
Antimicrobial Susceptibility Profiles among Pseudomonas aeruginosa Isolated from Professional SCUBA Divers with Otitis Externa, Swimming Pools and the Ocean at a Diving Operation in South Africa

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of P. aeruginosa
2.2. Antimicrobial Sensitivity Profiling
2.3. Detection of ESBL and MBL Production
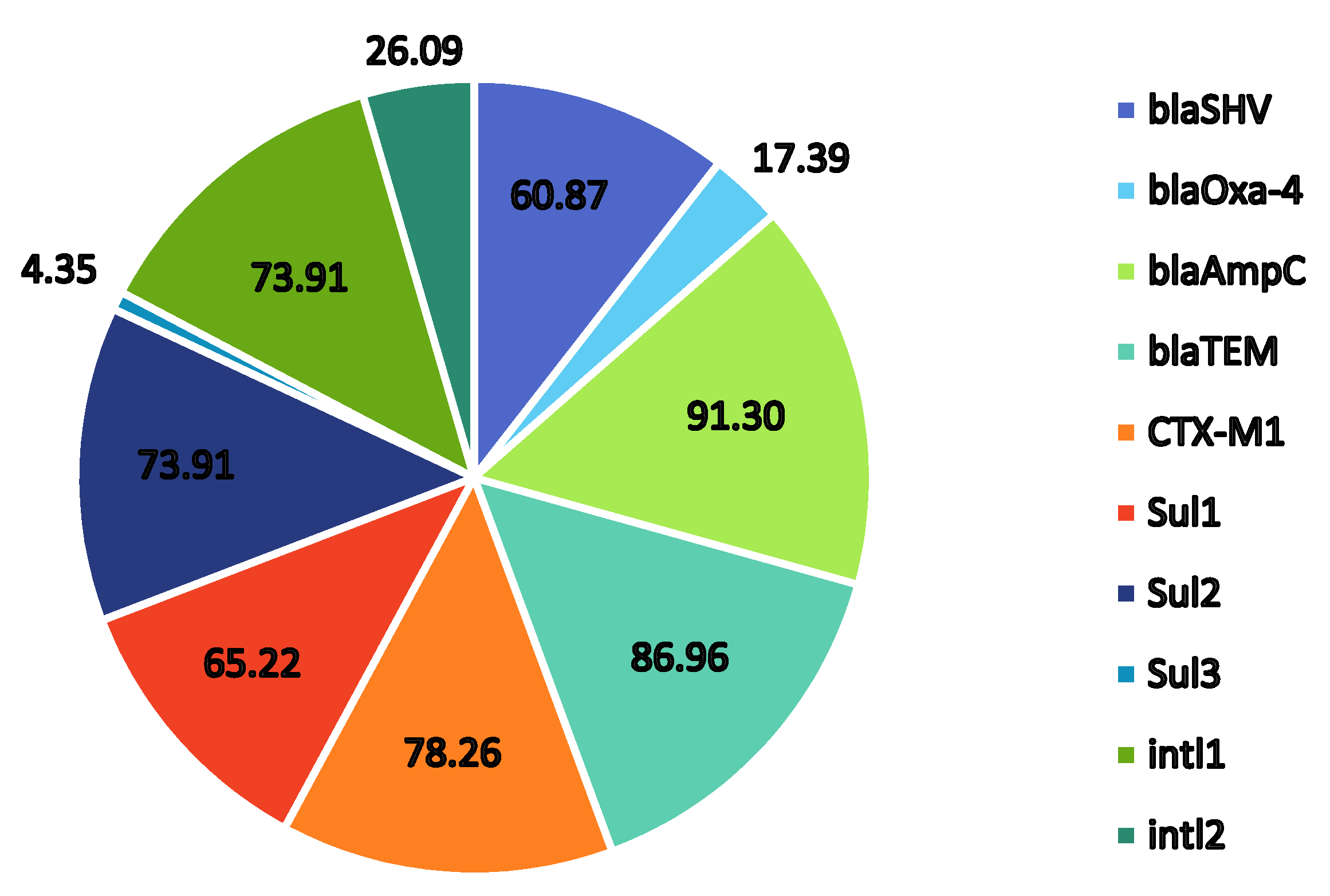
2.4. Presence of Trimethoprim–Sulfonamide-Resistant Genes
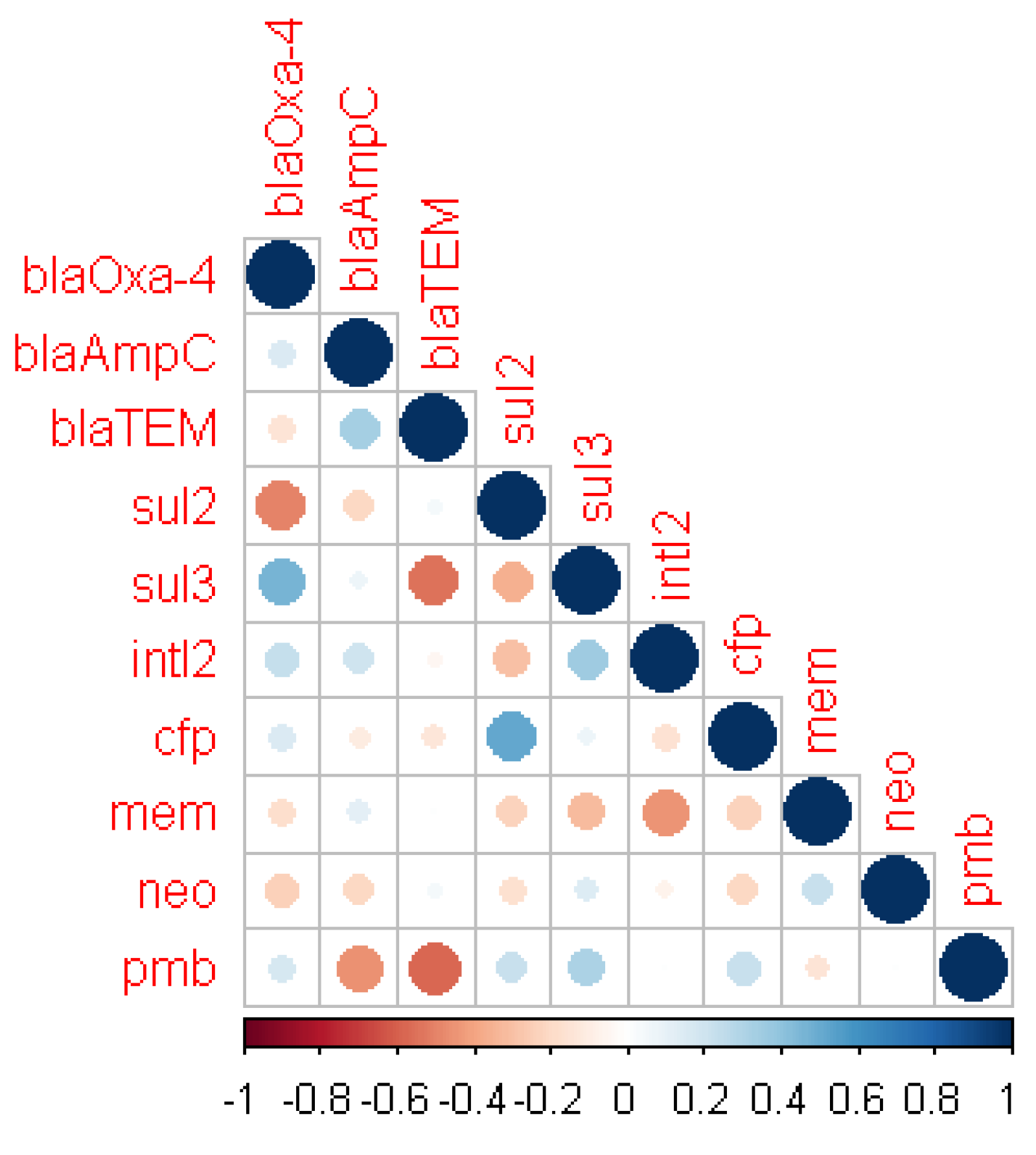
2.5. Correlation between Antibiotic Susceptibility and Resistance Genes
3. Materials and Methods
3.1. Ethics Statement
3.2. Sample Collection
3.3. Isolation and Identification of P. aeruginosa
3.4. Antimicrobial Sensitivity Testing
3.5. Phenotypic Detection of ESBLs and MBLs
3.6. PCR Detection of Genes Associated with Antimicrobial Resistance
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Wang, Q.; Low, W.; Wang, Y.; Zhu, X. Microbiological safety of household membrane water filter. J. Environ. Biol. 2013, 34, 481–487. [Google Scholar]
- Azizi, M.H. Ear disorders in Scuba Divers. Int. J. Occup. Environ. Med. 2011, 2, 21. [Google Scholar]
- Hlavsa, C.M.; Cikesh, L.B.; Roberts, A.V.; Kahler, M.A.; Vigar, M.; Hilborn, D.E.; Wade, J.T.; Roellig, D.M.; Murphy, L.J.; Xiao, L.; et al. Outbreaks associated with treated recreational water—United States, 2000–2014. Morb. Mortal. Wkly. Rep. 2018, 67, 547–551. [Google Scholar] [CrossRef]
- D’Agata, E. Pseudomonas aeruginosa and other pseudomonas species. In Infectious Disease Essentials, 8th ed.; Mandell, E.J., Dolin, R., Blaser, M.J., Eds.; Elsevier: Philadelphia, PA, USA, 2017; pp. 1–561. [Google Scholar]
- Geyer, M.; Howell-Jones, R.; Cunningham, R.; McNulty, C. Consensus of microbiology reporting of ear swab results to primary care clinicians in patients with otitis externa. Br. J. Biomed. Sci. 2011, 68, 174–180. [Google Scholar] [CrossRef]
- Rossolini, G.M.; Mantengoli, E. Treatment and control of severe infections caused by multi-resistant Pseudomonas aeruginosa. Clin. Microbiol. Infect. 2005, 11, 17–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillay, D.; Jardine, N.P. Recreational scuba divers knowledge regarding the audiological consequences of the sport. S. Afr. J. Sports Med. 2010, 22, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.P.; Singh, U. Malignant otitis externa: A review. J. Infect. Dis. Ther. 2015, 3, 204. [Google Scholar] [CrossRef]
- Illing, E.; Olaleye, O. Malignant otitis externa: A review of aetiology, presentation, investigations and current management strategies. Webmed Cent. Otorhinolaryngol. 2011, 2, WMC001725. [Google Scholar]
- Zowalaty, E.E.M.; Thani, A.A.A.; Webster, J.T.; Zowalaty, E.E.A.; Schweizer, P.H.; Nasrallah, N.G.; Marei, H.E.; Ashour, M.H. Pseudomonas aeruginosa: Arsenal of resistance mechanisms, decades of changing resistance profiles, and future antimicrobial therapies. Future Microbiol. 2015, 10, 1683–1706. [Google Scholar] [CrossRef]
- Puzniak, L.; De Pestel, D.D.; Srinivasan, A.; Ye, G.; Murray, J.; Merchant, S.; De Ryke, C.A.; Gupta, V. A combination antibiogram evaluation for Pseudomonas aeruginosa in respiratory and blood sources from intensive care unit (ICU) and non-ICU settings in U.S. hospitals. Antimicrob. Agents Chemother. 2019, 63, e02564-18. [Google Scholar] [CrossRef] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. WHO Pathogens Priority List Working Group. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Kali, A.; Srirangaraj, S.; Kumar, S.; Divya, H.A.; Kalyani, A.; Umadevi, S. Detection of metallo-β-lactamase producing Pseudomonas aeruginosa in intensive care units. Aust. Med. J. 2013, 6, 686–693. [Google Scholar] [CrossRef]
- Barbier, F.; Andremont, A.; Wolff, M.; Bouadma, L. Hospital-acquired pneumonia and ventilator-associated pneumonia: Recent advances in epidemiology and management. Curr. Opin. Pulm. Med. 2013, 19, 216–228. [Google Scholar] [CrossRef]
- Pachori, P.; Gothalwal, R.; Gandhi, P. Emergence of antibiotic resistance Pseudomonas aeruginosa in intensive care unit; a critical review. Genes Dis. 2019, 6, 109–119. [Google Scholar] [CrossRef]
- Shu, J.-C.; Kuo, A.-J.; Su, L.-H.; Liu, T.-P.; Lee, M.-H.; Su, I.-N.; Wu, T.-L. Development of carbapenem resistance in Pseudomonas aeruginosa is associated with OprD polymorphisms, particularly the amino acid substitution at codon 170. Antimicrob. Chemother. 2017, 72, 2489–2495. [Google Scholar] [CrossRef] [Green Version]
- Adjei, C.B.; Govinden, U.; Moodley, K.; Essack, S.Y. Molecular characterisation of multidrug-resistant Pseudomonas aeruginosa from a private hospital in Durban, South Africa. S. Afr. J. Infect. Dis. 2018, 33, 38–41. [Google Scholar]
- Ekwanzala, M.D.; Dewar, J.B.; Kamika, I.; Momba, M.N.B. Systematic review in South Africa reveals antibiotic resistance genes shared between clinical and environmental settings. Infect Drug Resist. 2018, 11, 1907–1920. [Google Scholar] [CrossRef] [Green Version]
- Kao, C.-Y.; Chen, S.-S.; Hung, K.-H.H.; Wu, H.-M.; Hsueh, P.-R.; Jing-Jou Yan, J.-J.; Wu, J.-J. Overproduction of active efflux pump and variations of OprD dominate in imipenem resistant Pseudomonas aeruginosa isolated from patients with bloodstream infections in Taiwan. BMC Microbiol. 2016, 16, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabar, M.M.; Mirkalantari, S.; Amoli, R.I. Detection of ctx-M gene in ESBL producing E. coli strains isolated from urinary tract infection in Semnan, Iran. Electron. Physician 2016, 8, 2686–26990. [Google Scholar] [CrossRef] [PubMed]
- Varriale, L.; Dipineto, L.; Russo, T.P.; Borrelli, L.; Romano, V.; D’Orazio, S.; Pace, A.; Menna, L.F.; Fioretti, A.; Santaniello, A. Antimicrobial resistance of Escherichia coli and Pseudomonas aeruginosa from companion birds. Antibiotics 2020, 9, 780. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Liu, H.; Lan, T.; Dong, L.; Hu, H.; Zhao, S.; Zhang, Y.; Zheng, N.; Wang, J. Antibiotic resistance patterns of Pseudomonas spp. isolated from raw milk revealed by whole genome sequencing. Front. Microbiol. 2020, 11, 1005. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Zerr, D.M.; McNutt, M.A.; Berry, J.E.; Burns, J.L.; Kapur, R.P. Pseudomonas aeruginosa syntrophy in chronically colonized airways of cystic fibrosis patients. Antimicrob. Agents Chemother. 2012, 56, 5971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakibaie, M.S.F.; Hashemi, A.; Adeli, S. Detection of TEM, SHV and PER type extended-spectrum beta-lactamase genes among clinical strains of Pseudomonas aeroginosa isolated from burnt patients Shafa hospital, Kerman, Iran. Iran J. Basic Med. Sci. 2008, 11, 104–111. [Google Scholar]
- Rossolini, G.M.; D’Andrea, M.M.; Mugnaioli, C. The spread of CTXM-type extended-spectrum beta-lactamases. Clin. Microbiol. Infect. 2008, 14 (Suppl. 1), 33–41. [Google Scholar] [CrossRef] [Green Version]
- Dabir, S.; Mohammadi, J.; Alizadeh, A.; Karimi, F.; Nori, K.; Mahmoudi, S.; Pournajafi, A. Investigation of the prevalence of CTX-M-1 beta-lactamase gene in Pseudomonas aeruginosa strains isolated from urinary tract infections in Zanjan Hospitals, Iran. S. Asian Res. J. Biol. Appl. Biosci. 2020, 2, 20–24. [Google Scholar] [CrossRef]
- Hsieh, W.-S.; Wang, N.-Y.; Feng, J.-A.; Weng, L.-C.; Wu, H.-H. Identification of DHA-23, a novel plasmid-mediated and inducible AmpC beta-lactamase from Enterobacteriaceae in Northern Taiwan. Front. Microbiol. 2015, 6, 436. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Ogo, M.; Koike, T.; Takada, H.; Newman, B. Sulfonamide and tetracycline resistance genes in total and culturable bacterial assemblages in South African aquatic environments. Front. Microbiol. 2015, 6, 796. [Google Scholar] [CrossRef]
- Xu, Z.; Li, L.; Shirtliff, M.E.; Alam, M.J.; Yamasaki, S.; Shi, L. Occurrence and characteristics of class 1 and 2 integrons in Pseudomonas aeruginosa isolates from patients in Southern China. J. Clin. Microbiol. 2009, 47, 230–234. [Google Scholar] [CrossRef] [Green Version]
- Quinn, P.J.; Markey, B.K.; Leonard, F.C.; Fitz Patrick, E.S.; Fanning, S.; Hartigan, P.J. Veterinary Microbiology and Microbial Diseases, 2nd ed.; Blackwell Publishing Ltd.: Ames, IA, USA, 2011; pp. 287–290. [Google Scholar]
- Clinical and Laboratory Standards Institute. Methods for Determining Bactericidal Activity of Antimicrobial Agents Approved Guideline; CLSI Document M100S; CLSI: Malvern, PA, USA, 2017; Volume 27, pp. 42–44. [Google Scholar]
- De, S.A.; Kumar, H.S.; Baveja, M.S. Prevalence of metallo-β-lactamase producing Pseudomonas aeruginosa and Acinetobacter species in intensive care areas in a tertiary care hospital. Indian J. Crit. Care Med. 2010, 14, 217–219. [Google Scholar] [CrossRef] [Green Version]
- Abrar, S.; Ain, A.U.; Liaqat, H.; Hussain, S.; Rasheed, F.; Riaz, S. Distribution of blaCTX—M, blaTEM, blaSHV and blaOXA genes in extended-spectrum-β-lactamase-producing clinical isolates: A three-year multi-center study from Lahore, Pakistan. Antimicrob. Resist. Infect. Control 2019, 8, 80. [Google Scholar] [CrossRef]
- Jouini, A.; Vinué, L.; Slama, K.B.; Sáenz, Y.; Klibi, N.; Hammami, S.; Boudabous, A.; Torres, C. Characterization of CTX-M and SHV extended-spectrum beta-lactamases and associated resistance genes in Escherichia coli strains of food samples in Tunisia. J. Antimicrob. Chemother. 2007, 60, 1137–1141. [Google Scholar] [CrossRef] [PubMed]
- Machado, E.; Cantón, R.; Baquero, F.; Galán, J.C.; Rollán, A.; Peixe, L.; Coque, T.M. Integron content of extended-spectrum-beta-lactamase-producing Escherichia coli strains over 12 years in a single hospital in Madrid, Spain. Antimicrob. Agents Chemother. 2005, 49, 1823–1829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Foundation for Statistical Computing; R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R.-project.org/ (accessed on 23 December 2021).
- Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://github.com/taiyun/corrplot (accessed on 23 December 2021).



{kind=link}
{kind=link}
| Antibiotics | Abbreviation | Total Number of Resistant Isolates n = 22 | |
|---|---|---|---|
| S No. (%) | R No. (%) | ||
| β-lactams | |||
| Cefotaxime | cfx | 0(0) | 22(100) |
| Cefaperazone | cfp | 2(9.1) | 20(90.9) |
| Ceftazidime | caz | 6(227.3) | 16(72.7) |
| Meropenem | mem | 10(45.5) | 12(54.5) |
| Amoxicillin | amx | 0(0) | 22(100) |
| Augmentin | amc | 0(0) | 22(100) |
| Aztreonam | atm | 4(18.2) | 18(81.8) |
| Imipenem | ipm | 0(0) | 22(100) |
| Piperacillin–Tazobactam | tzp | 0(0) | 22(100) |
| Penicillin G | pen | 0(0) | 22(100) |
| Non β-lactams | |||
| Neomycin | neo | 10(45.5) | 12(54.5) |
| Ciprofloxacin | cip | 0(0) | 22(100) |
| Amikacin | amk | 0(0) | 22(100) |
| Polymyxin B | pmb | 15(68.2) | 7(31.8) |
| Ofloxacin | ofx | 0(0) | 22(100) |
| Bactrim (Trimethoprim–sulfamethoxazole) | sxt | 22(100) | 0(0) |
| Resistant Gene 1 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Isolate 2 | blaSHV | blaOxa-4 | blaAmpC | blaTEM | CTX-M1 | Sul1 | Sul2 | Sul3 | intl1 | intl2 |
| BPT2 | + | - | - | - | + | - | + | - | - | - |
| APT1 | + | - | + | + | + | + | + | - | + | - |
| APT2 | - | - | + | + | + | + | + | - | - | + |
| ADO1 | + | + | + | + | + | + | - | - | + | - |
| ADO2 | - | - | + | + | + | + | - | - | + | + |
| OE1 | - | - | + | + | + | - | + | - | + | - |
| OE2 | - | + | + | + | + | - | - | - | + | - |
| OE3 | + | - | + | + | + | - | + | - | - | - |
| OE4 | - | - | + | + | + | + | + | - | + | - |
| OE5 | + | - | + | + | + | - | + | - | + | - |
| OE6 | + | - | - | + | + | + | + | - | + | - |
| OE8 | + | - | + | + | - | - | + | - | + | - |
| TwOE1 | + | - | + | + | + | + | + | - | + | - |
| PW1 | + | + | + | - | - | + | - | + | + | + |
| PW2 | - | - | + | + | + | + | + | - | + | - |
| PW3 | + | - | + | + | + | - | - | - | + | + |
| OSW1 | + | + | + | + | - | + | + | - | + | + |
| OSW2 3 | - | - | + | + | + | + | + | - | + | - |
| OSW3 | + | - | + | + | + | + | + | - | + | - |
| OBW1 | + | - | + | + | + | + | - | - | - | - |
| OBW2 | + | - | + | - | + | + | + | - | + | - |
| OBW3 | - | - | + | + | - | - | + | - | - | - |
| OBW4 | - | - | + | + | - | + | + | - | - | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maclean, K.; Njamo, F.O.J.P.; Serepa-Dlamini, M.H.; Kondiah, K.; Green, E. Antimicrobial Susceptibility Profiles among Pseudomonas aeruginosa Isolated from Professional SCUBA Divers with Otitis Externa, Swimming Pools and the Ocean at a Diving Operation in South Africa. Pathogens 2022, 11, 91. https://doi.org/10.3390/pathogens11010091
Maclean K, Njamo FOJP, Serepa-Dlamini MH, Kondiah K, Green E. Antimicrobial Susceptibility Profiles among Pseudomonas aeruginosa Isolated from Professional SCUBA Divers with Otitis Externa, Swimming Pools and the Ocean at a Diving Operation in South Africa. Pathogens. 2022; 11(1):91. https://doi.org/10.3390/pathogens11010091
Chicago/Turabian StyleMaclean, Kevin, Fernande Olpa J Pankendem Njamo, Mahloro Hope Serepa-Dlamini, Kulsum Kondiah, and Ezekiel Green. 2022. "Antimicrobial Susceptibility Profiles among Pseudomonas aeruginosa Isolated from Professional SCUBA Divers with Otitis Externa, Swimming Pools and the Ocean at a Diving Operation in South Africa" Pathogens 11, no. 1: 91. https://doi.org/10.3390/pathogens11010091