
In Silico Evaluation of the Haplotype Diversity, Phylogenetic Variation and Population Structure of Human E. granulosus sensu stricto (G1 Genotype) Sequences

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.2. Alignment and Phylogenetic Analysis
2.3. Haplotype Analysis and Networking
3. Results
3.1. Polymorphism and Haplotype Analysis
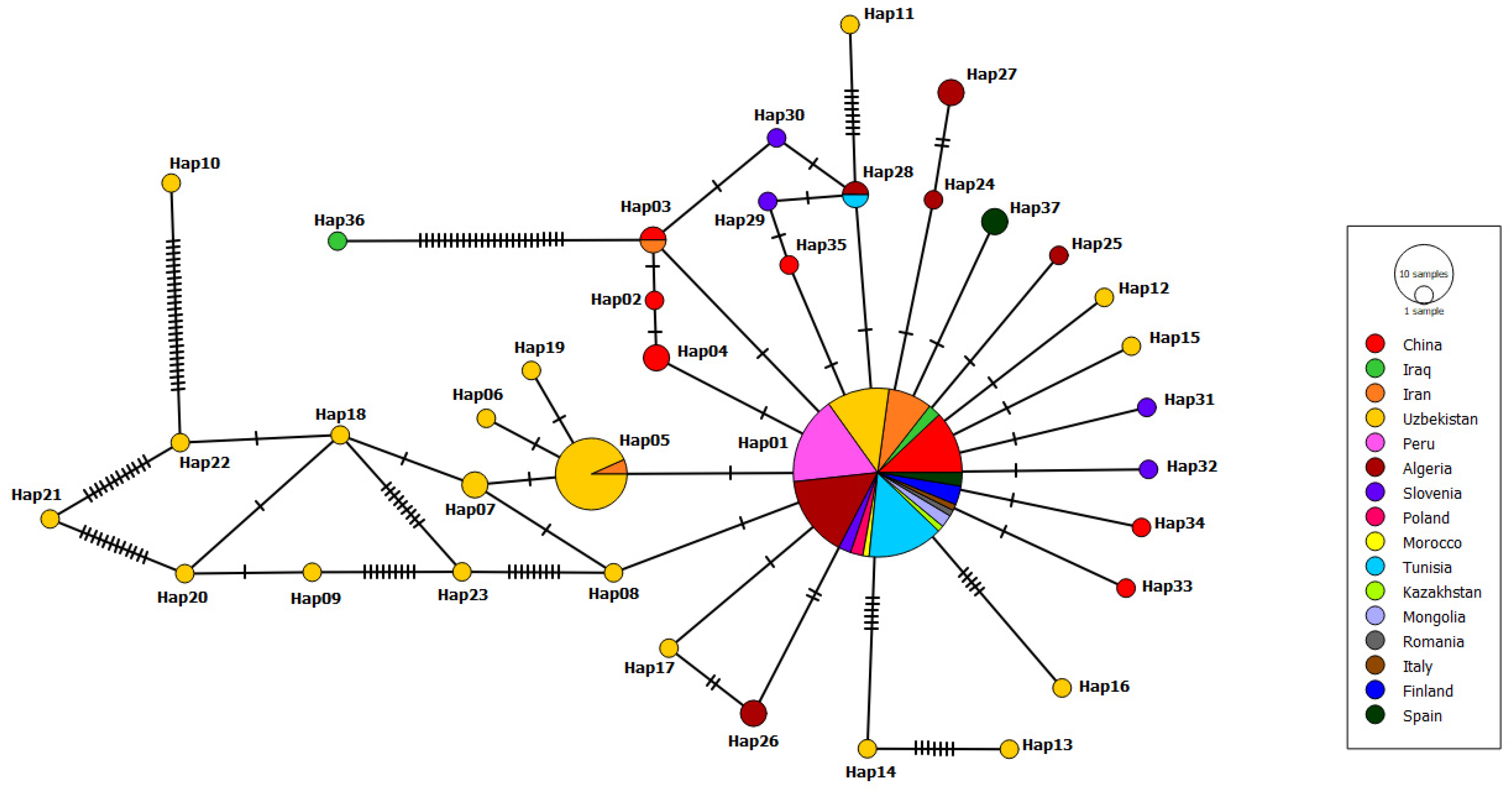
3.2. Haplotype Network
3.3. Phylogenetic Tree
3.4. Gene Flow, Diversity and Neutrality Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McManus, D.P.; Thompson, R.C.A. Molecular epidemiology of cystic echinococcosis. Parasitology 2003, 127, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Batsch, A.J.G.K. Naturgeschichte Der Bandwurmgattung Überhaupt und Ihrer Arten Insbesondere; Bey Johann Jacob Gebauer: Halle, Germany, 1786. [Google Scholar]
- Moro, P.; Schantz, P.M. Cystic echinococcosis in the Americas. Parasitol. Int. 2006, 55, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Deplazes, P.; Rinaldi, L.; Rojas, C.A.; Torgerson, P.R.; Harandi, M.F.; Romig, T.; Antolova, D.; Schurer, J.M.; Lahmar, S.; Cringoli, G.; et al. Global distribution of alveolar and cystic echinococcosis. Adv. Parasitol. 2017, 95, 315–493. [Google Scholar] [PubMed] [Green Version]
- World Health Organization. Multicriteria-Based Ranking for Risk Management of Food-Borne Parasites: Report of a Joint FAO; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- World Health Organization. Sustaining the Drive to Overcome the Global Impact of Neglected Tropical Diseases: Second WHO Report on Neglected Diseases; WHO: Geneva, Switzerland, 2013. [Google Scholar]
- Vuitton, D.A.; McManus, D.P.; Rogan, M.T.; Romig, T.; Gottstein, B.; Naidich, A.; Tuxun, T.; Wen, H.; da Silva, A.M. International consensus on terminology to be used in the field of echinococcoses. Parasite 2020, 27, 41. [Google Scholar] [CrossRef]
- Nakao, M.; McManus, D.P.; Schantz, P.M.; Craig, P.S.; Ito, A. A molecular phylogeny of the genus Echinococcus inferred from complete mitochondrial genomes. Parasitology 2006, 134, 713–722. [Google Scholar] [CrossRef] [Green Version]
- Busi, M.; Šnábel, V.; Varcasia, A.; Garippa, G.; Perrone, V.; De Liberato, C.; D’Amelio, S. Genetic variation within and between G1 and G3 genotypes of Echinococcus granulosus in Italy revealed by multilocus DNA sequencing. Vet. Parasitol. 2007, 150, 75–83. [Google Scholar] [CrossRef]
- Rojas, C.A.A.; Romig, T.; Lightowlers, M.W. Echinococcus granulosus sensu lato genotypes infecting humans–review of current knowledge. Int. J. Parasitol. 2014, 44, 9–18. [Google Scholar] [CrossRef]
- Lymbery, A.J. Phylogenetic pattern, evolutionary processes and species delimitation in the genus Echinococcus. Adv. Parasitol. 2017, 95, 111–145. [Google Scholar]
- Jenkins, D.J.; Romig, T.; Thompson, R.C.A. Emergence/re-emergence of Echinococcus spp.—A global update. Int. J. Parasitol. 2005, 35, 1205–1219. [Google Scholar] [CrossRef]
- Lewall, D.B. Hydatid disease: Biology, pathology, imaging and classification. Clin. Radiol. 1998, 53, 863–874. [Google Scholar] [CrossRef]
- Thompson, R.C.A. The taxonomy, phylogeny and transmission of Echinococcus. Exp. Parasitol. 2008, 119, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Patrice Bourée, M.D. Hydatidosis: Dynamics of transmission. World J. Surg. 2001, 25, 429. [Google Scholar]
- Kammerer, W.S.; Schantz, P.M. Echinococcal disease. Infect. Dis. Clin. N. Am. 1993, 7, 605–618. [Google Scholar] [CrossRef]
- Moro, P.; Schantz, P.M. Echinococcosis: A review. Int. J. Infect. Dis. 2009, 13, 125–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moro, P.L.; Lopera, L.; Cabrera, M.; Cabrera, G.; Silva, B.; Gilman, R.H.; Moro, M.H. Endemic focus of cystic echinococcosis in a coastal city of Peru. Am. J. Trop. Med. Hyg. 2004, 71, 327–329. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G.; Gruttadauria, S.; Biondi, A.; Marventano, S.; Mistretta, A. Worldwide epidemiology of liver hydatidosis including the Mediterranean area. World J. Gastroenterol. 2012, 18, 1425. [Google Scholar] [CrossRef]
- Haag, K.L.; Ayala, F.J.; Kamenetzky, L.; Gutierrez, A.M.; Rosenzvit, M. Livestock trade history, geography, and parasite strains: The mitochondrial genetic structure of Echinococcus granulosus in Argentina. J. Parasitol. 2004, 90, 234–239. [Google Scholar] [CrossRef]
- Shimshony, A. Epidemiology of emerging zoonoses in Israel. Emerg. Infect. Dis. 1997, 3, 229. [Google Scholar] [CrossRef]
- Bart, J.M.; Abdukader, M.; Zhang, Y.L.; Lin, R.Y.; Wang, Y.H.; Nakao, M.; Ito, A.; Craig, P.S.; Piarroux, R.; Vuitton, D.A.; et al. Genotyping of human cystic echinococcosis in Xinjiang, PR China. Parasitology 2006, 133, 571–579. [Google Scholar] [CrossRef]
- Buishi, I.E.; Njoroge, E.M.; Bouamra, O.; Craig, P.S. Canine echinococcosis in northwest Libya: Assessment of coproantigen ELISA, and a survey of infection with analysis of risk-factors. Vet. Parasitol. 2005, 130, 223–232. [Google Scholar] [CrossRef]
- Ahmed, E.S.; Helmy, H.; El Setouhy, M.; Craig, P.S.; Ramzy, R.M.R. A retrospective hospital study of human cystic echinococcosis in Egypt. East. Mediterr. Health J. 2004, 10, 349–357. [Google Scholar]
- Sadjjadi, S.M. Present situation of echinococcosis in the Middle East and Arabic North Africa. Parasitol. Int. 2006, 55, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Bardonnet, K.; Benchikh-Elfegoun, M.C.; Bart, J.M.; Harraga, S.; Hannache, N.; Haddad, S.; Dumon, H.; Vuitton, D.A.; Piarroux, R. Cystic echinococcosis in Algeria: Cattle act as reservoirs of a sheep strain and may contribute to human contamination. Vet. Parasitol. 2003, 116, 35–44. [Google Scholar] [CrossRef]
- Mwambete, K.D.; Ponce-Gordo, F.; Cuesta-Bandera, C. Genetic identification and host range of the Spanish strains of Echinococcus granulosus. Acta Trop. 2004, 91, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Eckert, J.; Thompson, R.C. Echinococcus strains in Europe: A review. Ann. Trop. Med. Parasitol. 1998, 39, 1–8. [Google Scholar]
- Garippa, G. Updates on cystic echinococcosis (CE) in Italy. Parassitologia 2006, 48, 57. [Google Scholar]
- Azlaf, R.; Dakkak, A. Epidemiological study of the cystic echinococcosis in Morocco. Vet. Parasitol. 2006, 137, 83–93. [Google Scholar] [CrossRef]
- La Rue, M.L.D.; Dinkel, A.; Mackenstedt, U.; Romig, T. New data on Echinococcus spp. in southern Brazil. Rev. Inst. Med. Trop. Sao Paulo. 2006, 48, 103–104. [Google Scholar] [CrossRef] [Green Version]
- Thompson, R.C.A.; McManus, D. Aetiology: Parasites and life-cyles. In WHO/OIE Manual on Echinococcosis in Humans and Animals: A Public Health Problem of Global Concern; Eckert, J., Gemmell, M.A., Meslin, F.-X., Pawlowski, Z.S., Eds.; World Organisation for Animal Health: Paris, France, 2001; pp. 1–19. [Google Scholar]
- Eckert, J.; Gemmell, M.A.; Meslin, F.X.; Pawlowski, Z.S. WHO/OIE Manual on Echinococcosis in Humans and Animals: A Public Health Problem of Global Concern; World Organisation for Animal Health: Paris, France, 2001. [Google Scholar]
- McManus, D.P. The molecular epidemiology of Echinococcus granulosus and cystic hydatid disease. Trans. R. Soc. Trop. Med. Hyg. 2002, 96, 151–157. [Google Scholar] [CrossRef]
- Jia, W.; Yan, H.; Lou, Z.; Ni, X.; Dyachenko, V.; Li, H.; Littlewood, D.T.J. Mitochondrial genes and genomes support a cryptic species of tapeworm within Taenia taeniaeformis. Acta Trop. 2012, 123, 154–163. [Google Scholar] [CrossRef]
- Bowles, J.; McManus, D.P. NADH dehydrogenase 1 gene sequences compared for species and strains of the genus Echinococcus. Int. J. Parasitol. 1993, 23, 969–972. [Google Scholar] [CrossRef]
- Bowles, J.; Blair, D.; McManus, D.P. Genetic variants within the genus Echinococcus identified by mitochondrial DNA sequencing. Mol. Biochem. Parasitol. 1992, 54, 165–173. [Google Scholar] [CrossRef]
- Bowles, J.; Blair, D.; McManus, D.P. Molecular genetic characterization of the cervid strain (‘northern form’) of Echinococcus granulosus. Parasitology 1994, 109, 215–221. [Google Scholar] [CrossRef]
- Knudsen, B.; Knudsen, T.; Flensborg, M.; Sandmann, H.; Heltzen, M.; Andersen, A.; Sinding, J.B. CLC Main Workbench, Version 5.5; CLC bio: Aarhus, Denmark, 2007. [Google Scholar]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Maddison, D.R.; Swofford, D.L.; Maddison, W.P. NEXUS: An extensible file format for systematic information. Syst. Biol. 1997, 46, 590–621. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Tünger, Ö. Dünyada Kistik Ekinokokkoz Epidemiyolojisi. Turkiye Parazitol. Derg. 2013, 37, 47–52. [Google Scholar]
- Eckert, J.; Deplazes, P. Biological, epidemiological, and clinical aspects of echinococcosis, a zoonosis of increasing concern. Clin. Microbiol. Rev. 2004, 17, 107–135. [Google Scholar] [CrossRef] [Green Version]
- Unat, E.K.; Yücel, A.; Atlas, K.; Samastı, M. Unat’ın Tıp Parazitolojisi. Cerrahpasa Med. J. 1995, 15, 440–459. [Google Scholar]
- Kinkar, L.; Laurimäe, T.; Acosta-Jamett, G.; Andresiuk, V.; Balkaya, I.; Casulli, A.; Gasser, R.B.; van der Giessen, J.; González, L.M.; Haag, K.L.; et al. Global phylogeography and genetic diversity of the zoonotic tapeworm Echinococcus granulosus sensu stricto genotype G1. Int. J. Parasitol. 2018, 48, 729–742. [Google Scholar] [CrossRef]
- Ramos-Onsins, S.E.; Rozas, J. Statistical properties of new neutrality tests against population growth. Mol. Biol. Evol. 2002, 19, 2092–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vamathevan, J.J.; Hasan, S.; Emes, R.D.; Amrine-Madsen, H.; Rajagopalan, D.; Topp, S.D.; Kumar, V.; Word, M.; Simmons, M.D.; Foord, S.M.; et al. The role of positive selection in determining the molecular cause of species differences in disease. BMC Evol. Biol. 2008, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, J.C.; Schneider, J.A.; Tanguay, D.A.; Choi, J.; Acharya, T.; Stanley, S.E.; Jiang, R.; Messer, C.J.; Chew, A.; Han, J.H.; et al. Haplotype variation and linkage disequilibrium in 313 human genes. Science 2001, 293, 489–493. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.-X. Statistical tests of neutrality of mutations against population growth, hitchhiking and back ground selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.L.; Kong, X.Y.; Yu, Z.N.; Kong, J.; Ma, S.; Chen, L.M. Genetic diversity and historical demography of Chinese shrimp Feneropenaeus chinensis in Yellow Sea and Bohai Sea based on mitochondrial DNA analysis. Afr. J. Biotechnol. 2009, 8, 1193–1202. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mt-CO1 | mt-ND1 | ||||
|---|---|---|---|---|---|
| Origin | No. of Isolates | Accession Numbers | Origin | No. of Isolates | Accession Numbers |
| China | 43 | DQ356874-75-76-77-78-79-80/83, KJ628328-29-30-31-32-33-34-35, AB688602-03-04-05-06-07-08-09-10-11/13-14-15-16-17-18-19, MH050608-09-10-11-12-13-14-15-16-17 | Uzbekistan | 43 | MN696570/72/76-77-78-79-80-81-82-83-84-85-86-87-88-89-90-91-92-93-94-95-96-98-99, MN696600-01-02-03-04/06-07-08-09-10-11-12-13/15/19-20-21-22 |
| Tunisia | 13 | MG672264-65-66-67-68-69-70-71-72-73-74-75-76 | Algeria | 20 | MG672128, KT316342, KR349038-39-40-41-42-43-44, MG672282/84-85-86-87-88-89-90-91-92-93 |
| Pakistan | 12 | MK229295-96-97/99, MK229301-02/04/13/15/17-18-19 | China | 17 | AY572548, KJ556993-94, EU072111-12-13-14, MH050620-21-22-23-24-25-26-27-28-29 |
| Mongolia | 12 | MG672254-55, AB893242-43-44-45-46-47-48-49-50-51 | Peru | 14 | JF946597-98-99, JF946600-01-02-03-04-05-06-07-08-09/24, |
| Algeria | 12 | MG672128, MG672283-84-85-86-87-88-89-90-91-92-93 | Tunisia | 13 | MG672264-65-66-67-68-69-70-71-72-73-74-75-76 |
| Turkey | 10 | EU006783, GU951512-13, MG886833-34-35-36-37-38-39 | Iran | 9 | KT284349, MG672245, JF836800-01-02-03, JF836797-98-99 |
| Iran | 9 | KR337817, MW350099, MT073987, MG672245, MH025946-47, JQ250810/12/15 | Slovenia | 6 | MT239133-34-35/38/40/42 |
| Russia | 9 | AB777904/07-08, AB688136-37-38-39-40-41 | Spain | 4 | KU925413-14, MG672129/37 |
| Spain | 4 | MG672129/37, KU925413-14, | Iraq | 3 | FJ226756, MN231833-34 |
| Finland | 3 | MG672132, KY766884, KU925429 | Finland | 3 | MG672132, KY766884, KU925429 |
| India | 2 | JX854029-30 | Mongolia | 2 | MG672254-55 |
| Morocco | 1 | EF367266 | Poland | 2 | KT780298- KT780300 |
| Romania | 1 | MG672138 | Romania | 1 | MG672138 |
| Kazakhstan | 1 | MG672257 | Kazakhstan | 1 | MG672257 |
| Italy | 1 | MG672135 | Italy | 1 | MG672135 |
| Morocco | 1 | EF367298 | |||
| Haplotype Name | No. of Isolate | Accession Numbers |
|---|---|---|
| Hap01 | 2 | KJ628335-China, KJ628331-China |
| Hap02 | 1 | KR337817-Iran |
| Hap03 | 79 | GU951513-Turkey, GU951512-Turkey, JX854029-India, MG886839-Turkey, MG886838-Turkey, MG886837-Turkey, MG886836-Turkey, MG886835-Turkey, MG886834-Turkey, MG886833-Turkey, DQ356883-China, MW350099-Iran, MT073987-Iran, MK229301-Pakistan, MK229296-Pakistan, MH050617-China, MH050615-China, MH050614-China, MH050612-China, MH050609-China, MH050608-China, MH025946-Iran, KJ628334-China, KJ628333-China, KJ628332-China, KJ628330-China, KJ628329-China, AB688617-China, AB688616-China, AB688614-China, AB688611-China, AB688610-China, AB688609-China, AB688608-China, AB688607-China, AB688603-China, AB688602-China, JQ250815-Iran, AB893250-Mongolia, AB893249-Mongolia, AB893248-Mongolia, AB893247-Mongolia, AB893245-Mongolia, AB893244-Mongolia, AB893243-Mongolia, AB777908-Russia, AB777907-Russia, AB777904-Russia, AB688141-Russia, AB688136-Russia, MG672293-Algeria, MG672292-Algeria, MG672291-Algeria, MG672289-Algeria, MG672288-Algeria, MG672287-Algeria, MG672285-Algeria, MG672284-Algeria, MG672283-Algeria, MG672276-Tunisia, MG672275-Tunisia, MG672274-Tunisia, MG672272-Tunisia, MG672270-Tunisia, MG672269-Tunisia, MG672268-Tunisia, MG672266-Tunisia, MG672265-Tunisia, MG672264-Tunisia, MG672257-Kazakhstan, MG672254-Mongolia, MG672245-Iran, MG672135-Italy, MG672132-Finland, MG672129-Spain, MG672128-Algeria, KY766884-Finland, KU925429-Finland, KU925413-Spain |
| Hap04 | 1 | EU006783-Turkey |
| Hap05 | 2 | JX854030-India, MH050610-China |
| Hap06 | 2 | MK229304-Pakistan, MK229299-Pakistan |
| Hap07 | 1 | DQ356880-China |
| Hap08 | 1 | DQ356879-China |
| Hap09 | 2 | DQ356878-China, AB688604-China |
| Hap10 | 12 | DQ356877-China, MK229302-Pakistan, MK229297-Pakistan, MH050611-China, JQ250812-Iran, JQ250810-Iran, AB893246-Mongolia, AB688140-Russia, AB688139-Russia, MG672290-Algeria, MG672267-Tunisia, MG672255-Mongolia |
| Hap11 | 2 | DQ356876-China, DQ356875-China |
| Hap12 | 1 | DQ356874-China |
| Hap13 | 2 | MK229319-Pakistan, MK229313-Pakistan |
| Hap14 | 3 | MK229318-Pakistan, AB688618-China, MG672286-Algeria |
| Hap15 | 1 | MK229317-Pakistan |
| Hap16 | 1 | MK229315-Pakistan |
| Hap17 | 1 | MK229295-Pakistan |
| Hap18 | 2 | MH050616-China, AB688606-China |
| Hap19 | 1 | MH050613-China |
| Hap20 | 1 | EF367266-Morocco |
| Hap21 | 1 | MH025947-Iran |
| Hap22 | 1 | KJ628328-China |
| Hap23 | 1 | AB688619-China |
| Hap24 | 1 | AB688615-China |
| Hap25 | 1 | AB688613-China |
| Hap26 | 1 | AB688605-China |
| Hap27 | 1 | AB893251-Mongolia |
| Hap28 | 1 | AB893242-Mongolia |
| Hap29 | 1 | AB688138-Russia |
| Hap30 | 1 | AB688137-Russia |
| Hap31 | 1 | MG672273-Tunisia |
| Hap32 | 1 | MG672271-Tunisia |
| Hap33 | 1 | MG672138-Romania |
| Hap34 | 2 | MG672137-Spain, KU925414-Spain |
| Haplotype Name | No. of Isolate | Accession Numbers |
|---|---|---|
| Hap01 | 83 | KU925413-Spain, EU072111-China, FJ226756-Iraq, KT284349-Iran, JF836803-Iran, JF836802-Iran, JF836801-Iran, JF836799-Iran, JF836797-Iran, MN696622-Uzbekistan, MN696621-Uzbekistan, MN696620-Uzbekistan, MN696619-Uzbekistan, MN696606-Uzbekistan, MN696596-Uzbekistan, MN696591-Uzbekistan, MN696583-Uzbekistan, MN696572-Uzbekistan, MN696570-Uzbekistan, JF946609-Peru, JF946608-Peru, JF946607-Peru, JF946606-Peru, JF946605-Peru, JF946604-Peru, JF946603-Peru, JF946602-Peru, JF946601-Peru, JF946600-Peru, JF946599-Peru, JF946598-Peru, JF946597-Peru, KR349044-Algeria, KR349042-Algeria, KR349038-Algeria, MT239138-Slovenia, MT239133-Slovenia, KT780300-Poland, KT780298-Poland, JF946624-Peru, MH050629-China, MH050628-China, MH050626-China, MH050625-China, MH050622-China, MH050621-China, MH050620-China, KJ556994-China, KJ556993-China, MN231834-Iraq, EF367298-Morocco, MG672293-Algeria, MG672292-Algeria, MG672291-Algeria, MG672289-Algeria, MG672288-Algeria, MG672287-Algeria, MG672285-Algeria, MG672284-Algeria, MG672283-Algeria, MG672276-Tunisia, MG672275-Tunisia, MG672274-Tunisia, MG672273-Tunisia, MG672272-Tunisia, MG672271-Tunisia, MG672270-Tunisia, MG672268-Tunisia, MG672267-Tunisia, MG672266-Tunisia, MG672265-Tunisia, MG672264-Tunisia, MG672257-Kazakhstan, MG672255-Mongolia, MG672254-Mongolia, MG672245-Iran, MG672138-Romania, MG672135-Italy, MG672132-Finland, MG672129-Spain, MG672128-Algeria, KY766884-Finland, KU925429-Finland |
| Hap02 | 1 | EU072114-China |
| Hap03 | 2 | EU072113-China, JF836798-Iran |
| Hap04 | 2 | EU072112-China, AY572548-China |
| Hap05 | 15 | JF836800-Iran, MN696613-Uzbekistan, MN696612-Uzbekistan, MN696610-Uzbekistan, MN696609-Uzbekistan, MN696607-Uzbekistan, MN696599-Uzbekistan, MN696598-Uzbekistan, MN696590-Uzbekistan, MN696589-Uzbekistan, MN696587-Uzbekistan, MN696585-Uzbekistan, MN696584-Uzbekistan, MN696582-Uzbekistan, MN696581-Uzbekistan |
| Hap06 | 1 | MN696615-Uzbekistan |
| Hap07 | 2 | MN696611-Uzbekistan, MN696608-Uzbekistan |
| Hap08 | 1 | MN696604-Uzbekistan |
| Hap09 | 1 | MN696603-Uzbekistan |
| Hap10 | 1 | MN696602-Uzbekistan |
| Hap11 | 1 | MN696601-Uzbekistan |
| Hap12 | 1 | MN696600-Uzbekistan |
| Hap13 | 1 | MN696595-Uzbekistan |
| Hap14 | 1 | MN696594-Uzbekistan |
| Hap15 | 1 | MN696593-Uzbekistan |
| Hap16 | 1 | MN696592-Uzbekistan |
| Hap17 | 1 | MN696588-Uzbekistan |
| Hap18 | 1 | MN696586-Uzbekistan |
| Hap19 | 1 | MN696580-Uzbekistan |
| Hap20 | 1 | MN696579-Uzbekistan |
| Hap21 | 1 | MN696578-Uzbekistan |
| Hap22 | 1 | MN696577-Uzbekistan |
| Hap23 | 1 | MN696576-Uzbekistan |
| Hap24 | 1 | KT316342-Algeria |
| Hap25 | 1 | KR349043-Algeria |
| Hap26 | 2 | KR349041-Algeria, MG672286-Algeria |
| Hap27 | 2 | KR349040-Algeria, MG672290-Algeria |
| Hap28 | 2 | KR349039-Algeria, MG672269-Tunisia |
| Hap29 | 1 | MT239142-Slovenia |
| Hap30 | 1 | MT239140-Slovenia |
| Hap31 | 1 | MT239135-Slovenia |
| Hap32 | 1 | MT239134-Slovenia |
| Hap33 | 1 | MH050627-China |
| Hap34 | 1 | MH050624-China |
| Hap35 | 1 | MH050623-China |
| Hap36 | 1 | MN231833-Iraq |
| Hap37 | 2 | MG672137-Spain KU925414-Spain |
| n | H | hd ± SD | πd ± SD | Tajima’s D | p Value | Fu’s Fs | p Value | FLD | p Value | FLF | p Value | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mt-CO1 | 133 | 34 | 0.640 ± 0.048 | 0.00255 ± 0.00031 | −2.47269 | *. p < 0.01 | −49.797 | 0.000 | −3.97170 | p < 0.02 | −4.03871 | p < 0.02 |
| mt-ND1 | 140 | 37 | 0.639 ± 0.047 | 0.00611 ± 0.00147 | −2.80355 | *. p < 0.001 | −31.231 | 0.000 | −8.50154 | p < 0.02 | −7.14019 | p < 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selcuk, M.A.; Celik, F.; Kesik, H.K.; Gunyakti Kilinc, S.; Ahmed, H.; Jiang, N.; Simsek, S.; Cao, J. In Silico Evaluation of the Haplotype Diversity, Phylogenetic Variation and Population Structure of Human E. granulosus sensu stricto (G1 Genotype) Sequences. Pathogens 2022, 11, 1346. https://doi.org/10.3390/pathogens11111346
Selcuk MA, Celik F, Kesik HK, Gunyakti Kilinc S, Ahmed H, Jiang N, Simsek S, Cao J. In Silico Evaluation of the Haplotype Diversity, Phylogenetic Variation and Population Structure of Human E. granulosus sensu stricto (G1 Genotype) Sequences. Pathogens. 2022; 11(11):1346. https://doi.org/10.3390/pathogens11111346
Chicago/Turabian StyleSelcuk, Muhammed Ahmed, Figen Celik, Harun Kaya Kesik, Seyma Gunyakti Kilinc, Haroon Ahmed, Nan Jiang, Sami Simsek, and Jianping Cao. 2022. "In Silico Evaluation of the Haplotype Diversity, Phylogenetic Variation and Population Structure of Human E. granulosus sensu stricto (G1 Genotype) Sequences" Pathogens 11, no. 11: 1346. https://doi.org/10.3390/pathogens11111346