromis mossambicus: Evidence for Co-Introduction. Aquat. Invasions 2019, 14, 332–349. [Google Scholar] [CrossRef]
- Noga, E.J.; Levy, M.G. Phylum Dinoflagellata. In Fish Diseases and Disorders. Volume 1: Protozoan and Metazoan Infections; CABI: Wallingford, UK, 2006; pp. 16–45. [Google Scholar]
- Ramesh, K.S.; Mohan, C.V.; Shankar, K.M.; Ahmed, I. Piscinoodinium Sp. Infection in Juveniles of Common Carp (Cyprinus Carpio), Mahseer (Tor Khudree) and Tilapia (Oreochromis mossambicus). J. Aquac. Trop. 2002, 15, 281–288. [Google Scholar]
- Ferraz, E.; Sommerville, C. Pathology of Piscinoodinium Sp. (Protozoa: Dinoflagellida), Parasites of the Ornamental Freshwater Catfishes Corydoras spp. and Brochis splendens (Pisces: Callichthyidae). Dis. Aquat. Organ. 1998, 33, 43–49. [Google Scholar] [CrossRef]
- van Duijn, C. Diseases of Fish; Thomas H. Charles: Springfield, MA, USA, 1973. [Google Scholar]
- Jacobs, D.L. A New Parasitic Dinoflagellate from Fresh-Water Fish. Trans. Am. Microsc. Soc. 1946, 65, 1–17. [Google Scholar] [CrossRef]
- Levy, M.G.; Litaker, R.W.; Goldstein, R.J.; Dykstra, M.J.; Vandersea, M.W.; Noga, E.J. Piscinoodinium, a Fish-Ectoparasitic Dinoflagellate, Is a Member of the Class Dinophyceae, Subclass Gymnodiniphycidae: Convergent Evolution with Amyloodinium. J. Parasitol. 2007, 93, 1006–1015. [Google Scholar] [CrossRef]
- Abraham, R. Dawkinsia Arulius; The IUCN Red List of Threatened Species 2015: e.T172500A70168511. Available online: https://www.iucnredlist.org/species/172500/70168511 (accessed on 14 July 2022).
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A Simple Salting out Procedure for Extracting DNA from Human Nucleated Cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [Green Version]
- Jeffery, K.R.; Bateman, K.; Bayley, A.; Feist, S.W.; Hulland, J.; Longshaw, C.; Stone, D.; Woolford, G.; Way, K. Isolation of a Cyprinid Herpesvirus 2 from Goldfish, Carassius auratus (L.), in the UK. J. Fish. Dis. 2007, 30, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Oyamatsu, T.; Matoyama, H.; Yamamoto, K.-Y.; Fukuda, H. A Trial for the Detection of Carp Edema Virus by Using Polymerase Chain Reaction. Aquac. Sci. 1997, 45, 247–251. [Google Scholar] [CrossRef]
- Swaminathan, T.R.; Raj, N.S.; Preena, P.G.; Pradhan, P.K.; Sood, N.; Kumar, R.G.; Sudhagar, A.; Sood, N.K. Infectious Spleen and Kidney Necrosis Virus-associated Large-scale Mortality in Farmed Giant Gourami, Osphronemus Goramy, in India. J. Fish. Dis. 2021, 44, 2043–2053. [Google Scholar] [CrossRef]
- Nishizawa, T.; Mori, I.K.; Nakai, T.; Furusawa, I.; Muroga, K. Polymerase Chain Reaction (PCR) Amplification of RNA of Striped Jack Nervous Necrosis Virus (SJNNV). Dis. Aquat. Organ. 1994, 18, 103–107. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (ITOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Murray, A.G.; Peeler, E.J. A Framework for Understanding the Potential for Emerging Diseases in Aquaculture. Prev. Vet. Med. 2005, 67, 223–235. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Harvell, C.D.; Conrad, J.M.; Friedman, C.S.; Kent, M.L.; Kuris, A.M.; Powell, E.N.; Rondeau, D.; Saksida, S.M. Infectious Diseases Affect Marine Fisheries and Aquaculture Economics. Ann. Rev. Mar. Sci. 2015, 7, 471–496. [Google Scholar] [CrossRef] [Green Version]
- Russell, R.E.; DiRenzo, G.V.; Szymanski, J.A.; Alger, K.E.; Grant, E.H.C. Principles and Mechanisms of Wildlife Population Persistence in the Face of Disease. Front. Ecol. Evol. 2020, 8, 569016. [Google Scholar] [CrossRef]
- Bailey, C.; Segner, H.; Casanova-Nakayama, A.; Wahli, T. Who Needs the Hotspot? The Effect of Temperature on the Fish Host Immune Response to Tetracapsuloides bryosalmonae the Causative Agent of Proliferative Kidney Disease. Fish Shellfish. Immunol. 2017, 63, 424–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, C.; Schmidt-Posthaus, H.; Segner, H.; Wahli, T.; Strepparava, N. Are Brown Trout Salmo trutta fario and Rainbow Trout Oncorhynchus mykiss Two of a Kind? A Comparative Study of Salmonids to Temperature-Influenced Tetracapsuloides bryosalmonae Infection. J. Fish. Dis. 2018, 41, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Ott, B.M.; Litaker, R.W.; Holland, W.C.; Delwiche, C.F. Using RDNA Sequences to Define Dinoflagellate Species. PLoS ONE 2022, 17, e0264143. [Google Scholar] [CrossRef] [PubMed]
- Shivam, S.; El-Matbouli, M.; Kumar, G. Development of Fish Parasite Vaccines in the OMICs Era: Progress and Opportunities. Vaccines 2021, 9, 179. [Google Scholar] [CrossRef]
- Fariya, N.; Kaur, H.; Singh, M.; Abidi, R.; El-Matbouli, M.; Kumar, G. Morphological and Molecular Characterization of a New Myxozoan, Myxobolus grassi sp. Nov. (Myxosporea), Infecting the Grass Carp, Ctenopharyngodon idella in the Gomti River, India. Pathogens 2022, 11, 303. [Google Scholar] [CrossRef]
- Sreenivasan, N.; Mahesh, N.; Raghavan, R. Freshwater Fishes of Cauvery Wildlife Sanctuary, Western Ghats of Karnataka, India. J. Threat. Taxa 2021, 13, 17470–17476. [Google Scholar] [CrossRef]
- Sudhagar, A.; Kumar, G.; El-Matbouli, M. The Malacosporean Myxozoan Parasite Tetracapsuloides bryosalmonae: A Threat to Wild Salmonids. Pathogens 2019, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Sudhagar, A.; Kumar, G.; El-Matbouli, M. Transcriptome Analysis Based on RNA-Seq in Understanding Pathogenic Mechanisms of Diseases and the Immune System of Fish: A Comprehensive Review. Int. J. Mol. Sci. 2018, 19, 245. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Primer ID | Primer Sequence (5′-3′) | Annealing Temperature (°C) | Target Size (bp) | Purpose | Reference |
|---|---|---|---|---|---|
| GCG18SF | CTGGTGATCCTGCCAGTAGTC | 58 | 2334 | Piscinoodinium sp. PCR | [12] |
| ITSR8 | TAACCTGCATTCATGCTGTG | ||||
| INT_F | GTCTGGTGCCAGCAGCCGCGG | 58 | - | Sequencing | Present study |
| INT_R | GTACAAAGGGCAGGGACGTA | 58 | - | Sequencing | Present study |
| T7 | TAATACGACTCACTATAGGG | 55 | - | Sequencing | Universal primer |
| SP6 | ATTTAGGTGACACTATAG | 55 | - | Sequencing | Universal primer |
| CyHV_F | CCCAGCAACATGTGCGACGG | 55 | 362 | CyHV-2 PCR | [15] |
| CyHV_R | CCGTARTGAGAGTTGGCGCA | ||||
| CEV_F1 | GCTGTTGCAACCATTTGAGA | 60 | 548 | CEV nested PCR (outer) | [16] |
| CEV_R1 | TGCAGGTTGCTCCTAATCCT | ||||
| CEV_F2 | GCTGCTGCACTTTTAGGAGG | 55 | 181 | CEV nested PCR (inner) | |
| CEV_R1 | TGCAAGTTATTTCGATGCCA | ||||
| ISKNVF | ATGTCTGCAATCTCAGGTGC | 55 | 1362 | ISKNV PCR | [17] |
| ISKNVR | TTACAGGATAGGGAAGCCTG | ||||
| VNN_F2 | CGTGTCAGTCATGTGTCGCT | 55 | 430 | VNN PCR | [18] |
| VNN_R3 | CGAGTCAACACGGGTGAAGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sudhagar, A.; Sundar Raj, N.; Mohandas, S.P.; Serin, S.; Sibi, K.K.; Sanil, N.K.; Raja Swaminathan, T. Outbreak of Parasitic Dinoflagellate Piscinoodinium sp. Infection in an Endangered Fish from India: Arulius Barb (Dawkinsia arulius). Pathogens 2022, 11, 1350. https://doi.org/10.3390/pathogens11111350
Sudhagar A, Sundar Raj N, Mohandas SP, Serin S, Sibi KK, Sanil NK, Raja Swaminathan T. Outbreak of Parasitic Dinoflagellate Piscinoodinium sp. Infection in an Endangered Fish from India: Arulius Barb (Dawkinsia arulius). Pathogens. 2022; 11(11):1350. https://doi.org/10.3390/pathogens11111350
Chicago/Turabian StyleSudhagar, Arun, Nithianantham Sundar Raj, Sowmya Pazhur Mohandas, Shaji Serin, Konnoth Kuttappan Sibi, Nandiath Karayi Sanil, and Thangaraj Raja Swaminathan. 2022. "Outbreak of Parasitic Dinoflagellate Piscinoodinium sp. Infection in an Endangered Fish from India: Arulius Barb (Dawkinsia arulius)" Pathogens 11, no. 11: 1350. https://doi.org/10.3390/pathogens11111350