

Tight Junctions, the Key Factor in Virus-Related Disease
 ,
,
Abstract
:1. Introduction
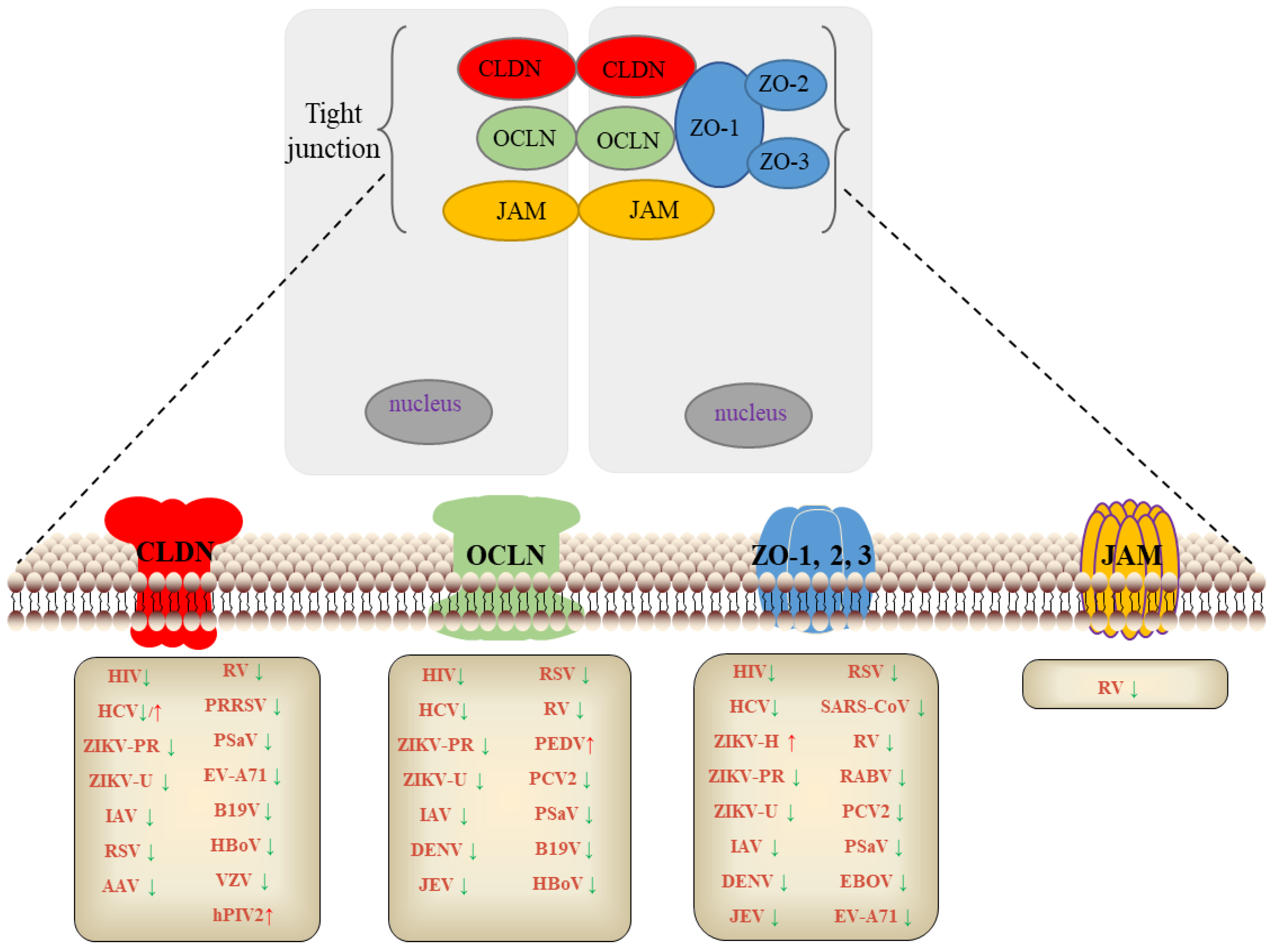
2. TJs and Virus
2.1. TJs and Human Immunodeficiency Virus (HIV)
2.2. TJs and Flavivirus
2.2.1. HCV
2.2.2. ZIKV
2.2.3. DENV
2.2.4. JEV
2.3. TJs and Rabies Virus (RABV)
2.4. TJs and Influenza A Virus (IV)
2.5. TJs and Respiratory Syncytial Virus (RSV)
2.6. TJs and SARS Coronavirus (SARS-CoV)
2.7. TJs and Adeno-Associated Viruses (AAV)
2.8. TJs and Rotavirus (RV)
2.9. TJs and Porcine Epidemic Diarrhea Virus (PEDV)
2.10. TJs and Porcine Reproductive and Respiratory Syndrome Virus (PRRSV)
2.11. TJs and Porcine Circovirus Type 2 Virus (PCV2)
2.12. TJs and Porcine Sapovirus (PoSaV)
2.13. TJs and Other Pathogens

{kind=link}
{kind=link}
| Viruses | Viral Genome | Model/Cells | Tight Junctions | Up/Down Regulate | Effects on Virus Infection | References |
|---|---|---|---|---|---|---|
| HIV | ssRNA | Rhesus monkeys, Rat, HBMEC | OCLN, ZO-1, CLDN-1, 2, 3, 4, 5 | ↓ NF-κB and MAPK pathways involved in IL-17, IL-6, TNF-α | Inhibiting | [12,13,14,15,16,17,18,19,20,21,22,23,24,25,26] |
| HCV | Positive ssRNA | Mice | CLDN-1, OCLD, ZO-1 | ↓ Contributes to the internalization of virus, and the EC2 and C-termini of OCLN, involved in HCV infection | Promoting | [28,29,30,31,32,33,34,35,36,37,38] |
| HCV | Positive ssRNA | Huh7.5.1 cells | CLDN-6, 9, 12 | ↑ Interact with the major HCV receptor CD81 | Promoting | [32,33,34] |
| ZIKV-H | Positive ssRNA | Mice | ZO-1 | ↑ Upregulates the expression of ZO-1 in human BBB | Promoting | [42,43] |
| ZIKV-PR | Positive ssRNA | Mice | ZO-1, OCLD, CLDN-5 | ↓ Downregulated the expression of ZO-1, OCLN, and CLDN-5 and penetrated into the brain parenchyma early after infection | Inhibiting | [42,43,44,45,46,47,48] |
| ZIKV-U | Positive ssRNA | Mice | ZO-1, OCLD, CLDN-5 | Inhibiting | [42,43,44,45,46,47,48] | |
| DENV | Positive ssRNA | human endothelial | ZO-1 | ↓ Downregulates the expression of TNF-α, cause a significant decrease in ZO-1 expression and peripheral localization in epithelial and endothelial cells | Inhibiting | [49,50,51,52,53] |
| JEV | Positive ssRNA | Mice | ZO-1, OCLN, CLDN-5 | ↓ Downregulation of OCLN, CLDN5 and ZO-1 expression; IP10 induces TNF-a, which leads to BBB destruction. | Inhibiting | [54] |
| RABV | Negative ssRNA | BHK-21 | ZO-1 | ↓ IFN-λ is regulated by RABV, which maintains ZO-1 expression in the BBB, thus ensuring the integrity of the TJ | Inhibiting | [55] |
| IAV | 8-segment negative ssRNA | Ferret, Mice | OCLN, ZO-1, CLDN-1,3 | ↓ Destructs BBB; alter the cytoskeleton and morphology of airway epithelial cells | Inhibiting | [56,57,58,59,60,61,62,63,64,65] |
| RSV | Negative ssRNA | Mice | OCLN, ZO-1, CLDN-1 | ↓ Causes airway epithelial barrier dysfunction, leading to an immune response and airway inflammation enhancement | Inhibiting | [66,67,68] |
| SRAS-CoV | Positive ssRNA | MDCK | ZO-1 | ↓ Affects the integrity of lung epithelial cells by downregulating the expression of ZO-1 | Inhibiting | [69,70] |
| AAV | ssDNA | Mice | ZO-1, OCLN, CLDN | ↓ Downregulating CLDN11 | Inhibiting | [71,72,73,74,75,76,77,78] |
| RV | 11-segment dsRNA | Humans, Mice | OCLN, CLDN, JAM-A, ZO-1 | ↓ Induces redistribution of OCLN, CLND, JAM-A and ZO-1 into the cytoplasm and increases accessory cell permeability of polarized MDCK cells | Inhibiting | [79,80] |
| PEDV | Positive ssRNA | Vero-E6, IPEC-J2 | OCLN | ↑ Viral entry is closely associated with internalization of OCLN | Promoting | [81] |
| PRRSV | Positive ssRNA | Marc-145, PAM | CLDN-4 | ↓ Inhibits virus adsorption, PRRSV GP3 can interact with the second loop ECL2 of CLDN4 | Inhibiting | [9] |
| PCV2 | ssDNA | Pig | ZO-1, OCLD | ↓ PCV2 downregulates ZO-1 and OCLN in the lung through JNK/MAPK pathway. | Inhibiting | [84] |
| PoSaV | Positive ssRNA | Intestinal epithelial cells | ZO-1, 2, CLDN-1 | ↓ PSaV enters the target cells by inducing early dissociation of TJs, which binds to the OCLN that acts as a functional co-receptor | Inhibiting | [85,86] |
| EBOV | Negative ssRNA | Sertoli | ZO-1, 2 | ↓ Disruption of BTBs formed by intercellular TJs in testes through reduces the expression of ZO-1 and ZO-2 | Inhibiting | [87] |
| EV-A71 | Positive ssRNA | Mice | ZO-1, CLDN-5 | ↓ VP1 downregulates CLDN5 | Inhibiting | [88,89] |
| B19V | ssDNA | Mice | CLDN-1, OCLN | ↓ Reduces the expression of CLDN-1 and OCLN | Inhibiting | [90] |
| HBoV | ssDNA | Mice | CLDN-1, OCLN | ↓ Reduces the expression of CLDN-1 and OCLN | Inhibiting | [90] |
| VZV | dsDNA | Macaques | CLDN-2, 10, 18 | ↓ Reduces the expression of CLDN2, CLDN10 and CLDN18 in alveolar epithelial cells | Inhibiting | [91] |
| hPIV2 | Negative ssRNA | Airway epithelial cells | CLDN1 | ↑ Induces the expression of CLDN1 in airway epithelial cells | Promoting | [95] |
| HEV | Positive ssRNA | HBMVEC | CLDN5, OCLN and ZO-1 | ↓ Reduces the expression of CLDN-5, OCLN and ZO-1 | Inhibiting | [96] |
3. Antiviral Drugs Target TJs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Diamond, J.M. Twenty-first Bowditch lecture. The epithelial junction: Bridge, gate, and fence. Physiologist 1977, 20, 10–18. [Google Scholar]
- Zihni, C.; Mills, C.; Matter, K.; Balda, M.S. Tight junctions: From simple barriers to multifunctional molecular gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564–644. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Mariscal, L.; Betanzos, A.; Nava, P.; Jaramillo, B.E. Tight junction proteins. Prog. Biophys. Mol. Biol. 2003, 81, 1–44. [Google Scholar] [CrossRef]
- Barbara, G.; Barbaro, M.R.; Fuschi, D.; Palombo, M.; Falangone, F.; Cremon, C.; Marasco, G.; Stanghellini, V. Inflammatory and Microbiota-Related Regulation of the Intestinal Epithelial Barrier. Front. Nutr. 2021, 13, 718356–718364. [Google Scholar] [CrossRef] [PubMed]
- Czupalla, C.J.; Liebner, S.; Devraj, K. In vitro models of the blood-brain barrier. Methods Mol. Biol. 2014, 1135, 415–437. [Google Scholar] [PubMed]
- Mital, P.; Hinton, B.T.; Dufour, J.M. The blood-testis and blood-epididymis barriers are more than just their tight junctions. Biol. Reprod. 2011, 84, 851–859. [Google Scholar] [CrossRef] [Green Version]
- Gong, S.; Cao, G.; Li, F.; Chen, Z.; Pan, X.; Ma, H.; Zhang, Y.; Yu, B.; Kou, J. Endothelial Conditional Knockdown of NMMHC IIA (Nonmuscle Myosin Heavy Chain IIA) Attenuates Blood-Brain Barrier Damage During Ischemia-Reperfusion Injury. Stroke 2021, 52, 1053–1064. [Google Scholar] [CrossRef] [PubMed]
- Torres-Flores, J.M.; Arias, C.F. Tight Junctions Go Viral! Viruses 2015, 7, 5145–5154. [Google Scholar] [CrossRef] [PubMed]
- Ding, G.; Liu, J.; Shao, Q.; Wang, B.; Feng, J.; Li, Y.; Li, L.; Cao, S.; Cong, F.; Zhao, Y.; et al. Porcine Reproductive and Respiratory Syndrome Virus Structural Protein GP3 Regulates Claudin 4 To Facilitate the Early Stages of Infection. J. Virol. 2020, 94, e00124-20. [Google Scholar] [CrossRef] [PubMed]
- Yusuke, K.; Youhei, Y.; Satomi, S.; Shinji, K.; Fumio, S.; Yoko, N.; Takeo, U. MA026, an anti-hepatitis C virus compound, opens tight junctions of the epithelial cell membrane. J. Antibiot. 2018, 70, 691–694. [Google Scholar]
- Laura, R.; Ok-Ryul, S.; Jannick, P.; François, H.; Laurent, L.; Charles, H.G.; Alexandre, V.; Lucie, F.; Sandrine, B.; Thomas, F.B.; et al. Identification of Piperazinylbenzenesulfonamides as New Inhibitors of Claudin-1 Trafficking and Hepatitis C Virus Entry. J. Virol. 2018, 92, e01982-17. [Google Scholar]
- Passmore, J.A.; Jaspan, H.B.; Masson, L. Genital inflammation, immune activation and risk of sexual HIV acquisition. Curr. Opin. HIV AIDS 2016, 11, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Chand, H.S.; Vazquez-Guillamet, R.; Royer, C.; Rudolph, K.; Mishra, N.; Singh, S.P.; Hussain, S.S.; Barrett, E.; Callen, S.; Byrareddy, S.N.; et al. Cigarette smoke and HIV synergistically affect lung pathology in cynomolgus macaques. J. Clin. Investig. 2018, 128, 5428–5433. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Torben, W.; Mansfield, J.; Alvarez, X.; Vande Stouwe, C.; Li, J.; Byrareddy, S.N.; Didier, P.J.; Pahar, B.; Molina, P.E.; et al. Cannabinoid Attenuation of Intestinal Inflammation in Chronic SIV-Infected Rhesus Macaques Involves T Cell Modulation and Differential Expression of Micro-RNAs and Pro-inflammatory Genes. Front. Immunol. 2019, 10, 914. [Google Scholar] [CrossRef]
- Banks, W.A.; Robinson, S.M.; Nath, A. Permeability of the blood–brain barrier to HIV-1 Tat. Exp. Neurol. 2005, 193, 218–245. [Google Scholar] [CrossRef]
- Hudson, L.; Liu, J.; Nath, A.; Jones, M.; Raghavan, R.; Narayan, O.; Male, D.; Everall, I. Detection of the human immunodeficiency virus regulatory protein tat in CNS tissues. J. Neurovirol. 2000, 6, 145–155. [Google Scholar] [CrossRef]
- Zou, M.; Huang, W.; Jiang, W.; Wu, Y.; Chen, Q. Role of Cav-1 in HIV-1 Tat-Induced Dysfunction of Tight Junctions and Abeta-Transferring Proteins. Oxid. Med. Cell Longev. 2019, 2019, 3403206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, V.H.; Kafka, J.K.; Kaushic, C. Influence of Common Mucosal Co-Factors on HIV Infection in the Female Genital Tract. Am. J. Reprod. Immunol. 2014, 71, 543–554. [Google Scholar] [CrossRef]
- Nazli, A.; Dizzell, S.; Zahoor, M.A.; Ferreira, V.H.; Kafka, J.; Woods, M.W.; Ouellet, M.; Ashkar, A.A.; Tremblay, M.J.; Bowdish, D.M.; et al. Interferon-beta induced in female genital epithelium by HIV-1 glycoprotein 120 via Toll-like-receptor 2 pathway acts to protect the mucosal barrier. Cell Mol. Immunol. 2019, 16, 178–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, A.; Graves, J.; Reidy, M.A. Mitogen-activated protein kinases mediate matrix metal-loproteinase-9 expression in vascular smooth muscle cells. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 2527–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, S.M.; Song, H.Y.; Lee, J.A.; Lee, S.J.; Choi, S.Y. Extracellular HIV-1 Tat up-regulates ex-pression of matrix metalloproteinase-9 via a MAPK-NF-κB dependent pathway in human astrocytes. Exp. Mol. Med. 2009, 41, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.; Lyons, J.G.; Tan, T.K.; Wang, Y.; Hsu, T.T.; Min, D.; Succar, L.; Rangan, G.K.; Hu, M.; Henderson, B.R.; et al. Disruption of E-cadherin by matrix metalloproteinase directly mediates epithelial-mesenchymal transition downstream of transforming growth factor-beta1 in renal tubular epithelial cells. Am. J. Pathol. 2009, 175, 580–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symowicz, J.; Adley, B.P.; Gleason, K.J.; Johnson, J.J.; Ghosh, S.; Fishman, D.A.; Hudson, L.G.; Stack, M.S. Engagement of collagen-binding integrins promotes matrix metalloproteinase-9-dependent E-cadherin ecto-domain shedding in ovarian carcinoma cells. Cancer Res. 2007, 67, 2030–2039. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.T.; Bürgers, H.F.; Rabie, T.; Marti, H.H. Matrix metalloproteinase-9 mediates hypox-ia-induced vascular leakage in the brain via tight junction rearrangement. J. Cereb. Blood. Flow. Metab. 2010, 30, 837–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sufiawati, I.; Tugizov, S.M. HIV-induced matrix metalloproteinase-9 activation through mitogen-activated protein kinase signalling promotes HSV-1 cell-to-cell spread in oral epithelial cells. J. Gen. Virol. 2018, 99, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Cui, Y.; Shen, X.; Wang, S.; Yang, G.B. IL-17A and IL-17F repair HIV-1 gp140 damaged Caco-2 cell barriers by upregulating tight junction genes. Microbes Infect. 2019, 21, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Hinderfeld, A.S.; Phukan, N.; Bar, A.K.; Roberton, A.M.; Simoes-Barbosa, A. Cooperative Interactions between Trichomonas vaginalis and Associated Bacteria Enhance Paracellular Permeability of the Cervicovaginal Epithelium by Dysregulating Tight Junctions. Infect. Immun. 2019, 87, e00141-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, Q.; von Schaewen, M.; Hrebikova, G.; Heller, B.; Sandmann, L.; Plaas, M.; Ploss, A. Mice Expressing Minimally Humanized CD81 and Occludin Genes Support Hepatitis C Virus Uptake In Vivo. J. Virol. 2017, 91, e01799-16. [Google Scholar] [CrossRef] [Green Version]
- Ciesek, S.; von Hahn, T. Expression Cloning of Host Factors Required for the HCV Replication Cycle. Methods Mol. Biol. 2019, 1911, 169–182. [Google Scholar] [PubMed]
- Riad, S.E.; Elhelw, D.S.; Shawer, H.; El-Ekiaby, N.; Salah, A.; Zekri, A.; Esmat, G.; Amleh, A.; Abdelaziz, A.I. Disruption of Claudin-1 Expression by miRNA-182 Alters the Susceptibility to Viral Infectivity in HCV Cell Models. Front. Genet. 2018, 9, 93. [Google Scholar] [CrossRef] [Green Version]
- Belouzard, S.; Danneels, A.; Feneant, L.; Seron, K.; Rouille, Y.; Dubuisson, J. Entry and Release of Hepatitis C Virus in Polarized Human Hepatocytes. J. Virol. 2017, 91, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haid, S.; Grethe, C.; Dill, M.T.; Heim, M.; Kaderali, L.; Pietschmann, T. Isolate-dependent use of claudins for cell entry by hepatitis C virus. Hepatology 2014, 59, 24–34. [Google Scholar] [CrossRef]
- Huang, J.; Yin, P.; Zhang, L. COPII cargo claudin-12 promotes hepatitis C virus entry. J. Viral. Hepat. 2019, 26, 308–320. [Google Scholar] [CrossRef] [PubMed]
- Baktash, Y.; Madhav, A.; Coller, K.E.; Randall, G. Single Particle Imaging of Polarized Hepatoma Organoids upon Hepatitis C Virus Infection Reveals an Ordered and Sequential Entry Process. Cell Host Microbe 2018, 23, 382–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okai, K.; Ichikawa-Tomikawa, N.; Saito, A.C.; Watabe, T.; Sugimoto, K.; Fujita, D.; Ono, C.; Fukuhara, T.; Matsuura, Y.; Ohira, H.; et al. A novel occludin-targeting monoclonal antibody prevents hepatitis C virus infection in vitro. Oncotarget 2018, 9, 16588–16598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavie, M.; Linna, L.; Moustafa, R.I.; Belouzard, S.; Fukasawa, M.; Dubuisson, J. Role of the cytosolic domain of occludin in trafficking and hepatitis C virus infection. Traffic 2019, 20, 753–773. [Google Scholar] [CrossRef]
- Mailly, L.; Baumert, T.F. Hepatitis C virus infection and tight junction proteins: The ties that bind. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183296. [Google Scholar] [CrossRef]
- Gerold, G.; Moeller, R.; Pietschmann, T. Hepatitis C Virus Entry: Protein Interactions and Fusion Determinants Governing Productive Hepatocyte Invasion. Cold Spring Harb. Perspect. Med. 2020, 10, a036830. [Google Scholar] [CrossRef]
- Shao, T.; Zhao, C.; Li, F.; Gu, Z.; Liu, L.; Zhang, L.; Wang, Y.; He, L.; Liu, Y.; Liu, Q.; et al. Intestinal HIF-1alpha deletion exacerbates alcoholic liver disease by inducing intestinal dysbiosis and barrier dysfunction. J. Hepatol. 2018, 69, 886–895. [Google Scholar] [CrossRef]
- Sekhar, V.; Pollicino, T.; Diaz, G.; Engle, R.E.; Alayli, F.; Melis, M.; Kabat, J.; Tice, A.; Pomerenke, A.; Altan-Bonnet, N.; et al. Infection with hepatitis C virus depends on TACSTD2, a regulator of claudin-1 and occludin highly downregulated in hepatocellular carcinoma. PLoS Pathog. 2018, 14, e1006916. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.F.; Chu, L.W.; Liao, I.C.; Simanjuntak, Y.; Lin, Y.L.; Juan, C.C.; Ping, Y.H. The Mechanism of the Zika Virus Crossing the Placental Barrier and the Blood-Brain Barrier. Front. Microbiol. 2020, 11, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leda, A.R.; Bertrand, L.; Andras, I.E.; El-Hage, N.; Nair, M.; Toborek, M. Selective Disruption of the Blood-Brain Barrier by Zika Virus. Front. Microbiol. 2019, 10, 2158. [Google Scholar] [CrossRef]
- Rastogi, M.; Singh, S.K. Zika virus NS1 affects the junctional integrity of human brain microvascular endothelial cells. Biochimie 2020, 176, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.; Martin-Tapia, D.; Valdespino-Vazquez, Y.; Alarcon, L.; Espejel-Nunez, A.; Guzman-Huerta, M.; Munoz-Medina, J.E.; Shibayama, M.; Chavez-Munguia, B.; Estrada-Gutierrez, G.; et al. Syncytiotrophoblast of Placentae from Women with Zika Virus Infection Has Altered Tight Junction Protein Expression and Increased Paracellular Permeability. Cells 2019, 8, 1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, L.; Nie, Y.; Li, S.; Guo, M.; Yang, W.; Huang, R.; Chen, J.; Liu, Y.; Lu, X.; Chen, Z.; et al. Matrix metalloproteinase 9 facilitates Zika virus invasion of the testis by modulating the integrity of the blood-testis barrier. PLoS Pathog. 2020, 16, e1008509. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Z.Y.; Gao, N.; Wang, Z.Y.; Cui, X.Y.; Zhou, D.S.; Fan, D.Y.; Chen, H.; Wang, P.G.; An, J. Sertoli Cells Are Susceptible to ZIKV Infection in Mouse Testis. Front. Cell Infect. Microbiol. 2017, 7, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, M.U.; Zahedi-Amiri, A.; Glover, K.K.M.; Gao, A.; Nickol, M.E.; Kindrachuk, J.; Wilkins, J.A.; Coombs, K.M. Zika virus dysregulates human Sertoli cell proteins involved in spermatogenesis with little effect on tight junctions. PLoS Negl. Trop. Dis. 2020, 14, e0008335. [Google Scholar] [CrossRef] [PubMed]
- Tamhankar, M.; Patterson, J.L. Directional entry and release of Zika virus from polarized epithelial cells. Virol. J. 2019, 16, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanthick, C.; Suttitheptumrong, A.; Rawarak, N.; Pattanakitsakul, S.-N. Transcytosis Involvement in Transport System and Endothelial Permeability of Vascular Leakage during Dengue Virus Infection. Viruses 2018, 10, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inyoo, S.; Suttitheptumrong, A.; Pattanakitsakul, S.-N. Synergistic Effect of TNF-α and Dengue Virus Infection on Adhesion Molecule Reorganization in Human Endothelial Cells. Jpn. J. Infect. Dis. 2017, 70, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Carod-Artal, F.J.; Wichmann, O.; Farrar, J.; Gascón, J. Neurological complications of dengue virus infection. Lancet Neurol. 2013, 12, 906–919. [Google Scholar] [CrossRef]
- Mustafa, Y.M.; Meuren, L.M.; Coelho, S.V.A.; de Arruda, L.B. Pathways Exploited by Flaviviruses to Counteract the Blood-Brain Barrier and Invade the Central Nervous System. Front. Microbiol. 2019, 10, 525. [Google Scholar] [CrossRef]
- Idris, F.; Muharram, S.H.; Zaini, Z.; Alonso, S.; Diah, S. Invasion of a murine in vitro blood-brain barrier co-culture model by dengue virus serotypes 1 to 4. Arch. Virol. 2019, 164, 1069–1083. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Wang, H.; Lou, W.; Ma, L.; Li, Y.; Zhang, N.; Wang, C.; Li, F.; Awais, M.; Cao, S.; et al. IP-10 Promotes Blood-Brain Barrier Damage by Inducing Tumor Necrosis Factor Alpha Production in Japanese Encephalitis. Front. Immunol. 2018, 9, 1148. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, L.; Luo, Z.; Zhang, Y.; Lv, L.; Zhao, J.; Sui, B.; Huang, F.; Cui, M.; Fu, Z.F.; et al. Interferon-lambda Attenuates Rabies Virus Infection by Inducing Interferon-Stimulated Genes and Alleviating Neurological Inflammation. Viruses 2020, 12, 405. [Google Scholar] [CrossRef] [Green Version]
- Short, K.R.; Kasper, J.; van der Aa, S.; Andeweg, A.C.; Zaaraoui-Boutahar, F.; Goeijenbier, M.; Richard, M.; Herold, S.; Becker, C.; Scott, D.P.; et al. Influenza virus damages the alveolar barrier by disrupting epithelial cell tight junctions. Eur. Respir. J. 2016, 47, 954–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pharo, E.A.; Williams, S.M.; Boyd, V.; Sundaramoorthy, V.; Durr, P.A.; Baker, M.L. Host-Pathogen Responses to Pandemic Influenza H1N1pdm09 in a Human Respiratory Airway Model. Viruses 2020, 12, 679. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Zi, X.; Peng, Y.; Wang, Z.; Hong, H.; Yan, Y.; Guan, W.; Tan, K.S.; Liu, J.; Ong, H.H.; et al. H3N2 influenza virus infection enhances oncostatin M expression in human nasal epithelium. Exp. Cell Res. 2018, 371, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Goldsmith, C.S.; Kumar, A.; Belser, J.A.; Sun, X.; Pappas, C.; Brock, N.; Bai, Y.; Levine, M.; Tumpey, T.M.; et al. Tropism and Infectivity of a Seasonal A(H1N1) and a Highly Pathogenic Avian A(H5N1) Influenza Virus in Primary Differentiated Ferret Nasal Epithelial Cell Cultures. J. Virol. 2019, 93, e00080-19. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Liu, X.; Chen, F.; Zuo, K.; Wu, C.; Yan, Y.; Chen, W.; Lin, W.; Xie, Q. Avian Influenza Virus Subtype H9N2 Affects Intestinal Microbiota, Barrier Structure Injury, and Inflammatory Intestinal Disease in the Chicken Ileum. Viruses 2018, 10, 270. [Google Scholar] [CrossRef] [Green Version]
- Smirnova, S.S.; Pisareva, M.M.; Smirnova, T.D.; Sivak, K.V.; Vorobiev, K.V. Long-Term Maintenance of the Functional Changes Induced by Influenza A Virus and/or LPS in Human Endothelial ECV-304 Cell Sublines. Cell Tissue Biol. 2019, 13, 283–291. [Google Scholar] [CrossRef]
- Günzel, D.; Yu, A.S. Claudins and the modulation of tight junction permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokuhara, Y.; Morinishi, T.; Matsunaga, T.; Sakai, M.; Sakai, T.; Ohsaki, H.; Kadota, K.; Kushida, Y.; Haba, R.; Hirakawa, E. Nuclear expression of claudin-3 in human colorectal adeno-carcinoma cell lines and tissues. Oncol. Lett. 2018, 15, 99–108. [Google Scholar] [PubMed]
- Sreenivasan, C.C.; Thomas, M.; Antony, L.; Wormstadt, T.; Hildreth, M.B.; Wang, D.; Hause, B.; Francis, D.H.; Li, F.; Kaushik, R.S. Development and characterization of swine primary respiratory epithelial cells and their susceptibility to infection by four influenza virus types. Virology 2019, 528, 152–163. [Google Scholar] [CrossRef]
- Liu, J.; Wang, H.; Fang, M.; Chen, X.; Zeng, X. A human cell polarity protein Lgl2 regulates influenza A virus nucleoprotein exportation from nucleus in MDCK cells. J. Biosci. 2020, 45, 67. [Google Scholar] [CrossRef]
- Deng, Y.; Herbert, J.A.; Robinson, E.; Ren, L.; Smyth, R.L.; Smith, C.M. Neutrophil-Airway Epithelial Interactions Result in Increased Epithelial Damage and Viral Clearance during Respiratory Syncytial Virus Infection. J. Virol. 2020, 94, e02161-19. [Google Scholar] [CrossRef] [Green Version]
- Smallcombe, C.C.; Linfield, D.T.; Harford, T.J.; Bokun, V.; Ivanov, A.I.; Piedimonte, G.; Rezaee, F. Disruption of the airway epithelial barrier in a murine model of respiratory syncytial virus infection. Am. J. Physiol. Lung Cell Mol. Physiol. 2019, 316, L358–L368. [Google Scholar] [CrossRef]
- Liu, J.J.; Zhang, T.; Mi, Y.M. Effects of respiratory syncytial virus infection on epidermal growth factor receptor, tight junction association proteins and mucin in airway epithelial cells. Zhongguo Dang Dai Er Ke Za Zhi 2019, 21, 294–303. [Google Scholar] [PubMed]
- Toto, A.; Ma, S.; Malagrino, F.; Visconti, L.; Pagano, L.; Stromgaard, K.; Gianni, S. Comparing the binding properties of peptides mimicking the Envelope protein of SARS-CoV and SARS-CoV-2 to the PDZ domain of the tight junction-associated PALS1 protein. Protein Sci. 2020, 29, 2038–2042. [Google Scholar] [CrossRef] [PubMed]
- Teoh, K.T.; Siu, Y.L.; Chan, W.L.; Schluter, M.A.; Liu, C.J.; Peiris, J.S.; Bruzzone, R.; Margolis, B.; Nal, B. The SARS coronavirus E protein interacts with PALS1 and alters tight junction formation and epithelial morphogenesis. Mol. Biol. Cell 2010, 21, 3838–3852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foust, K.D.; Nurre, E.; Montgomery, C.L.; Hernandez, A.; Chan, C.M.; Kaspar, B.K. Intravascular AAV9 preferentially targets neonatal neurons and adult astrocytes. Nat. Biotechnol. 2009, 27, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogg, V.C.; Liu, C.J.; Margolis, B. Multiple regions of Crumbs3 are required for tight junction formation in MCF10A cells. J. Cell Sci. 2005, 118, 2859–2869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madigan, V.J.; Tyson, T.O.; Yuziuk, J.A.; Pillai, M.; Moller-Tank, S.; Asokan, A. A CRISPR Screen Identifies the Cell Polarity Determinant Crumbs 3 as an Adeno-associated Virus Restriction Factor in Hepatocytes. J. Virol. 2019, 93, e00943-19. [Google Scholar] [CrossRef] [PubMed]
- Szymaniak, A.D.; Mahoney, J.E.; Cardoso, W.V.; Varelas, X. Crumbs3-Mediated Polarity Directs Airway Epithelial Cell Fate through the Hippo Pathway Effector Yap. Dev Cell 2015, 34, 283–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrade, A.; Sade, H.; Couraud, P.O.; Romero, I.A.; Weksler, B.B.; Niewoehner, J. Expression and localization of claudins-3 and -12 in transformed human brain endothelium. Fluids Barriers CNS 2012, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Holdcraft, R.W.; Shima, J.E.; Griswold, M.D.; Braun, R.E. Androgens regulate the permeability of the blood-testis barrier. Proc. Natl. Acad. Sci. USA 2005, 102, 16696–16700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrow, C.M.; Tyagi, G.; Simon, L.; Carnes, K.; Murphy, K.M.; Cooke, P.S.; Hofmann, M.C.; Hess, R.A. Claudin 5 expression in mouse seminiferous epithelium is dependent upon the transcription factor ets variant 5 and contributes to blood-testis barrier function. Biol. Reprod. 2009, 81, 871–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, S.; Kanatsu-Shinohara, M.; Ogonuki, N.; Matoba, S.; Ogura, A.; Shinohara, T. In Vivo Genetic Manipulation of Spermatogonial Stem Cells and Their Microenvironment by Adeno-Associated Viruses. Stem Cell Rep. 2018, 10, 1551–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soliman, M.; Cho, E.H.; Park, J.G.; Kim, J.Y.; Alfajaro, M.M.; Baek, Y.B.; Kim, D.S.; Kang, M.I.; Park, S.I.; Cho, K.O. Rotavirus-Induced Early Activation of the RhoA/ROCK/MLC Signaling Pathway Mediates the Disruption of Tight Junctions in Polarized MDCK Cells. Sci. Rep. 2018, 8, 13931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagbom, M.; De Faria, F.M.; Winberg, M.E.; Westerberg, S.; Nordgren, J.; Sharma, S.; Keita, A.V.; Loitto, V.; Magnusson, K.E.; Svensson, L. Neurotrophic Factors Protect the Intestinal Barrier from Rotavirus Insult in Mice. mBio 2020, 11, e02834-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Guo, L.; Zhang, J.; Xu, Y.; Gu, W.; Feng, L.; Wang, Y. Tight Junction Protein Occludin Is a Porcine Epidemic Diarrhea Virus Entry Factor. J. Virol. 2017, 91, e00202-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkard, C.; Opriessnig, T.; Mileham, A.J.; Stadejek, T.; Ait-Ali, T.; Lillico, S.G.; Whitelaw, C.B.A.; Archibald, A.L. Pigs Lacking the Scavenger Receptor Cysteine-Rich Domain 5 of CD163 Are Resistant to Porcine Reproductive and Respiratory Syndrome Virus 1 Infection. J. Virol. 2018, 92, e00415-18. [Google Scholar] [CrossRef] [Green Version]
- Du, T.; Nan, Y.; Xiao, S.; Zhao, Q.; Zhou, E.M. Antiviral Strategies against PRRSV Infection. Trends Microbiol. 2017, 25, 968–979. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhou, H.; Lin, H.; Ma, Z.; Fan, H. Porcine circovirus type 2 exploits JNK-mediated disruption of tight junctions to facilitate Streptococcus suis translocation across the tracheal epithelium. Vet. Res. 2020, 51, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Q.; Wang, C.; Liu, H.; Wu, Q.; Liang, S.; Cen, M.; Dong, Q.; Wei, Y.; Chen, Y.; Ouyang, K.; et al. Pathogenic Characteristics of a Porcine Astrovirus Strain Isolated in China. Viruses 2019, 11, 1156. [Google Scholar] [CrossRef] [Green Version]
- Alfajaro, M.M.; Cho, E.H.; Kim, D.S.; Kim, J.Y.; Park, J.G.; Soliman, M.; Baek, Y.B.; Park, C.H.; Kang, M.I.; Park, S.I.; et al. Early Porcine Sapovirus Infection Disrupts Tight Junctions and Uses Occludin as a Coreceptor. J. Virol. 2019, 93, e01773-18. [Google Scholar] [CrossRef] [Green Version]
- Coffin, K.M.; Liu, J.; Warren, T.K.; Blancett, C.D.; Kuehl, K.A.; Nichols, D.K.; Bearss, J.J.; Schellhase, C.W.; Retterer, C.J.; Weidner, J.M.; et al. Persistent Marburg Virus Infection in the Testes of Nonhuman Primate Survivors. Cell Host Microbe 2018, 24, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Jiao, J.; Wang, C.; Zhang, L. Epithelial physical barrier defects in chronic rhinosinusitis. Expert. Rev. Clin. Immunol. 2019, 15, 679–688. [Google Scholar] [CrossRef]
- Wang, W.; Sun, J.; Wang, N.; Sun, Z.; Ma, Q.; Li, J.; Zhang, M.; Xu, J. Enterovirus A71 capsid protein VP1 increases blood-brain barrier permeability and virus receptor vimentin on the brain endothelial cells. J. Neurovirol. 2020, 26, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Lin, C.-Y.; Chung, Y.-H.; Shi, Y.-F.; Tzang, B.-S.; Hsu, T.-C. The VP1 unique region of human parvovirus B19 and human bocavirus induce lung injury in naïve Balb/c mice. PLoS ONE 2018, 13, e0202667. [Google Scholar]
- Ouwendijk, W.J.; Van den Ham, H.J.; Delany, M.W.; Van Kampen, J.J.; Van Nierop, G.P.; Mehraban, T.; Zaaraoui-Boutahar, F.; Van, I.W.F.; Van den Brand, J.M.; De Vries, R.D.; et al. Alveolar barrier disruption in varicella pneumonia is associated with neutrophil extracellular trap formation. JCI Insight 2020, 5, e138900. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Qiu, K.; He, Q.; Lei, Q.; Lu, W. Mechanisms of Blood-Brain Barrier Disruption in Herpes Simplex Encephalitis. J. Neuroimmune Pharmacol. 2019, 14, 157–172. [Google Scholar] [CrossRef] [PubMed]
- Rahn, E.; Their, K.; Petermann, P.; Rubsam, M.; Staeheli, P.; Iden, S.; Niessen, C.M.; Knebel-Morsdorf, D. Epithelial Barriers in Murine Skin during Herpes Simplex Virus 1 Infection: The Role of Tight Junction Formation. J. Investig. Dermatol. 2017, 137, 884–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, I.; Shinozaki, E.; Sakata, S.; Yamamoto, N.; Fujisaki, J.; Muramatsu, Y.; Hirota, T.; Takeuchi, K.; Takahashi, S.; Yamaguchi, K.; et al. Enrichment of CLDN18-ARHGAP fusion gene in gastric cancers in young adults. Cancer Sci. 2019, 110, 1352–1363. [Google Scholar] [CrossRef] [PubMed]
- Yumine, N.; Matsumoto, Y.; Ohta, K.; Fukasawa, M.; Nishio, M. Claudin-1 inhibits human parainfluenza virus type 2 dissemination. Virology 2019, 531, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Shi, R.; Liu, T.; She, R.; Wu, Q.; An, J.; Hao, W.; Soomro, M.H. Brain Infection by Hepatitis E Virus Probably via Damage of the Blood-Brain Barrier Due to Alterations of Tight Junction Proteins. Front. Cell Infect. Microbiol. 2019, 9, 52. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, X.Q.; Zhang, B.; Gu, J.; Meng, F.Z.; Liu, H.; Zhou, L.; Wang, X.; Hou, W.; Ho, W.Z. Bowman Birk Inhibitor Suppresses Herpes Simplex Virus Type 2 Infection of Human Cervical Epithelial Cells. Viruses 2018, 10, 557. [Google Scholar] [CrossRef] [Green Version]
- Colpitts, C.C.; Tawar, R.G.; Mailly, L.; Thumann, C.; Heydmann, L.; Durand, S.C.; Xiao, F.; Robinet, E.; Pessaux, P.; Zeisel, M.B.; et al. Humanisation of a claudin-1-specific monoclonal antibody for clinical prevention and cure of HCV infection without escape. Gut 2018, 67, 736–745. [Google Scholar] [CrossRef] [Green Version]
- Denolly, S.; Cosset, F.L. A master regulator of tight junctions involved in hepatitis C virus entry and pathogenesis. Hepatology 2017, 65, 1756–1764. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, J.T.; Rathore, A.P.S.; Soundarajan, G.; St John, A.L. Japanese encephalitis virus neuropenetrance is driven by mast cell chymase. Nat. Commun. 2019, 10, 706. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, Y.; Shirasago, Y.; Kondoh, M.; Suzuki, T.; Wakita, T.; Hanada, K.; Yagi, K.; Fukasawa, M. Monoclonal Antibodies against Occludin Completely Prevented Hepatitis C Virus Infection in a Mouse Model. J. Virol. 2018, 92, e02258-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Lu, X.; Ling, L.; Li, H.; Ou, Y.; Shi, X.; Lu, Y.; Zhang, Y.; Chen, D. Houttuynia cordata polysaccharides ameliorate pneumonia severity and intestinal injury in mice with influenza virus infection. J. Ethnopharmacol. 2018, 218, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Q.; Liu, Y.; Zhang, B.; Liu, H.; Shao, D.D.; Liu, J.B.; Wang, X.; Zhou, L.N.; Hu, W.H.; Ho, W.Z. IL-22 suppresses HSV-2 replication in human cervical epithelial cells. Cytokine 2019, 123, 154776. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, G.; Shao, Q.; Yu, H.; Liu, J.; Li, Y.; Wang, B.; Sang, H.; Li, D.; Bing, A.; Hou, Y.; et al. Tight Junctions, the Key Factor in Virus-Related Disease. Pathogens 2022, 11, 1200. https://doi.org/10.3390/pathogens11101200
Ding G, Shao Q, Yu H, Liu J, Li Y, Wang B, Sang H, Li D, Bing A, Hou Y, et al. Tight Junctions, the Key Factor in Virus-Related Disease. Pathogens. 2022; 11(10):1200. https://doi.org/10.3390/pathogens11101200
Chicago/Turabian StyleDing, Guofei, Qingyuan Shao, Haiyan Yu, Jiaqi Liu, Yingchao Li, Bin Wang, Haotian Sang, Dexin Li, Aiying Bing, Yanmeng Hou, and et al. 2022. "Tight Junctions, the Key Factor in Virus-Related Disease" Pathogens 11, no. 10: 1200. https://doi.org/10.3390/pathogens11101200