
The Role of Growth Factors in the Pathogenesis of Dengue: A Scoping Review

Abstract
:1. Introduction
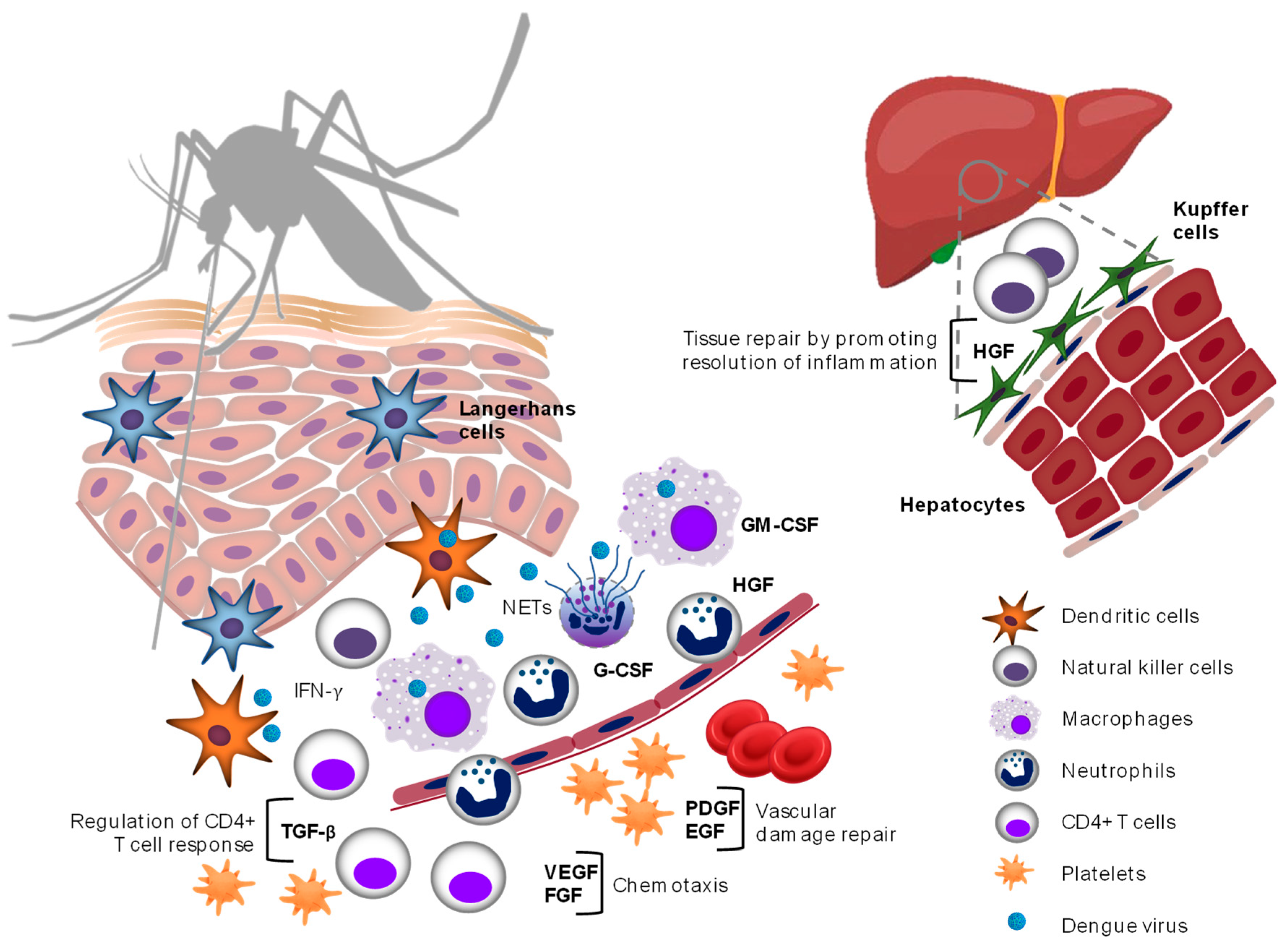
2. Results
2.1. Vascular Endothelial Growth Factor (VEGF)
2.2. Granulocyte–Macrophage Colony-Stimulating Factor (GM-CSF)
2.3. Granulocyte Colony-Stimulating Factor (G-CSF)
2.4. Transforming Growth Factor Beta (TGF-β)
2.5. Fibroblast Growth Factor (FGF)
2.6. Platelet-Derived Growth Factor (PDGF)
2.7. Epidermal Growth Factor (EGF)
2.8. Hepatocyte Growth Factor (HGF)
3. Discussion
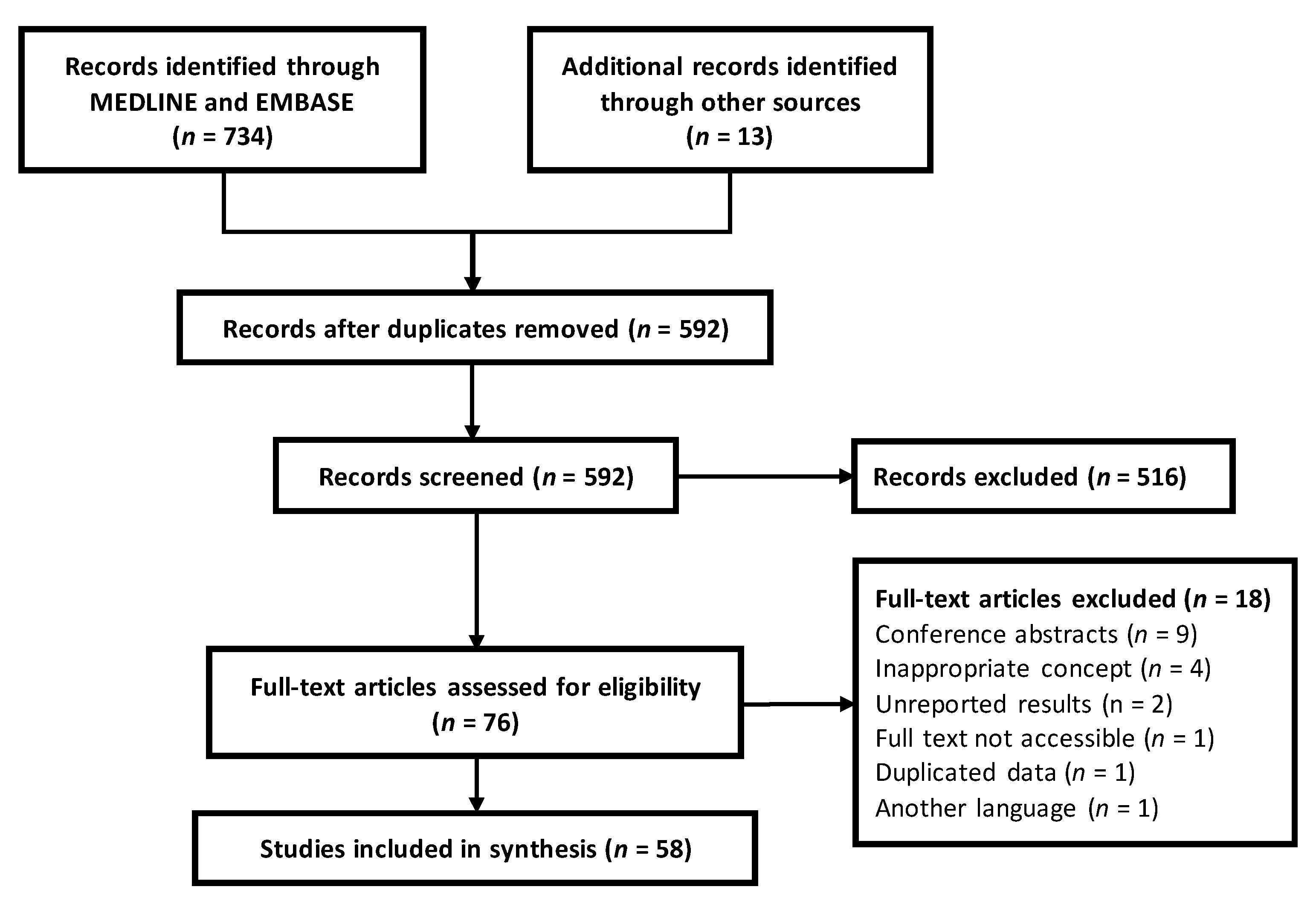
4. Materials and Methods
4.1. Search Strategies
4.2. Elegibilty Criteria
4.3. Study Selection
4.4. Data Extraction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization (WHO). Dengue and Severe Dengue Geneva. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue. (accessed on 12 March 2022).
- Malavige, G.N.; Ogg, G.S. Pathogenesis of vascular leak in dengue virus infection. Immunology 2017, 151, 261–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, P.; Sabeena, S.P.; Varma, M.; Arunkumar, G. Current Understanding of the Pathogenesis of Dengue Virus Infection. Curr. Microbiol. 2021, 78, 17–32. [Google Scholar] [CrossRef]
- Soo, K.M.; Khalid, B.; Ching, S.M.; Tham, C.L.; Basir, R.; Chee, H.Y. Meta-analysis of biomarkers for severe dengue infections. PeerJ 2017, 5, e3589. [Google Scholar] [CrossRef] [Green Version]
- Stone, W.L.; Logan, L.; Varacallo, M. Physiology, Growth Factor. 2021. Available online: https://www.ncbi.nlm.nih.gov/books/NBK442024/ (accessed on 10 May 2022).
- Hondermarck, H.; Bartlett, N.W.; Nurcombe, V. The role of growth factor receptors in viral infections: An opportunity for drug repurposing against emerging viral diseases such as COVID-19? FASEB Bioadv. 2020, 2, 296–303. [Google Scholar] [CrossRef]
- Da Silveira, L.T.C.; Tura, B.; Santos, M. Systematic review of dengue vaccine efficacy. BMC Infect. Dis. 2019, 19, 750. [Google Scholar] [CrossRef] [PubMed]
- Low, J.G.; Ooi, E.E.; Vasudevan, S.G. Current Status of Dengue Therapeutics Research and Development. J. Infect. Dis. 2017, 215 (Suppl. S2), S96–S102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tricco, A.C.; Lillie, E.; Zarin, W.; O’Brien, K.K.; Colquhoun, H.; Levac, D.; Moher, D.; Peters, M.D.J.; Horsley, T.; Weeks, L.; et al. PRISMA Extension for Scoping Reviews (PRISMA-ScR): Checklist and Explanation. Ann. Intern. Med. 2018, 169, 467–473. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Dengue Haemorrhagic Fever: Diagnosis, Treatment, Prevention and Control; World Health Organization: Geneva, Switzerland, 1997; Available online: https://apps.who.int/iris/handle/10665/41988 (accessed on 10 May 2022).
- World Health Organization (WHO). Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control; World Health Organization: Geneva, Switzerland, 2009; Available online: https://apps.who.int/iris/handle/10665/44188 (accessed on 10 May 2022).
- Becquart, P.; Wauquier, N.; Nkoghe, D.; Ndjoyi-Mbiguino, A.; Padilla, C.; Souris, M.; Leroy, E.M. Acute dengue virus 2 infection in Gabonese patients is associated with an early innate immune response, including strong interferon alpha production. BMC Infect. Dis. 2010, 10, 356. [Google Scholar] [CrossRef] [Green Version]
- Conroy, A.L.; Gélvez, M.; Hawkes, M.; Rajwans, N.; Liles, W.C.; Villar-Centeno, L.A.; Kain, K.C. Host biomarkers distinguish dengue from leptospirosis in Colombia: A case-control study. BMC Infect. Dis. 2014, 14, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conroy, A.L.; Gélvez, M.; Hawkes, M.; Rajwans, N.; Tran, V.; Liles, W.C.; Villar-Centeno, L.A.; Kain, K.C. Host biomarkers are associated with progression to dengue haemorrhagic fever: A nested case-control study. Int. J. Infect. Dis. 2015, 40, 45–53. [Google Scholar] [CrossRef]
- Del Moral-Hernández, O.; Martínez-Hernández, N.E.; Mosso-Pani, M.A.; Hernández-Sotelo, D.; Illades-Aguiar, B.; Flores-Alfaro, E.; Antonio-Vejar, V.; Leyva-Vázquez, M.A. Association DENV1 and DENV2 infection with high serum levels of soluble thrombomodulin and VEGF in patients with dengue fever and dengue hemorrhagic fever. Int. J. Clin. Exp. Med. 2014, 7, 370–378. [Google Scholar]
- Furuta, T.; Murao, L.A.; Lan, N.T.; Huy, N.T.; Huong, V.T.; Thuy, T.T.; Tham, V.D.; Nga, C.T.; Ha, T.T.; Ohmoto, Y.; et al. Association of mast cell-derived VEGF and proteases in Dengue shock syndrome. PLoS Negl. Trop. Dis. 2012, 6, e1505. [Google Scholar] [CrossRef] [Green Version]
- García, C.L.; Martínez, D.; Castro, D.; Salgado, D.M.; Narváez, C.; Puerta, L.; Rodríguez, J.A. Anticuerpos IgG4 específicos anti Aedes aegypti como factor protector en niños con dengue grave. Rev. Med. 2019, 27, 11–20. [Google Scholar] [CrossRef]
- Her, Z.; Kam, Y.W.; Gan, V.C.; Lee, B.; Thein, T.L.; Tan, J.J.; Lee, L.K.; Fink, K.; Lye, D.C.; Rénia, L.; et al. Severity of Plasma Leakage Is Associated with High Levels of Interferon γ-Inducible Protein 10, Hepatocyte Growth Factor, Matrix Metalloproteinase 2 (MMP-2), and MMP-9 During Dengue Virus Infection. J. Infect. Dis. 2017, 215, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Kumar, Y.; Liang, C.; Bo, Z.; Rajapakse, J.C.; Ooi, E.E.; Tannenbaum, S.R. Serum Proteome and Cytokine Analysis in a Longitudinal Cohort of Adults with Primary Dengue Infection Reveals Predictive Markers of DHF. PLoS Negl. Trop. Dis. 2012, 6, e1887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, G.K.K.; Gan, S.C.; Zainal, N.; Naidu, K.D.; Amin-Nordin, S.; Khoo, C.S.; Ridzuan, M.I.; Said, R.M.; Mansor, M.M.; Zailani, M.H.; et al. The predictive and diagnostic accuracy of vascular endothelial growth factor and pentraxin-3 in severe dengue. Pathog. Glob. Health 2018, 112, 334–341. [Google Scholar] [CrossRef]
- Mangione, J.N.; Huy, N.T.; Lan, N.T.; Mbanefo, E.C.; Ha, T.T.; Bao, L.Q.; Nga, C.T.; Tuong, V.V.; Dat, T.V.; Thuy, T.T.; et al. The association of cytokines with severe dengue in children. Trop. Med. Health 2014, 42, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Mutiara Koh, S.C.L.; Bachtiar, A.; Hariman, H. The Vascular Endothelium in Patients with Dengue Haemorrhagic Fever. Open Access Maced. J. Med. Sci. 2019, 7, 2221–2225. [Google Scholar] [CrossRef] [Green Version]
- Nanda, J.D.; Jung, C.J.; Satria, R.D.; Jhan, M.K.; Shen, T.J.; Tseng, P.C.; Wang, Y.T.; Ho, T.S.; Lin, C.F. Serum IL-18 Is a Potential Biomarker for Predicting Severe Dengue Disease Progression. J. Immunol. Res. 2021, 2021. [Google Scholar] [CrossRef] [PubMed]
- Orsi, F.A.; Angerami, R.N.; Mazetto, B.M.; Quaino, S.K.; De Paula, E.V.; Annichino-Bizzachi, J.M. Bleeding complications in dengue are not associated with significant changes in the modulators of the endothelial barrier. J. Infect. Dev. Ctries. 2014, 8, 799–803. [Google Scholar] [CrossRef] [Green Version]
- Patra, G.; Saha, B.; Mukhopadhyay, S. Study of serum VEGF levels in patients with severe dengue infection admitted in a tertiary care hospital in Kolkata. J. Med. Virol. 2019, 91, 1873–1876. [Google Scholar] [CrossRef] [PubMed]
- Singla, M.; Kar, M.; Sethi, T.; Kabra, S.K.; Lodha, R.; Chandele, A.; Medigeshi, G.R. Immune Response to Dengue Virus Infection in Pediatric Patients in New Delhi, India—Association of Viremia, Inflammatory Mediators and Monocytes with Disease Severity. PLoS Negl. Trop. Dis. 2016, 10, e0004497. [Google Scholar] [CrossRef] [Green Version]
- Srikiatkhachorn, A.; Ajariyakhajorn, C.; Endy, T.P.; Kalayanarooj, S.; Libraty, D.H.; Green, S.; Ennis, F.A.; Rothman, A.L. Virus-induced decline in soluble vascular endothelial growth receptor 2 is associated with plasma leakage in dengue hemorrhagic fever. J. Virol. 2007, 81, 1592–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, P.; Chakravarti, A.; Aggarwal, S.; Uppal, B.; Bhalla, P. Elevated levels of vascular endothelial growth factor in adults with severe dengue infection. Virus Dis. 2016, 27, 48–54. [Google Scholar] [CrossRef]
- Tramontini Gomes de Sousa Cardozo, F.; Baimukanova, G.; Lanteri, M.C.; Keating, S.M.; Moraes Ferreira, F.; Heitman, J.; Pannuti, C.S.; Pati, S.; Romano, C.M.; Cerdeira Sabino, E. Serum from dengue virus-infected patients with and without plasma leakage differentially affects endothelial cells barrier function in vitro. PLoS ONE 2017, 12, e0178820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, C.S.; Lo, H.W.; Teng, H.C.; Lo, W.C.; Ker, C.G. Elevated levels of plasma VEGF in patients with dengue hemorrhagic fever. FEMS Immunol. Med. Microbiol. 2005, 43, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Van de Weg, C.A.; Pannuti, C.S.; de Araújo, E.S.; van den Ham, H.J.; Andeweg, A.C.; Boas, L.S.; Felix, A.C.; Carvalho, K.I.; de Matos, A.M.; Levi, J.E.; et al. Microbial translocation is associated with extensive immune activation in dengue virus infected patients with severe disease. PLoS Negl. Trop. Dis. 2013, 7, e2236. [Google Scholar] [CrossRef]
- Van de Weg, C.A.; Pannuti, C.S.; van den Ham, H.J.; de Araújo, E.S.; Boas, L.S.; Felix, A.C.; Carvalho, K.I.; Levi, J.E.; Romano, C.M.; Centrone, C.C.; et al. Serum angiopoietin-2 and soluble VEGF receptor 2 are surrogate markers for plasma leakage in patients with acute dengue virus infection. J. Clin. Virol. 2014, 60, 328–335. [Google Scholar] [CrossRef] [Green Version]
- Yong, Y.K.; Tan, H.Y.; Jen, S.H.; Shankar, E.M.; Natkunam, S.K.; Sathar, J.; Manikam, R.; Sekaran, S.D. Aberrant monocyte responses predict and characterize dengue virus infection in individuals with severe disease. J. Transl. Med. 2017, 15, 121. [Google Scholar] [CrossRef]
- Appanna, R.; Wang, S.M.; Ponnampalavanar, S.A.; Lum, L.C.; Sekaran, S.D. Cytokine factors present in dengue patient sera induces alterations of junctional proteins in human endothelial cells. Am. J. Trop. Med. Hyg. 2012, 87, 936–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros, T.A.D.C.; Batista, D.D.O.; Torrentes de Carvalho, A.; Costa Faria, N.R.D.; Barreto-Vieira, D.F.; Jácome, F.C.; Barth, O.M.; Nogueira, R.M.R.; Neves, P.C.D.C.; Matos, D.C.D.S.; et al. Different aspects of platelet evaluation in dengue: Measurement of circulating mediators, ability to interact with the virus, the degree of activation and quantification of intraplatelet protein content. Virus Res. 2019, 260, 163–172. [Google Scholar] [CrossRef]
- Butthep, P.; Chunhakan, S.; Yoksan, S.; Tangnararatchakit, K.; Chuansumrit, A. Alteration of cytokines and chemokines during febrile episodes associated with endothelial cell damage and plasma leakage in dengue hemorrhagic fever. Pediatr. Infect. Dis. J. 2012, 31, e232–e238. [Google Scholar] [CrossRef] [PubMed]
- Chagan-Yasutan, H.; Ndhlovu, L.C.; Lacuesta, T.L.; Kubo, T.; Leano, P.S.; Niki, T.; Oguma, S.; Morita, K.; Chew, G.M.; Barbour, J.D.; et al. Galectin-9 plasma levels reflect adverse hematological and immunological features in acute dengue virus infection. J. Clin. Virol. 2013, 58, 635–640. [Google Scholar] [CrossRef] [Green Version]
- Rathakrishnan, A.; Wang, S.M.; Hu, Y.; Khan, A.M.; Ponnampalavanar, S.; Lum, L.C.; Manikam, R.; Sekaran, S.D. Cytokine expression profile of dengue patients at different phases of illness. PLoS ONE 2012, 7, e52215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathupan, P.; Khongphattanayothin, A.; Srisai, J.; Srikaew, K.; Poovorawan, Y. The role of vascular endothelial growth factor leading to vascular leakage in children with dengue virus infection. Ann. Trop. Paediatr. 2007, 27, 179–184. [Google Scholar] [CrossRef]
- Seet, R.C.; Chow, A.W.; Quek, A.M.; Chan, Y.H.; Lim, E.C. Relationship between circulating vascular endothelial growth factor and its soluble receptors in adults with dengue virus infection: A case-control study. Int. J. Infect. Dis. 2009, 13, e248–e253. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, C.L.; Gélvez, M.; Niederbacher, J. Regulators of endothelial integrity as severity predictors in dengue. Biomedica 2016, 36, 148–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, U.K.; Kalita, J.; Singh, A.P. Role of vascular endothelial growth factor (VEGF) in the neurological manifestations of dengue: A preliminary study. Inflammation 2014, 37, 611–614. [Google Scholar] [CrossRef]
- Yacoub, S.; Lam, P.K.; Vu, L.H.M.; Le, T.L.; Ha, N.T.; Toan, T.T.; Thu Van, N.; Quyen, N.T.H.; Duyen, H.T.L.; Kinh, N.V.; et al. Association of microvascular function and endothelial biomarkers with clinical outcome in dengue: An observational study. J. Infect. Dis. 2016, 214, 697–706. [Google Scholar] [CrossRef] [Green Version]
- Kalita, J.; Chauhan, P.S.; Mani, V.E.; Bhoi, S.K.; Misra, U.K. VEGF and its receptors in dengue virus infection. J. Med. Virol. 2015, 87, 1449–1455. [Google Scholar] [CrossRef]
- Bozza, F.A.; Cruz, O.G.; Zagne, S.M.; Azeredo, E.L.; Nogueira, R.M.; Assis, E.F.; Bozza, P.T.; Kubelka, C.F. Multiplex cytokine profile from dengue patients: MIP-1beta and IFN-gamma as predictive factors for severity. BMC Infect. Dis. 2008, 8, 86. [Google Scholar] [CrossRef]
- Gowri Sankar, S.; Alwin Prem Anand, A. Cytokine IP-10 and GM-CSF are prognostic biomarkers for severity in secondary dengue infection. Hum. Immunol. 2021, 82, 438–445. [Google Scholar] [CrossRef]
- Gowri Sankar, S.; Mowna Sundari, T.; Alwin Prem Anand, A. Emergence of Dengue 4 as Dominant Serotype During 2017 Outbreak in South India and Associated Cytokine Expression Profile. Front. Cell Infect. Microbiol. 2021, 11, 681937. [Google Scholar] [CrossRef]
- Li, H.; Li, Y.; Wen, B.; Zhang, J.; Wang, C.; Song, Z.; Li, S.; Qu, X.; Huang, R.; Liu, W. Dengue virus and Japanese encephalitis virus infection of the central nervous system share similar profiles of cytokine accumulation in cerebrospinal fluid. Cent. Eur. J. Immunol. 2017, 42, 218–222. [Google Scholar] [CrossRef]
- Maneekan, P.; Leaungwutiwong, P.; Misse, D.; Luplertlop, N. T helper (Th) 1 and Th2 cytokine expression profile in dengue and malaria infection using magnetic bead-based bio-plex assay. Southeast Asian J. Trop. Med. Public Health 2013, 44, 31–36. [Google Scholar] [PubMed]
- Oliveira, R.A.D.S.; Cordeiro, M.T.; Moura, P.M.M.F.D.; Baptista Filho, P.N.B.; Braga-Neto, U.D.M.; Marques, E.T.D.A.; Gil, L.H.V.G. Serum cytokine/chemokine profiles in patients with dengue fever (DF) and dengue hemorrhagic fever (FHD) by using protein array. J. Clin. Virol. 2017, 89, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Patro, A.R.K.; Mohanty, S.; Prusty, B.K.; Singh, D.K.; Gaikwad, S.; Saswat, T.; Chattopadhyay, S.; Das, B.K.; Tripathy, R.; Ravindran, B. Cytokine Signature Associated with Disease Severity in Dengue. Viruses 2019, 11, 34. [Google Scholar] [CrossRef] [Green Version]
- Puc, I.; Ho, T.C.; Yen, K.L.; Vats, A.; Tsai, J.J.; Chen, P.L.; Chien, Y.W.; Lo, Y.C.; Perng, G.C. Cytokine Signature of Dengue Patients at Different Severity of the Disease. Int. J. Mol. Sci. 2021, 22, 2879. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.H.; Lin, C.Y.; Chang, K.; Urbina, A.N.; Assavalapsakul, W.; Thitithanyanont, A.; Lu, P.L.; Chen, Y.H.; Wang, S.F. A clinical and epidemiological survey of the largest dengue outbreak in Southern Taiwan in 2015. Int. J. Infect. Dis. 2019, 88, 88–99. [Google Scholar] [CrossRef] [Green Version]
- Meena, A.A.; Murugesan, A.; Sopnajothi, S.; Yong, Y.K.; Ganesh, P.S.; Vimali, I.J.; Vignesh, R.; Elanchezhiyan, M.; Kannan, M.; Dash, A.P.; et al. Increase of Plasma TNF-α Is Associated with Decreased Levels of Blood Platelets in Clinical Dengue Infection. Viral Immunol. 2020, 33, 54–60. [Google Scholar] [CrossRef]
- Mendonça, V.R.R.; Andrade, B.B.; Souza, L.C.L.; Magalhães, B.M.L.; Mourão, M.P.G.; Lacerda, M.V.G.; Barral-Netto, M. Unravelling the patterns of host immune responses in Plasmodium vivax malaria and dengue co-infection. Malar J. 2015, 14, 315. [Google Scholar] [CrossRef]
- Cui, L.; Lee, Y.H.; Thein, T.L.; Fang, J.; Pang, J.; Ooi, E.E.; Leo, Y.S.; Ong, C.N.; Tannenbaum, S.R. Serum Metabolomics Reveals Serotonin as a Predictor of Severe Dengue in the Early Phase of Dengue Fever. PLoS Negl. Trop. Dis. 2016, 10, e0004607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halsey, E.S.; Baldeviano, G.C.; Edgel, K.A.; Vilcarromero, S.; Sihuincha, M.; Lescano, A.G. Symptoms and Immune Markers in Plasmodium/Dengue Virus Co-infection Compared with Mono-infection with Either in Peru. PLoS Negl. Trop. Dis. 2016, 10, e0004646. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, R.; Elbishbishi, E.A.; Chaturvedi, U.C.; Nagar, R.; Mustafa, A.S. Profile of transforming growth factor-beta 1 in patients with dengue haemorrhagic fever. Int. J. Exp. Pathol. 1999, 80, 143–149. [Google Scholar] [CrossRef]
- Azeredo, E.L.; Zagne, S.M.; Alvarenga, A.R.; Nogueira, R.M.; Kubelka, C.F.; de Oliveira-Pinto, L.M. Activated peripheral lymphocytes with increased expression of cell adhesion molecules and cytotoxic markers are associated with dengue fever disease. Mem. Inst. Oswaldo Cruz 2006, 101, 437–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, N.; Jain, A.; Garg, R.K.; Kumar, R.; Agrawal, O.P.; Lakshmana Rao, P.V. Serum levels of IL-8, IFNγ, IL-10, and TGF β and their gene expression levels in severe and non-severe cases of dengue virus infection. Arch Virol. 2015, 160, 1463–1475. [Google Scholar] [CrossRef]
- Tillu, H.; Tripathy, A.S.; Reshmi, P.V.; Cecilia, D. Altered profile of regulatory T cells and associated cytokines in mild and moderate dengue. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 453–461. [Google Scholar] [CrossRef]
- Patra, G.; Mallik, S.; Saha, B.; Mukhopadhyay, S. Assessment of chemokine and cytokine signatures in patients with dengue infection: A hospital-based study in Kolkata, India. Acta Trop. 2019, 190, 73–79. [Google Scholar] [CrossRef]
- Jayaratne, H.E.; Wijeratne, D.; Fernando, S.; Kamaladasa, A.; Gomes, L.; Wijewickrama, A.; Ogg, G.S.; Malavige, G.N. Regulatory T-cells in acute dengue viral infection. Immunology 2018, 154, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Laur, F.; Murgue, B.; Deparis, X.; Roche, C.; Cassar, O.; Chungue, E. Plasma levels of tumour necrosis factor alpha and transforming growth factor beta-1 in children with dengue 2 virus infection in French Polynesia. Trans. R Soc. Trop. Med. Hyg. 1998, 92, 654–656. [Google Scholar] [CrossRef]
- Djamiatun, K.; Faradz, S.M.; Setiati, T.E.; Netea, M.G.; van der Ven, A.J.; Dolmans, W.M. Increase of plasminogen activator inhibitor-1 and decrease of transforming growth factor-b1 in children with dengue haemorrhagic fever in Indonesia. J. Trop. Pediatr. 2011, 57, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Malavige, G.N.; Huang, L.C.; Salimi, M.; Gomes, L.; Jayaratne, S.D.; Ogg, G.S. Cellular and cytokine correlates of severe dengue infection. PLoS ONE 2012, 7, e50387. [Google Scholar] [CrossRef] [Green Version]
- Harenberg, A.; de Montfort, A.; Jantet-Blaudez, F.; Bonaparte, M.; Boudet, F.; Saville, M.; Jackson, N.; Guy, B. Cytokine Profile of Children Hospitalized with Virologically-Confirmed Dengue during Two Phase III Vaccine Efficacy Trials. PLoS Negl. Trop. Dis. 2016, 10, e0004830. [Google Scholar] [CrossRef]
- Dhenni, R.; Yohan, B.; Alisjahbana, B.; Lucanus, A.; Riswari, S.F.; Megawati, D.; Haryanto, S.; Gampamole, D.; Hayati, R.F.; Sari, K.; et al. Comparative cytokine profiling identifies common and unique serum cytokine responses in acute chikungunya and dengue virus infection. BMC Infect. Dis. 2021, 21, 639. [Google Scholar] [CrossRef]
- Voraphani, N.; Theamboonlers, A.; Khongphatthanayothin, A.; Srisai, C.; Poovorawan, Y. Increased level of hepatocyte growth factor in children with dengue virus infection. Ann. Trop. Paediatr. 2010, 30, 213–218. [Google Scholar] [CrossRef]
- Imamura, R.; Matsumoto, K. Hepatocyte growth factor in physiology and infectious diseases. Cytokine 2017, 98, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Lateef, Z.; Wise, L.M. Exploitation of receptor tyrosine kinases by viral-encoded growth factors. Growth Factors 2018, 36, 118–140. [Google Scholar] [CrossRef] [PubMed]
- Melincovici, C.S.; Boşca, A.B.; Şuşman, S.; Mărginean, M.; Mihu, C.; Istrate, M.; Moldovan, I.M.; Roman, A.L.; Mihu, C.M. Vascular endothelial growth factor (VEGF)—Key factor in normal and pathological angiogenesis. Rom. J. Morphol. Embryol. 2018, 59, 455–467. [Google Scholar]
- Bates, D.O.; Beazley-Long, N.; Benest, A.V.; Ye, X.; Ved, N.; Hulse, R.P.; Barratt, S.; Machado, M.J.; Donaldson, L.F.; Harper, S.J.; et al. Physiological Role of Vascular Endothelial Growth Factors as Homeostatic Regulators. Compr. Physiol. 2018, 8, 955–979. [Google Scholar] [CrossRef]
- De Azeredo, E.L.; Monteiro, R.Q.; de-Oliveira Pinto, L.M. Thrombocytopenia in Dengue: Interrelationship between Virus and the Imbalance between Coagulation and Fibrinolysis and Inflammatory Mediators. Mediat. Inflamm. 2015, 2015, 313842. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, E.; Habib, A.; Lebret, M.; Yang, R.; Levy-Toledano, S.; Tobelem, G. Thrombin induces angiogenesis and vascular endothelial growth factor expression in human endothelial cells: Possible relevance to HIF-1alpha. J. Thromb. Haemost. 2003, 1, 1096–1102. [Google Scholar] [CrossRef]
- Bode, M.; Mackman, N. Regulation of tissue factor gene expression in monocytes and endothelial cells: Thromboxane A2 as a new player. Vascul. Pharmacol. 2014, 62, 57–62. [Google Scholar] [CrossRef]
- Leal de Azeredo, E.; Solorzano, V.E.; de Oliveira, D.B.; Marinho, C.F.; de Souza, L.J.; da Cunha, R.V.; Damasco, P.V.; Kubelka, C.F.; de-Oliveira-Pinto, L.M. Increased circulating procoagulant and anticoagulant factors as TF and TFPI according to severity or infecting serotypes in human dengue infection. Microbes Infect. 2017, 19, 62–68. [Google Scholar] [CrossRef]
- Alkharsah, K.R. VEGF Upregulation in Viral Infections and Its Possible Therapeutic Implications. Int. J. Mol. Sci. 2018, 19, 1642. [Google Scholar] [CrossRef] [Green Version]
- Olsson, A.K.; Dimberg, A.; Kreuger, J.; Claesson-Welsh, L. VEGF receptor signalling—In control of vascular function. Nat. Rev. Mol. Cell. Biol. 2006, 7, 359–371. [Google Scholar] [CrossRef]
- Hamilton, J.A. GM-CSF in inflammation. J Exp Med. 2020, 217, e20190945. [Google Scholar] [CrossRef] [Green Version]
- Martin, K.R.; Wong, H.L.; Witko-Sarsat, V.; Wicks, I.P. G-CSF—A double edge sword in neutrophil mediated immunity. Semin. Immunol. 2021, 54, 101516. [Google Scholar] [CrossRef]
- Sun, L.; Rautela, J.; Delconte, R.B.; Souza-Fonseca-Guimaraes, F.; Carrington, E.M.; Schenk, R.L.; Herold, M.J.; Huntington, N.D.; Lew, A.M.; Xu, Y.; et al. GM-CSF Quantity Has a Selective Effect on Granulocytic vs. Monocytic Myeloid Development and Function. Front Immunol. 2018, 9, 1922. [Google Scholar] [CrossRef]
- Wu, M.F.; Chen, S.T.; Yang, A.H.; Lin, W.W.; Lin, Y.L.; Chen, N.J.; Tsai, I.S.; Li, L.; Hsieh, S.L. CLEC5A is critical for dengue virus-induced inflammasome activation in human macrophages. Blood 2013, 121, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.F.; Chen, S.T.; Hsieh, S.L. Distinct regulation of dengue virus-induced inflammasome activation in human macrophage subsets. J. Biomed. Sci. 2013, 20, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opasawatchai, A.; Amornsupawat, P.; Jiravejchakul, N.; Chan-In, W.; Spoerk, N.J.; Manopwisedjaroen, K.; Singhasivanon, P.; Yingtaweesak, T.; Suraamornkul, S.; Mongkolsapaya, J.; et al. Neutrophil Activation and Early Features of NET Formation Are Associated With Dengue Virus Infection in Human. Front. Immunol. 2018, 9, 3007. [Google Scholar] [CrossRef] [Green Version]
- Hinck, A.P. Structural studies of the TGF-βs and their receptors—Insights into evolution of the TGF-β superfamily. FEBS Lett. 2012, 586, 1860–1870. [Google Scholar] [CrossRef]
- Xu, X.; Zheng, L.; Yuan, Q.; Zhen, G.; Crane, J.L.; Zhou, X.; Cao, X. Transforming growth factor-β in stem cells and tissue homeostasis. Bone Res. 2018, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Travis, M.A.; Sheppard, D. TGF-β activation and function in immunity. Annu. Rev. Immunol. 2014, 32, 51–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanjabi, S.; Oh, S.A.; Li, M.O. Regulation of the Immune Response by TGF-β: From Conception to Autoimmunity and Infection. Cold Spring Harb. Perspect. Biol. 2017, 9, a022236. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Vargas, L.A.; Hernández-Flores, K.G.; Thomas-Dupont, P.; Izaguirre-Hernández, I.Y.; Sánchez-Marce, E.E.; Remes-Ruiz, R.; Fonseca-Coronado, S.; Hernández-Romano, P.A.; Flores-Collins, M.E.; Vivanco-Cid, H. Characterization of the IL-17 and CD4+ Th17 Cells in the Clinical Course of Dengue Virus Infections. Viruses 2020, 12, 1435. [Google Scholar] [CrossRef]
- Kang, Y.H.; Brummel, S.E.; Lee, C.H. Differential effects of transforming growth factor-beta 1 on lipopolysaccharide induction of endothelial adhesion molecules. Shock 1996, 6, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Karolczak, K.; Watala, C. Blood Platelets as an Important but Underrated Circulating Source of TGFβ. Int. J. Mol. Sci. 2021, 22, 4492. [Google Scholar] [CrossRef]
- Teixeira, G.S.; Andrade, A.A.; Torres, L.R.; Couto-Lima, D.; Moreira, O.C.; Abreu, R.S.; Waghabi, M.C.; de Souza, E.M. Suppression of TGF-β/Smad2 signaling by GW788388 enhances DENV-2 clearance in macrophages. J. Med. Virol. 2022, 94, 4359–4368. [Google Scholar] [CrossRef]
- Chen, R.F.; Wang, L.; Cheng, J.T.; Chuang, H.; Chang, J.C.; Liu, J.W.; Lin, I.C.; Yang, K.D. Combination of CTLA-4 and TGFbeta1 gene polymorphisms associated with dengue hemorrhagic fever and virus load in a dengue-2 outbreak. Clin. Immunol. 2009, 131, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, Y.; He, D. Diverse effects of platelet-derived growth factor-BB on cell signaling pathways. Cytokine 2019, 113, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Berlanga-Acosta, J.; Gavilondo-Cowley, J.; López-Saura, P.; González-López, T.; Castro-Santana, M.D.; López-Mola, E.; Guillén-Nieto, G.; Herrera-Martinez, L. Epidermal growth factor in clinical practice—A review of its biological actions, clinical indications and safety implications. Int. Wound J. 2009, 6, 331–346. [Google Scholar] [CrossRef] [PubMed]
- Martínez, C.E.; Smith, P.C.; Palma Alvarado, V.A. The influence of platelet-derived products on angiogenesis and tissue repair: A concise update. Front. Physiol. 2015, 6, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.; Sakai, K.; Nakamura, T.; Matsumoto, K. Hepatocyte growth factor twenty years on: Much more than a growth factor. J. Gastroenterol. Hepatol. 2011, 26 (Suppl. S1), 188–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilangumaran, S.; Villalobos-Hernandez, A.; Bobbala, D.; Ramanathan, S. The hepatocyte growth factor (HGF)-MET receptor tyrosine kinase signaling pathway: Diverse roles in modulating immune cell functions. Cytokine 2016, 82, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Galimi, F.; Cottone, E.; Vigna, E.; Arena, N.; Boccaccio, C.; Giordano, S.; Naldini, L.; Comoglio, P.M. Hepatocyte growth factor is a regulator of monocyte-macrophage function. J. Immunol. 2001, 166, 1241–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hübel, J.; Hieronymus, T. HGF/Met-Signaling Contributes to Immune Regulation by Modulating Tolerogenic and Motogenic Properties of Dendritic Cells. Biomedicines 2015, 3, 138–148. [Google Scholar] [CrossRef] [Green Version]
- Coudriet, G.M.; He, J.; Trucco, M.; Mars, W.M.; Piganelli, J.D. Hepatocyte growth factor modulates interleukin-6 production in bone marrow derived macrophages: Implications for inflammatory mediated diseases. PLoS ONE 2010, 5, e15384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutella, S.; Bonanno, G.; Procoli, A.; Mariotti, A.; de Ritis, D.G.; Curti, A.; Danese, S.; Pessina, G.; Pandolfi, S.; Natoni, F.; et al. Hepatocyte growth factor favors monocyte differentiation into regulatory interleukin (IL)-10++IL-12low/neg accessory cells with dendritic-cell features. Blood 2006, 108, 218–227. [Google Scholar] [CrossRef]
- Molnarfi, N.; Benkhoucha, M.; Juillard, C.; Bjarnadóttir, K.; Lalive, P.H. The neurotrophic hepatocyte growth factor induces protolerogenic human dendritic cells. J. Neuroimmunol. 2014, 267, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Shiota, G.; Okano, J.; Kawasaki, H.; Kawamoto, T.; Nakamura, T. Serum hepatocyte growth factor levels in liver diseases: Clinical implications. Hepatology 1995, 21, 106–112. [Google Scholar] [CrossRef]
- Wang, X.J.; Wei, H.X.; Jiang, S.C.; He, C.; Xu, X.J.; Peng, H.J. Evaluation of aminotransferase abnormality in dengue patients: A meta analysis. Acta Trop. 2016, 156, 130–136. [Google Scholar] [CrossRef]
- Almeida, R.S.; Ferreira, M.L.B.; Sonon, P.; Cordeiro, M.T.; Sadissou, I.; Diniz, G.T.N.; Militão-Albuquerque, M.F.P.; Franca, R.F.O.; Donadi, E.A.; Lucena-Silva, N. Cytokines and Soluble HLA-G Levels in the Acute and Recovery Phases of Arbovirus-Infected Brazilian Patients Exhibiting Neurological Complications. Front. Immunol. 2021, 12, 582935. [Google Scholar] [CrossRef] [PubMed]
- Kam, Y.W.; Leite, J.A.; Lum, F.M.; Tan, J.J.L.; Lee, B.; Judice, C.C.; Teixeira, D.A.T.; Andreata-Santos, R.; Vinolo, M.A.; Angerami, R.; et al. Specific Biomarkers Associated with Neurological Complications and Congenital Central Nervous System Abnormalities from Zika Virus-Infected Patients in Brazil. J. Infect. Dis. 2017, 216, 172–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ornitz, D.M.; Itoh, N. The Fibroblast Growth Factor signaling pathway. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 215–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Presta, M.; Andrés, G.; Leali, D.; Dell’Era, P.; Ronca, R. Inflammatory cells and chemokines sustain FGF2-induced angiogenesis. Eur. Cytokine Netw. 2009, 20, 39–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortese, M.; Kumar, A.; Matula, P.; Kaderali, L.; Scaturro, P.; Erfle, H.; Acosta, E.G.; Buehler, S.; Ruggieri, A.; Chatel-Chaix, L.; et al. Reciprocal Effects of Fibroblast Growth Factor Receptor Signaling on Dengue Virus Replication and Virion Production. Cell Rep. 2019, 27, 2579–2592.e6. [Google Scholar] [CrossRef] [Green Version]
- Norshidah, H.; Vignesh, R.; Lai, N.S. Updates on Dengue Vaccine and Antiviral: Where Are We Heading? Molecules 2021, 26, 6768. [Google Scholar] [CrossRef]
- Zakaria, M.K.; Carletti, T.; Marcello, A. Cellular Targets for the Treatment of Flavivirus Infections. Front. Cell Infect. Microbiol. 2018, 8, 398. [Google Scholar] [CrossRef]
- De Wispelaere, M.; LaCroix, A.J.; Yang, P.L. The small molecules AZD0530 and dasatinib inhibit dengue virus RNA replication via Fyn kinase. J. Virol. 2013, 87, 7367–7381. [Google Scholar] [CrossRef] [Green Version]
- Bekerman, E.; Neveu, G.; Shulla, A.; Brannan, J.; Pu, S.Y.; Wang, S.; Xiao, F.; Barouch-Bentov, R.; Bakken, R.R.; Mateo, R.; et al. Anticancer kinase inhibitors impair intracellular viral trafficking and exert broad-spectrum antiviral effects. J. Clin. Investig. 2017, 127, 1338–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duran, A.; Valero, N.; Mosquera, J.; Fuenmayor, E.; Alvarez-Mon, M. Gefitinib and pyrrolidine dithiocarbamate decrease viral replication and cytokine production in dengue virus infected human monocyte cultures. Life Sci. 2017, 191, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Wu, W.; Wan, Y.; Li, L.; Zhang, J.; Chen, X.; Liu, S.; Yao, X. Brivanib alaninate inhibited dengue virus proliferation through VEGFR2/AMPK pathway. Pharmacol. Res. 2021, 170, 105721. [Google Scholar] [CrossRef]
- Chuang, F.K.; Liao, C.L.; Hu, M.K.; Chiu, Y.L.; Lee, A.R.; Huang, S.M.; Chiu, Y.L.; Tsai, P.L.; Su, B.C.; Chang, T.H.; et al. Antiviral Activity of Compound L3 against Dengue and Zika Viruses in Vitro and in Vivo. Int. J. Mol Sci. 2020, 21, 4050. [Google Scholar] [CrossRef] [PubMed]
- Branche, E.; Tang, W.W.; Viramontes, K.M.; Young, M.P.; Sheets, N.; Joo, Y.; Nguyen, A.T.; Shresta, S. Synergism between the tyrosine kinase inhibitor sunitinib and Anti-TNF antibody protects against lethal dengue infection. Antivir. Res. 2018, 158, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Peters, M.D.J.; Godfrey, C.; McInerney, P.; Munn, Z.; Tricco, A.C.; Khalil, H. Chapter 11: Scoping reviews. In JBI Manual for Evidence Synthesis; JBI: Adelaide, Australia, 2020; Available online: https://synthesismanual.jbi.global (accessed on 12 March 2022).
- Fiestas Solorzano, V.E.; de Lima, R.C.; de Azeredo, E.L. Role of Growth Factors in the Pathogenesis of Dengue: Scoping Review Protocol. Available online: https://doi.org/10.17605/OSF.IO/JEZV5 (accessed on 10 March 2022).



{kind=link}
{kind=link}
| First Author | Location | Study Design | Age Group | Dengue Patients | Measurement of GFs * | Sample | Laboratory Technique | |
|---|---|---|---|---|---|---|---|---|
| Becquart et al., 2010 [12] | GA | Cross-sectional | Adults | DF: 36 | VEGF: GM-CSF: G-CSF: PDGF: HGF: FGF: | ↑ DF compared with HC (a, b, c) ↑ DF compared with HC (a, b, c) ↑ DF compared with HC (a, b, c) no difference between DF and HC no difference between DF and HC no difference between DF and HC | Plasma | Luminex |
| Conroy et al., 2014 [13] | CO | Case-control | Adults and children | Dengue: 113 | VEGF: | ↑ dengue patients compared with HC (a) | Serum | ELISA |
| Conroy et al., 2015 [14] | CO | Case-control | Adults and children | DF: 65 DHF: 46 | VEGF: | no difference between DF and DHF (a) | Serum | ELISA |
| Del Moral-Hernández et al., 2014 [15] | MX | Cross-sectional | Adults | DF: 70 DHF: 80 | VEGF: | ↑ DHF and DF compared to HC | Serum | ELISA |
| Furuta et al., 2012 [16] | VN | Cohort | Children | DF: 19 DHF: 84 | VEGF: | ↑ DHF compared with DF and HC | Plasma | ELISA |
| Garcia et al., 2019 [17] | CO | Cross-sectional | Children | DwWS: 15 SD: 16 | VEGF: | ↑ SD and DwWS compared with HC | Plasma | ELISA |
| Her et al., 2017 [18] | SG | Cohort | Adults | Non-leakage: 30 Mild leakage: 30 Significative leakage: 30 | VEGF: TGF-β: PDGF: EGF: HGF: | ↑ dengue patients compared with HC (a, b, c) ↑ TGF-β was associated with plasma leakage ↓ PDGF was associated with thrombocytopenia ↑ dengue patients compared with HC (a, b, c) ↑ dengue patients with significative leakage (b) | Plasma | Luminex |
| Kumar et al., 2012 [19] | SG | Cohort | Adults | DF: 44 DHF: 18 | VEGF: G-CSF: PDGF: FGF: | ↑ dengue patients compared with HC, DF > DHF (a) ↑ dengue patients compared with HC, DF > DHF (a) ↑ dengue patients compared with HC, DF > DHF (a) ↑ dengue patients compared with HC (a, b, c) | Serum | Luminex |
| Low et al., 2018 [20] | MY | Cohort | Adults | Nonsevere: 53 Severe: 29 | VEGF: | ↑ severe compared with nonsevere dengue (a) | Plasma | ELISA |
| Mangione et al., 2014 [21] | VN | Case-control | Children | DF: 6, DHF: 18, DSS: 33 | VEGF: | ↑ DSS compared with OFI no difference between DF, DHF and HC | Plasma | Luminex |
| Mutiara et al., 2019 [22] | ID | Cross-sectional | Adults and children | DHF: 50 | VEGF: | ↑ DHF compared with HC (a, b, c) | Plasma | ELISA |
| Nanda et al., 2021 [23] | TW | Cross-sectional | Adults | Nonsevere: 10 Severe: 20 | VEGF: | ↑ severe dengue with comorbidity compared to nonsevere and severe dengue without comorbidity | Plasma | Luminex |
| Orsi et al., 2014 [24] | BR | Cross-sectional | Adults | DF: 59 DHF: 10 | VEGF: | ↑ DHF compared with DF | Plasma | ELISA |
| Patra et al., 2019 [25] | IN | Cross-sectional | Adults | DwoWS: 54, DwWS: 25, SD: 19 | VEGF: | ↑ SD and DwWS compared with DwoWS and HC (b) | Serum | ELISA |
| Singla et al., 2016 [26] | IN | Cohort | Children | DwoWS: 21 DwWS: 30, SD: 46 | VEGF: | ↑ SD compared with DwWS (b) | Plasma | Luminex |
| Srikiatkhachorn et al., 2007 [27] | TH | Cohort | Children | DF: 22 DHF: 23 | VEGF: | ↑ DHF compared with DF (b) | Plasma | ELISA |
| Thakur et al., 2016 [28] | IN | Cross-sectional | Adults | Nonsevere: 58 Severe: 48 | VEGF: | ↑ severe compared with nonsevere dengue and HC (b, c) | Serum | ELISA |
| Tramontini et al., 2017 [29] | BR | Cross-sectional | Adults and children | Non-leakage: 29 Leakage: 28 | VEGF: GM-CSF: G-CSF: EGF: FGF: | ↑ leakage group compared with HC no difference between leakage, non-leakage and HC no difference between leakage, non-leakage and HC ↓ leakage group compared to non-leakage and HC ↓ dengue patients compared with HC | Serum | Luminex |
| Tseng et al., 2005 [30] | TW | Cross-sectional | Adults | DF: 39 DHF: 14 | VEGF: | ↑ DHF compared to DF and HC | Plasma | ELISA |
| van de Weg et al., 2013 [31] | BR | Cross-sectional | Adults and children | DwoWS: 50 DwWS: 49, SD: 33 | VEGF: G-CSF: EGF: HGF: | ↑ SD compared with DwWS and DwoWS ↑ SD compared with DwWS and DwoWS ↓ SD compared with DwWS and DwoWS ↑ SD compared with DwWS and DwoWS | Serum | Luminex |
| van de Weg et al., 2014 [32] | BR | Cross-sectional | Adults and children | Non-leakage: 56 Leakage: 49 | VEGF: | ↑ leakage and non-leakage groups compared with HC | Serum | ELISA |
| Yong et al., 2017 [33] | MY | Cohort | Adults | DwoWS: 43 DwWS: 92, SD: 6 | VEGF: HGF: | ↑ SD compared with DwWS and DwoWS (a, b) ↑ SD and DwWS compared with DwoWS (a) | Plasma | Luminex |
| Appanna et al., 2012 [34] | MY | Cohort | Adults | DF: 13 DHF: 29 | VEGF: G-CSF: PDGF: FGF: | ↓ DHF compared with HC (b) ↓ DHF and DF compared with HC (b, c) ↓ DHF compared with HC (b) ↓ DHF compared with HC (b) | Serum | Luminex |
| Barros et al., 2019 [35] | BR | Cross-sectional | Adults | DwoWS:53 DwWS:31 | VEGF: TGF-β: PDGF: HGF: | ↓ DwWS compared with DwoWS and HC ↓ DwWS compared with DwoWS and HC (intraplatelet) ↓ DwWS compared with DwoWS and HC ↑ DwWS compared with DwoWS and HC (intraplatelet) | Serum | Luminex |
| Butthep et al., 2012 [36] | TH | Cohort | Children | DF: 51 DHF: 196 | VEGF: EGF: | ↓ DHF compared with DF and OFI (a, b) ↓ DHF compared with DF and OFI (b) | Not specified | Chemiluminescent immunoassay |
| Chagan-Yasutan et al., 2013 [37] | PH | Cohort | Adults | Dengue: 65 | VEGF: GM-CSF: EGF: | ↓ dengue patients compared with HC (b, c) ↑ dengue patients compared with HC (b) ↓ dengue patients compared with HC (b, c) | Plasma | Luminex |
| Rathakrishnan et al., 2012 [38] | MY | Cohort | Adults | DwoWS: 11 DwWS: 29, SD: 4 | VEGF: GM-CSF: G-CSF: PDGF: FGF: | ↓ DwWS compared with DwoWS and HC (a, b) ↑ DwWS compared with HC (b) ↓ DwoWS and DwWS compared with HC (a, b, c) ↓ DwWS compared with DwoWS and HC (a, b, c) ↓ DwoWS and DwWS compared with HC (a, b, c) | Serum | Luminex |
| Sathupan et al., 2007 [39] | TH | Cohort | Children | DF: 15 DHF: 26 | VEGF: | ↓ DHF compared with DF and HC (b) | Serum & plasma | ELISA |
| Seet et al., 2009 [40] | SG | Case-control | Adults | DF: 34 DHF: 26 | VEGF: | ↓ DHF and DF compared with HC and OFI | Serum | ELISA |
| Figueroa et al., 2016 [41] | CO | Case-control | Adults and children | DF: 30 DHF: 30 | VEGF: | no difference between DF and DHF (a) | Serum | ELISA |
| Misra et al., 2014 [42] | IN | Cross-sectional | Adults | DF: 12 DHF: 9 | VEGF: | no difference between DHF and DF | Serum | ELISA |
| Yacoub et al., 2016 [43] | VN | Cohort | Adults and children | Non-leakage: 33 Leakage: 37 | VEGF: | no difference between leakage and non-leakage group | Plasma | Luminex |
| Kalita et al., 2015 [44] | IN | Cross-sectional | Adults | DF: 20 DHF: 7 | VEGF: | no difference between dengue patients and HC | Serum | ELISA |
| Bozza et al., 2008 [45] | BR | Cross-sectional | Adults | Mild: 20 Severe: 39 | GM-CSF: | ↑ severe compared to mild dengue | Plasma | Luminex |
| Gowri et al., 2021 [46] | IN | Cross-sectional | Adults and children | DF: 21, DHF: 35, DSS: 20 | GM-CSF: | ↑ dengue patients compared with HC (b) ↑ secondary compared to primary infection | Plasma | ELISA |
| Gowri S et al., 2021 [47] | IN | Cross-sectional | Adults and children | DF: 21, DHF: 35, DSS: 20 | GM-CSF: | ↑ DSS compared with DHF and DF (DENV-2) | Plasma | ELISA |
| Li et al., 2017 [48] | CN | Cross-sectional | Children | Dengue encephalitis: 29 | GM-CSF: | ↑ dengue encephalitis compared with HC | Serum & CSF | Luminex |
| Maneekan et al., 2013 [49] | TH | Cross-sectional | Not reported | Dengue: 20 | GM-CSF: | ↑ dengue patients compared to HC | Serum | Luminex |
| Oliveira et al., 2017 [50] | BR | Cohort | Adults and children | DF: 25 DHF: 19 | GM-CSF: G-CSF: | ↑ dengue patients compared with HC ↑ dengue patients compared with HC | Serum | Cytokine antibody array |
| Patro et al., 2019 [51] | IN | Cross-sectional | Adults | Nonsevere: 73 Severe: 12 | GM-CSF: G-CSF: FGF: | ↑ severe compared with nonsevere dengue and HC ↑ severe compared with nonsevere dengue ↑ severe compared with nonsevere dengue | Plasma | Luminex |
| Puc et al., 2021 [52] | TW | Cross-sectional | Adults | DwoWS: 128, DwWS: 103, SD: 53 | GM-CSF: | ↑ SD compared with DwWS, DwoWS and HC (b, c) | Plasma | Cytometric bead array |
| Wang et al., 2019 [53] | TW | Cross-sectional | Adults | DF: 40 DHF: 20 | GM-CSF: | ↑ DHF compared with DF and HC | Plasma | Luminex |
| Meena et al., 2020 [54] | IN | Cross-sectional | Adults and children | not specified | GM-CSF: | no difference between dengue patients and HC | Serum | Luminex |
| Mendonça et al., 2015 [55] | BR | Cross-sectional | Adults | Dengue: 30 | GM-CSF: G-CSF: | no difference between dengue, malaria and coinfection no difference between dengue, malaria and coinfection | Plasma | Luminex |
| Cui et al., 2016 [56] | SG | Cross-sectional | Adults | DF: 81 DHF: 81 | G-CSF: FGF: | ↑ DHF compared with DF ↑ DHF compared with DF | Serum | Luminex |
| Halsey et al., 2016 [57] | PE | Cross-sectional | Adults and children | Dengue: 51 | G-CSF: TGF-β: | ↓ dengue and coinfection compared to malaria ↑ dengue compared with malaria or coinfection | Serum | Luminex |
| Agarwal et al., 1999 [58] | IN | Cross-sectional | Adults and children | DF: 18 DHF: 61 | TGF-β | ↑ dengue patients compared to HC ↑ DHF compared with DF (b) | Serum | ELISA |
| Azeredo et al., 2006 [59] | BR | Cross-sectional | Adults | Mild: 31 Severe: 19 | TGF-β | ↑ severe compared with mild dengue and HC (a, b, c) | Plasma | ELISA |
| Pandey et al., 2015 [60] | IN | Cross-sectional | Adults and children | DwoWS: 37, DwWS: 53, SD: 121 | TGF-β: | ↑ dengue patients compared with HC ↑ SD and DwWS compared with DwoWS (b) | Plasma | ELISA |
| Tillu et al., 2016 [61] | IN | Cross-sectional | Adults and children | DwoWS: 31 DwWS: 20 | TGF-β | ↑ DwWS compared with DwoWS and HC | Plasma | Luminex |
| Patra et al., 2019 [62] | IN | Cross-sectional | Not reported | DwoWS: 32, DwWS: 21, SD: 9 | TGF-β: | ↑ SD and DwWS compared with DwoWS and HC (b) | Serum | ELISA |
| Jayaratne et al., 2018 [63] | LK | Cohort | Adults | DF: 27 DHF: 26 | TGF-β: | ↑ DHF compared to DF but decrease in both compared with HC | Plasma | ELISA |
| Laur et al., 1998 [64] | PF | Cohort | Children | DF: 33 DHF: 17 | TGF-β: | ↑ DHF compared with DF and HC (a) | Plasma | ELISA |
| Djamiatun et al., 2011 [65] | ID | Cohort | Children | DHF: 71 | TGF-β: | ↓ DHF compared with HC (b) | Plasma | ELISA |
| Malavige et al., 2012 [66] | LK | Cross-sectional | Adults | DHF: 112 | TGF-β | no difference between DHF with shock and without shock | Plasma | Luminex |
| Harenberg et al., 2016 [67] | Multi-center | Clinical trial | Children | Nonsevere: 154 Severe: 52 | EGF: FGF: | ↓ severe dengue compared with nonsevere dengue ↑ dengue compared to baseline | Serum | Luminex |
| Dhenni et al., 2021 [68] | ID | Cross-sectional | Adults and children | DF: 43 | PDGF: FGF: | ↓ DF compared with chikungunya and HC ↓ DF compared to chikungunya and HC | Plasma | Luminex |
| Voraphani et al., 2010 [69] | TH | Cohort | Children | DF: 17 DHF: 10 | HGF: | ↑ DHF and DF compared with HC (a, b) | Serum | ELISA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiestas Solórzano, V.E.; de Lima, R.C.; de Azeredo, E.L. The Role of Growth Factors in the Pathogenesis of Dengue: A Scoping Review. Pathogens 2022, 11, 1179. https://doi.org/10.3390/pathogens11101179
Fiestas Solórzano VE, de Lima RC, de Azeredo EL. The Role of Growth Factors in the Pathogenesis of Dengue: A Scoping Review. Pathogens. 2022; 11(10):1179. https://doi.org/10.3390/pathogens11101179
Chicago/Turabian StyleFiestas Solórzano, Victor Edgar, Raquel Curtinhas de Lima, and Elzinandes Leal de Azeredo. 2022. "The Role of Growth Factors in the Pathogenesis of Dengue: A Scoping Review" Pathogens 11, no. 10: 1179. https://doi.org/10.3390/pathogens11101179