
Ecology of Ixodes pacificus Ticks and Associated Pathogens in the Western United States



Abstract
:1. Introduction
2. Habitat
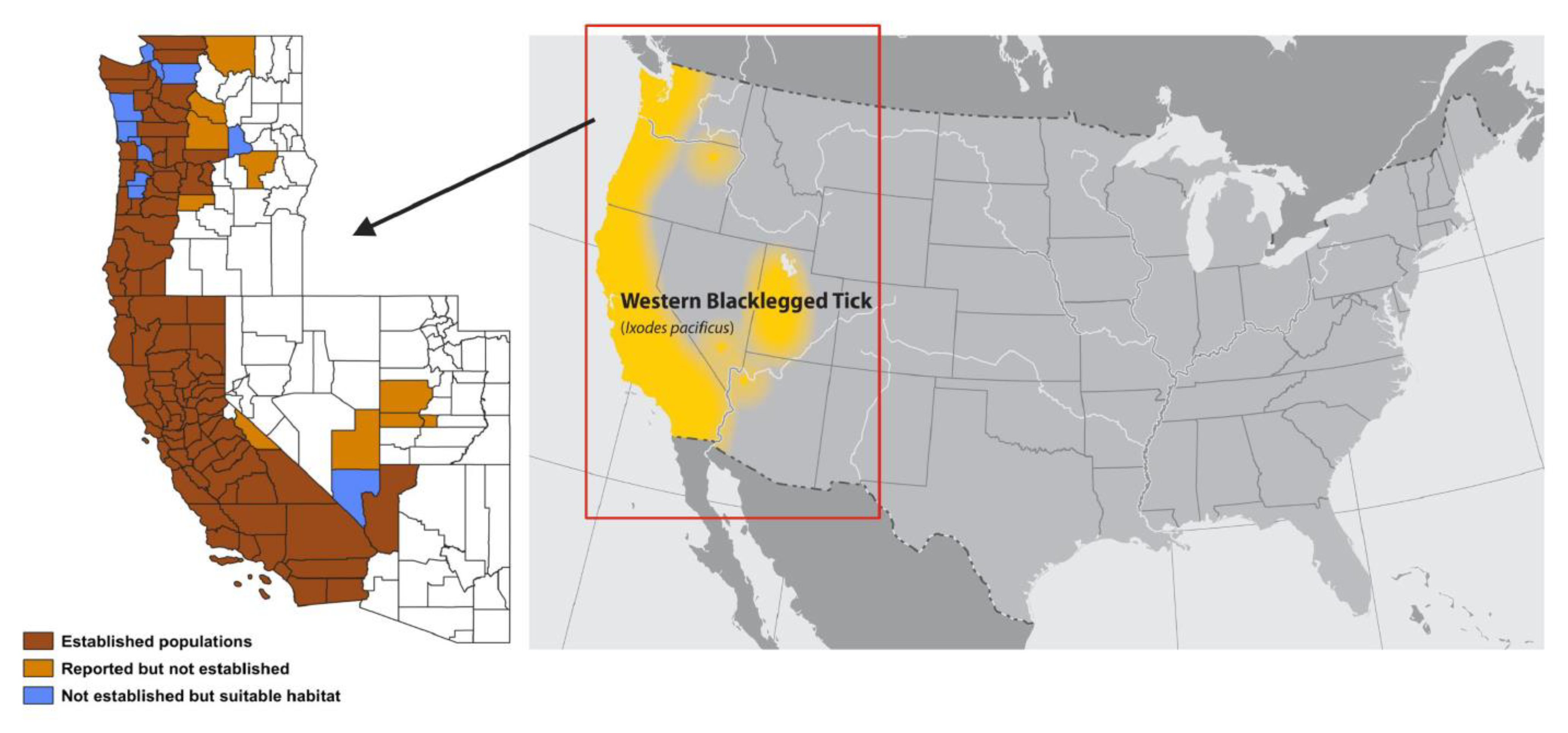
3. Tick Distribution
4. Hosts
5. Life Cycle
6. Environmental and Climate Factors Associated with Survival
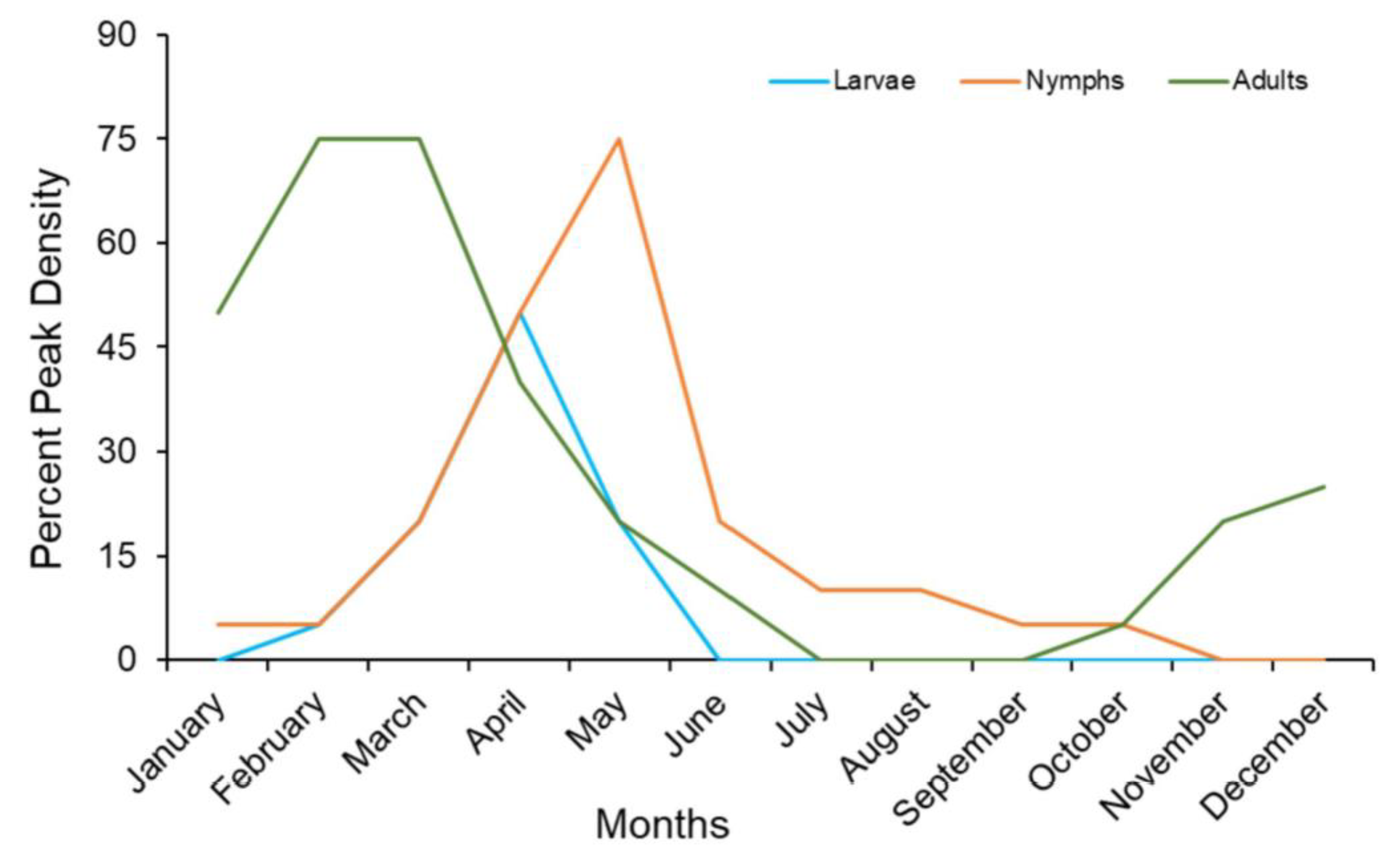
7. Seasonal Density
8. Infection Prevalence in Ixodes pacificus
8.1. Babesia Species
8.2. Bartonella Species
8.3. Borrelia Species
8.4. Rickettsiales Bacteria
8.4.1. Rickettsia Species
8.4.2. Anaplasma Species
8.4.3. Ehrlichia Species

{kind=link}
{kind=link}
| Disease | Pathogen | Reservoir Host | Reference |
|---|---|---|---|
| Babesiosis | Babesia odocoilei | Deer species | [61,62,63,64,65] |
| Bartonellosis | Bartonella hensele, B. quintana, B. washoensis, B. vinsonii subsp. berkhoffi | Ground squirrel (Otospermophilus beecheyi), Deer mouse (Peromyscus maniculatus), Brush mouse (P.boylii), Pinyon mouse (P. truei), Dusky-footed woodrat (Neotoma fuscipes) | [66,67,68,69,70,71,72,73,74] |
| Lyme Disease (B. burgdorferi), Hard Tick Relapsing Fever (B. miyamotoi) | Borrelia burgdorferi, B. miyamotoi, B. americana, B. bissettiae | Western grey squirrel (Sciurus griseus), birds of the order Passeriforms | [21,41,45,46,75,76,77,78,79,80,81,82] |
| Anaplasmosis | Anaplasma phagocytophilum | Dusky-footed woodrat (Neotoma fuscipes), Tree squirrels (Sciurus sp.), Chipmunk (Tamias sp.), Golden-crowned sparrow (Zonotrichia atricapilla), European starling (Sturnus vulgaris) | [89,90,91,92,93,94] |
| Human Monocytic Ehrlichosis, Human Granulocytic Ehrlichosis | Ehrlichia chaffensis, E. equi, E. phagocytophila | Deer species | [96,97,98,99] |
9. Pathogen Coinfection in I. pacificus
10. Tick Control
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caminade, C.; McIntyre, K.M.; Jones, A.E. Impact of recent and future climate change on vector-borne diseases. Ann. N. Y. Acad. Sci. 2019, 1436, 157–173. [Google Scholar] [CrossRef] [Green Version]
- Kilpatrick, A.M.; Randolph, S.E. Drivers, dynamics, and control of emerging vector-borne zoonotic diseases. Lancet 2012, 380, 1946–1955. [Google Scholar] [CrossRef] [Green Version]
- Ogden, N.H.; Artsob, H.; Lindsay, L.R.; Sockett, P.N. Lyme disease: A zoonotic disease of increasing importance to Canadians. Can. Fam. Physician 2008, 54, 1381–1384. [Google Scholar]
- Pascoe, E.L.; Plourde, B.T.; Lopéz-Perez, A.M.; Foley, J.E. Response of small mammal and tick communities to a catastrophic wildfire and implications for tickborne pathogens. J. Vector Ecol. 2020, 45, 269–284. [Google Scholar] [CrossRef]
- Allan, B.F. Influence of prescribed burns on the abundance of Amblyomma americanum (Acari: Ixodidae) in the Missouri Ozarks. J. Med. Entomol. 2009, 46, 1030–1036. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Ostfeld, R.S.; Peterson, A.T.; Poulin, R.; de la Fuente, J. Effects of environmental change on zoonotic disease risk: An ecological primer. Trends Parasitol. 2014, 30, 205–214. [Google Scholar] [CrossRef]
- Diuk-Wasser, M.A.; Hoen, A.G.; Cislo, P.; Brinkerhoff, R.; Hamer, S.A.; Rowland, M.; Cortinas, R.; Vourc’h, G.; Melton, F.; Hickling, G.J.; et al. Human risk of infection with Borrelia burgdorferi, the Lyme disease agent, in eastern United States. Am. J. Trop. Med. Hyg. 2012, 86, 320–327. [Google Scholar] [CrossRef]
- Piesman, J.; Eisen, L. Prevention of tickborne diseases. Annu. Rev. Entomol. 2008, 53, 323–343. [Google Scholar] [CrossRef] [PubMed]
- Burgdorfer, W.; Lane, R.S.; Barbour, A.G.; Gresbrink, R.A.; Anderson, J.R. The western black-legged tick, Ixodes pacificus: A vector of Borrelia burgdorferi. Am. J. Trop. Med. Hyg. 1985, 34, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.S.; Brown, R.N.; Piesman, J.; Peavey, C.A. Vector competence of Ixodes pacificus and Dermacentor occidentalis (Acari: Ixodidae) for various isolates of Lyme disease spirochetes. J. Med. Entomol. 1994, 31, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Eisen, R.J.; Eisen, L.; Beard, C.B. County-Scale Distribution of Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae) in the Continental United States. J. Med. Entomol. 2016, 53, 349–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clover, J.R.; Lane, R.S. Evidence implicating nymphal Ixodes pacificus (Acari: Ixodidae) in the epidemiology of Lyme disease in California. Am. J. Trop. Med. Hyg. 1995, 53, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Rollend, L.; Fish, D.; Childs, J.E. Transovarial transmission of Borrelia spirochetes by Ixodes scapularis: A summary of the literature and recent observations. Ticks Tick Borne Dis. 2013, 4, 46–51. [Google Scholar] [CrossRef]
- Kurokawa, C.; Lynn, G.E.; Pedra, J.; Pal, U.; Narasimhan, S.; Fikrig, E. Interactions between Borrelia burgdorferi and ticks. Nat. Rev. Microbiol. 2020, 18, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Kurtenbach, K.; Hanincová, K.; Tsao, J.I.; Margos, G.; Fish, D.; Ogden, N.H. Fundamental processes in the evolutionary ecology of Lyme borreliosis. Nat. Rev. Microbiol. 2006, 4, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Hauck, D.; Jordan, D.; Springer, A.; Schunack, B.; Pachnicke, S.; Fingerle, V.; Strube, C. Transovarial transmission of Borrelia spp., Rickettsia spp. and Anaplasma phagocytophilum in Ixodes ricinus under field conditions extrapolated from DNA detection in questing larvae. Parasit. Vectors 2020, 13, 176. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. How Ticks Spread Disease. Available online: https://www.cdc.gov/ticks/life_cycle_and_hosts.html (accessed on 21 November 2021).
- Girard, Y.A.; Travinsky, B.; Schotthoefer, A.; Fedorova, N.; Eisen, R.J.; Eisen, L.; Barbour, A.G.; Lane, R.S. Population structure of the Lyme borreliosis spirochete Borrelia burgdorferi in the western black-legged tick (Ixodes pacificus) in Northern California. Appl. Environ. Microbiol. 2009, 75, 7243–7252. [Google Scholar] [CrossRef] [Green Version]
- Merten, H.A.; Durden, L.A. A state-by-state survey of ticks recorded from humans in the United States. J. Vector Ecol. 2000, 25, 102–113. [Google Scholar]
- Brown, R.N.; Lane, R.S. Lyme disease in California: A novel enzootic transmission cycle of Borrelia burgdorferi. Science 1992, 256, 1439–1442. [Google Scholar] [CrossRef]
- Xu, G.; Pearson, P.; Dykstra, E.; Andrews, E.S.; Rich, S.M. Human-Biting Ixodes Ticks and Pathogen Prevalence from California, Oregon, and Washington. Vector Borne Zoonotic Dis. 2019, 19, 106–114. [Google Scholar] [CrossRef] [Green Version]
- Margos, G.; Hojgaard, A.; Lane, R.S.; Cornet, M.; Fingerle, V.; Rudenko, N.; Ogden, N.; Aanensen, D.M.; Fish, D.; Piesman, J. Multilocus sequence analysis of Borrelia bissettii strains from North America reveals a new Borrelia species, Borrelia kurtenbachii. Ticks Tick Borne Dis. 2010, 1, 151–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margos, G.; Piesman, J.; Lane, R.S.; Ogden, N.H.; Sing, A.; Straubinger, R.K.; Fingerle, V. Borrelia kurtenbachii sp. nov., a widely distributed member of the Borrelia burgdorferi sensu lato species complex in North America. Int. J. Syst. 2014, 64 Pt 1, 128–130. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.; Golovchenko, M.; Lin, T.; Gao, L.; Grubhoffer, L.; Oliver, J.H., Jr. Delineation of a new species of the Borrelia burgdorferi Sensu Lato Complex, Borrelia americana sp. nov. J. Clin. Microbiol. 2009, 47, 3875–3880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postic, D.; Ras, N.M.; Lane, R.S.; Hendson, M.; Baranton, G. Expanded diversity among Californian Borrelia isolates and description of Borrelia bissettii sp. nov. (formerly Borrelia group DN127). J. Clin. Microbiol. 1998, 36, 3497–3504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postic, D.; Garnier, M.; Baranton, G. Multilocus sequence analysis of atypical Borrelia burgdorferi sensu lato isolates—Description of Borrelia californiensis sp. nov., and genomospecies 1 and 2. Int. J. Med. Microbiol. 2007, 297, 263–271. [Google Scholar] [CrossRef]
- Margos, G.; Fedorova, N.; Kleinjan, J.E.; Hartberger, C.; Schwan, T.G.; Sing, A.; Fingerle, V. Borrelia lanei sp. nov. extends the diversity of Borrelia species in California. Int. J. Syst. 2017, 67, 3872–3876. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, A.M.; Hinckley, A.F.; Mead, P.S.; Hook, S.A.; Kugeler, K.J. Surveillance for Lyme Disease—United States, 2008–2015. Morb. Mortal. Wkly. Report. Surveill. Summ. 2017, 66, 1–12. [Google Scholar] [CrossRef]
- Spielman, A.; Wilson, M.L.; Levine, J.F.; Piesman, J. Ecology of Ixodes dammini-borne human babesiosis and Lyme disease. Ann. Rev. Entomol. 1985, 30, 439–460. [Google Scholar] [CrossRef]
- Salkeld, D.J.; Leonhard, S.; Girard, Y.A.; Hahn, N.; Mun, J.; Padgett, K.A.; Lane, R.S. Identifying the reservoir hosts of the Lyme disease spirochete Borrelia burgdorferi in California: The role of the western gray squirrel (Sciurus griseus). Am. J. Trop. Med. Hyg. 2008, 79, 535–540. [Google Scholar] [CrossRef]
- Salkeld, D.J.; Castro, M.B.; Bonilla, D.; Kjemtrup, A.; Kramer, V.L.; Lane, R.S.; Padgett, K.A. Seasonal activity patterns of the western black-legged tick, Ixodes pacificus, in relation to onset of human Lyme disease in northwestern California. Ticks Tick Borne Dis. 2014, 5, 790–796. [Google Scholar] [CrossRef]
- Salkeld, D.J.; Lane, R.S. Community ecology and disease risk: Lizards, squirrels, and the Lyme disease spirochete in California, USA. Ecology 2010, 91, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.B.; Jarnevich, C.S.; Monaghan, A.J.; Eisen, R.J. Modeling the Geographic Distribution of Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae) in the Contiguous United States. J. Med. Entomol. 2016, 53, 1176–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisen, L.; Eisen, R.J.; Lane, R.S. Geographical distribution patterns and habitat suitability models for presence of host-seeking ixodid ticks in dense woodlands of Mendocino County, California. J. Med. Entomol. 2006, 43, 415–427. [Google Scholar] [CrossRef]
- Lane, R.S.; Manweiler, S.A.; Stubbs, H.A.; Lennette, E.T.; Madigan, J.E.; Lavoie, P.E. Risk factors for Lyme disease in a small rural community in northern California. Am. J. Epidemiol. 1992, 136, 1358–1368. [Google Scholar] [CrossRef] [PubMed]
- Padgett, K.; Bonilla, D.; Kjemtrup, A.; Vilcins, I.M.; Yoshimizu, M.H.; Hui, L.; Sola, M.; Quintana, M.; Kramer, V. Large scale spatial risk and comparative prevalence of Borrelia miyamotoi and Borrelia burgdorferi sensu lato in Ixodes pacificus. PLoS ONE 2014, 9, e110853. [Google Scholar] [CrossRef]
- Billeter, S.A.; Yoshimizu, M.H.; Hu, R. Species composition and temporal distribution of adult ixodid ticks and prevalence of Borrelia burgdorferi sensu lato and Rickettsia species in Orange County, California. J. Vector Ecol. 2017, 42, 189–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, D.T.; Nekomoto, T.S.; Victor, J.C.; Paul, W.S.; Piesman, J. Reported distribution of Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae) in the United States. J. Med. Entomol. 1998, 35, 629–638. [Google Scholar] [CrossRef]
- MacDonald, A.J.; Briggs, C.J. Truncated seasonal activity patterns of the western blacklegged tick (Ixodes pacificus) in central and southern California. Ticks Tick Borne Dis. Dis. 2016, 7, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Slowik, T.J.; Lane, R.S. Nymphs of the western black-legged tick (Ixodes pacificus) collected from tree trunks in woodland-grass habitat. J. Vector Ecol. 2001, 26, 165–171. [Google Scholar]
- Macdonald, A.J.; Hyon, D.W.; Brewington, J.B.; O’Connor, K.E.; Swei, A.; Briggs, C.J. Lyme disease risk in southern California: Abiotic and environmental drivers of Ixodes pacificus (Acari: Ixodidae) density and infection prevalence with Borrelia burgdorferi. Parasit. Vectors 2017, 10, 7. [Google Scholar] [CrossRef] [Green Version]
- Padgett, K.A.; Bonilla, D.L. Novel exposure sites for nymphal Ixodes pacificus within picnic areas. Ticks Tick Borne Dis. 2011, 2, 191–195. [Google Scholar] [CrossRef]
- Davis, R.S.; Ramirez, R.A.; Anderson, J.L.; Bernhardt, S.A. Distribution and Habitat of Ixodes pacificus (Acari: Ixodidae) and Prevalence of Borrelia burgdorferi in Utah. J. Med. Entomol. 2015, 52, 1361–1367. [Google Scholar] [CrossRef]
- Porter, W.T.; Barrand, Z.A.; Wachara, J.; Davall, K.; Mihaljevic, J.R.; Pearson, T.; Salkeld, D.J.; Nieto, N.C. Predicting the current and future distribution of the western black-legged tick, Ixodes pacificus, across the Western US using citizen science collections. PLoS ONE 2021, 16, e0244754. [Google Scholar] [CrossRef] [PubMed]
- Castro, M.B.; Wright, S.A. Vertebrate hosts of Ixodes pacificus (Acari: Ixodidae) in California. J. Vector Ecol. 2007, 32, 140–149. [Google Scholar] [CrossRef]
- Wright, S.A.; Tucker, J.R.; Donohue, A.M.; Castro, M.B.; Kelley, K.L.; Novak, M.G.; Macedo, P.A. Avian Hosts of Ixodes pacificus (Acari: Ixodidae) and the Detection of Borrelia burgdorferi in Larvae Feeding on the Oregon Junco. J. Med. Entomol. 2011, 48, 852–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieto, N.C.; Holmes, E.A.; Foley, J.E. Survival rates of immature Ixodes pacificus (Acari: Ixodidae) ticks estimated using field-placed enclosures. J. Vector Ecol. 2010, 35, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Eisen, L.; Eisen, R.J.; Lane, R.S. The roles of birds, lizards, and rodents as hosts for the western black-legged tick Ixodes pacificus. J. Vector Ecol. 2004, 29, 295–308. [Google Scholar]
- Eisen, L.; Eisen, R.J.; Lane, R.S. Seasonal activity patterns of Ixodes pacificus nymphs in relation to climatic conditions. Med. Vet. Entomol. 2002, 16, 235–244. [Google Scholar] [CrossRef]
- Macdonald, A.J.; O’Neill, C.; Yoshimizu, M.H.; Padgett, K.A.; Larsen, A.E. Tracking seasonal activity of the western blacklegged tick across California. J. Appl. Ecol. 2019, 56, 2562–2573. [Google Scholar] [CrossRef]
- Peavey, C.A.; Lane, R.S. Field and laboratory studies on the timing of oviposition and hatching of the western black-legged tick, Ixodes pacificus (Acari: Ixodidae). Exp. Appl. Acarol. 1996, 20, 695–711. [Google Scholar] [CrossRef]
- Padgett, K.A.; Lane, R.S. Life cycle of Ixodes pacificus (Acari: Ixodidae): Timing of developmental processes under field and laboratory conditions. J. Med. Entomol. 2001, 38, 684–693. [Google Scholar] [CrossRef]
- Swei, A.; Meentemeyer, R.; Briggs, C.J. Influence of Abiotic and Environmental Factors on the Density and Infection Prevalence of Ixodes pacificus (Acari: Ixodidae) With Borrelia burgdorferi. J. Med. Entomol. 2011, 48, 20–28. [Google Scholar] [CrossRef]
- Eisen, R.J.; Eisen, L.; Ogden, N.H.; Beard, C.B. Linkages of Weather and Climate with Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae), Enzootic Transmission of Borrelia burgdorferi, and Lyme Disease in North America. J. Med. Entomol. 2016, 53, 250–261. [Google Scholar] [CrossRef] [Green Version]
- Eisen, R.J.; Feirer, S.; Padgett, K.A.; Hahn, M.B.; Monaghan, A.J.; Kramer, V.L.; Lane, R.S.; Kelly, M. Modeling Climate Suitability of the Western Blacklegged Tick in California. J. Med. Entomol. 2018, 55, 1133–1142. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, A.J.; Mccomb, S.; O’Neill, C.; Padgett, K.A.; Larsen, A.E. Projected climate and land use change alter western blacklegged tick phenology, seasonal host-seeking suitability and human encounter risk in California. Glob. Chang. Biol. 2020, 26, 5459–5474. [Google Scholar] [CrossRef]
- Macdonald, A.J.; Hyon, D.W.; Mcdaniels, A.; O’Connor, K.E.; Swei, A.; Briggs, C.J. Risk of vector tick exposure initially increases, then declines through time in response to wildfire in California. Ecosphere 2018, 9, e02227. [Google Scholar] [CrossRef]
- Padgett, K.A.; Casher, L.E.; Stephens, S.L.; Lane, R.S. Effect of Prescribed Fire for Tick Control in California Chaparral. J. Med. Entomol. 2009, 46, 1138–1145. [Google Scholar] [CrossRef]
- Macdonald, A.J. Abiotic and habitat drivers of tick vector abundance, diversity, phenology and human encounter risk in southern California. PLoS ONE 2018, 13, e0201665. [Google Scholar] [CrossRef] [Green Version]
- Eisen, R.J.; Clark, R.J.; Monaghan, A.J.; Eisen, L.; Delorey, M.J.; Beard, C.B. Host-Seeking Phenology of Ixodes pacificus (Acari: Ixodidae) Nymphs in Northwestern California in Relation to Calendar Week, Woodland Type, and Weather Conditions. J. Med. Entomol. 2017, 54, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Westblade, L.F.; Simon, M.S.; Mathison, B.A.; Kirkman, L.A. Babesia microti: From Mice to Ticks to an Increasing Number of Highly Susceptible Humans. J. Clin. Microbiol. 2017, 55, 2903–2912. [Google Scholar] [CrossRef] [Green Version]
- Eshoo, M.W.; Carolan, H.E.; Massire, C.; Chou, D.M.; Crowder, C.D.; Rounds, M.A.; Phillipson, C.A.; Schutzer, S.E.; Ecker, D.J. Survey of Ixodes pacificus Ticks in California Reveals a Diversity of Microorganisms and a Novel and Widespread Anaplasmataceae Species. PLoS ONE 2015, 10, e0135828. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.D.; Sajid, M.S.; Pascoe, E.L.; Foley, J.E. Detection of Babesia odocoilei in Humans with Babesiosis Symptoms. Diagnostics 2021, 11, 947. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.D.; Scott, C.M. Human Babesiosis Caused by Babesia duncani Has Widespread Distribution across Canada. Healthcare 2018, 6, 49. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Lu, X. Molecular Detection of Human Parasitic Pathogens, 1st ed.; Liu, D., Ed.; CRC Press: Boca Raton, FL, USA, 2012. [Google Scholar] [CrossRef]
- Chang, C.C.; Chomel, B.B.; Kasten, R.W.; Romano, V.; Tietze, N. Molecular Evidence of Bartonella spp. in Questing Adult Ixodes pacificus Ticks in California. J. Clin. Microbiol. 2001, 39, 1221–1226. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Hayashidani, H.; Pusterla, N.; Kasten, R.W.; Madigan, J.E.; Chomel, B.B. Investigation of Bartonella infection in ixodid ticks from California. Comp. Immunol. Microbiol. Infect. Dis. 2002, 25, 229–236. [Google Scholar] [CrossRef]
- Ziedins, A.C.; Chomel, B.B.; Kasten, R.W.; Kjemtrup, A.M.; Chang, C.-C. Molecular epidemiology of Bartonella species isolated from ground squirrels and other rodents in northern California. Epidemiol. Infect. 2016, 144, 1837–1844. [Google Scholar] [CrossRef]
- Kosoy, M.Y.; Regnery, R.L.; Tzianabos, T.; Marston, E.L.; Jones, D.C.; Green, D.; Maupin, G.O.; Olson, J.G.; Childs, J.E. Distribution, diversity, and host specificity of Bartonella in rodents from the Southeastern United States. Am. J. Trop. Med. Hyg. 1997, 57, 578–588. [Google Scholar] [CrossRef]
- Jardine, C.; Appleyard, G.; Kosoy, M.Y.; McColl, D.; Chirino-Trejo, M.; Wobeser, G.; Leighton, F.A. Rodent-associated Bartonella in Saskatchewan, Canada. Vector-Borne Zoonotic Dis. 2005, 5, 402–409. [Google Scholar] [CrossRef]
- Bai, Y.; Kosoy, M.Y.; Ray, C.; Brinkerhoff, R.J.; Collinge, S.K. Temporal and Spatial Patterns of Bartonella Infection in Black-tailed Prairie Dogs (Cynomys ludovicianus). Microb. Ecol. 2008, 56, 373–382. [Google Scholar] [CrossRef]
- Kosoy, M.; Murray, M.; Gilmore, R.D.; Bai, Y., Jr.; Gage, K.L. Bartonella strains from ground squirrels are identical to Bartonella washoensis isolated from a human patient. J. Clin. Microbiol. 2003, 41, 645–650. [Google Scholar] [CrossRef] [Green Version]
- Welch, D.F.; Carroll, K.C.; Hofmeister, E.K.; Persing, D.H.; Robison, D.A.; Steigerwalt, A.G.; Brenner, D.J. Isolation of a new subspecies, Bartonella vinsonii subsp. arupensis, from a cattle rancher: Identity with isolates found in conjunction with Borrelia burgdorferi and Babesia microti among naturally infected mice. J. Clin. Microbiol. 1999, 37, 2598–2601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Calisher, C.H.; Kosoy, M.Y.; Root, J.J.; Doty, J.B. Persistent Infection or Successive Reinfection of Deer Mice with Bartonella vinsonii subsp. arupensis. Appl. Environ. Microbiol. 2011, 77, 1728–1731. [Google Scholar] [CrossRef] [Green Version]
- Lane, R.S.; Mun, J.; Peribáñez, M.A.; Stubbs, H.A. Host-seeking behavior of Ixodes pacificus (Acari: Ixodidae) nymphs in relation to environmental parameters in dense-woodland and woodland-grass habitats. J. Vector Ecol. 2007, 32, 342. [Google Scholar] [CrossRef]
- Lane, R.S.; Mun, J.; Peribáñez, M.A.; Fedorova, N. Differences in prevalence of Borrelia burgdorferi and Anaplasma spp. infection among host-seeking Dermacentor occidentalis, Ixodes pacificus, and Ornithodoros coriaceus ticks in northwestern California. Ticks Tick Borne Dis. Dis. 2010, 1, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swei, A.; Ostfeld, R.S.; Lane, R.S.; Briggs, C.J. Impact of the experimental removal of lizards on Lyme disease risk. Proc. Biol. Sci. 2011, 278, 2970–2978. [Google Scholar] [CrossRef]
- Lane, R.S.; Mun, J.; Eisen, L.; Eisen, R.J. Refractoriness of the Western Fence Lizard (Sceloporus occidentalis) to the Lyme Disease Group Spirochete Borrelia Bissettii. J. Parasitol. 2006, 92, 691–696. [Google Scholar] [CrossRef]
- Salkeld, D.J.; Lagana, D.M.; Wachara, J.; Porter, W.T.; Nieto, N.C. Examining Prevalence and Diversity of Tickborne Pathogens in Questing Ixodes pacificus Ticks in California. Appl. Environ. Microbiol. 2021, 87, e00319-21. [Google Scholar] [CrossRef] [PubMed]
- Swei, A.; Bowie, V.C.; Bowie, R.C.K. Comparative genetic diversity of Lyme disease bacteria in Northern Californian ticks and their vertebrate hosts. Ticks Tick Borne Dis. 2015, 6, 414–423. [Google Scholar] [CrossRef]
- Mun, J.; Eisen, R.J.; Eisen, L.; Lane, R.S. Detection of a Borrelia miyamotoi sensu lato relapsing-fever group spirochete from Ixodes pacificus in California. J. Med. Entomol. 2006, 43, 120–123. [Google Scholar] [CrossRef]
- Sambado, S.; Salomon, J.; Crews, A.; Swei, A. Mixed transmission modes promote persistence of an emerging tickborne pathogen. Ecosphere 2020, 11, e03171. [Google Scholar] [CrossRef]
- Cheng, D.; Lane, R.S.; Moore, B.D.; Zhong, J. Host blood meal-dependent growth ensures transovarial transmission and transstadial passage of Rickettsia sp. phylotype G021 in the western black-legged tick (Ixodes pacificus). Ticks Tick Borne Dis. 2013, 4, 421–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, D.; Vigil, K.; Schanes, P.; Brown, R.N.; Zhong, J. Prevalence and burden of two rickettsial phylotypes (G021 and G022) in Ixodes pacificus from California by real-time quantitative PCR. Ticks Tick Borne Dis. 2013, 4, 280–287. [Google Scholar] [CrossRef] [Green Version]
- Kwan, J.Y.; Griggs, R.; Chicana, B.; Miller, C.; Swei, A. Vertical vs. horizontal transmission of the microbiome in a key disease vector, Ixodes pacificus. Mol. Ecol. 2017, 26, 6578–6589. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.J.; Torkelson, J.L.; Bodnar, J.; Mortazavi, B.; Laurent, T.; Deason, J.; Thephavongsa, K.; Zhong, J. The Rickettsia Endosymbiont of Ixodes pacificus Contains All the Genes of De Novo Folate Biosynthesis. PLoS ONE 2015, 10, e0144552. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, J.L.; Fitch, S.; Rosati, A.; Zhong, J. The folA gene from the Rickettsia endosymbiont of Ixodes pacificus encodes a functional dihydrofolate reductase enzyme. Ticks Tick Borne Dis. 2018, 9, 443–449. [Google Scholar] [CrossRef]
- Kurlovs, A.H.; Li, J.; Cheng, D.; Zhong, J. Ixodes pacificus Ticks Maintain Embryogenesis and Egg Hatching after Antibiotic Treatment of Rickettsia Endosymbiont. PLoS ONE 2014, 9, e104815. [Google Scholar] [CrossRef]
- Dumler, J.S.; Barbet, A.F.; Bekker, C.P.; Dasch, G.A.; Palmer, G.H.; Ray, S.C.; Rikihisa, Y.; Rurangirwa, F.R. Reorganization of genera in the families Rickettsiaceae and Anaplasmataceae in the order Rickettsiales: Unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equi and ‘HGE agent’ as subjective synonyms of Ehrlichia phagocytophila. Int. J. Syst. Evol. Microbiol. 2001, 51, 2145–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rejmanek, D.; Freycon, P.; Bradburd, G.; Dinstell, J.; Foley, J. Unique strains of Anaplasma phagocytophilum segregate among diverse questing and non-questing Ixodes tick species in the western United States. Ticks Tick Borne Dis. 2013, 4, 482–487. [Google Scholar] [CrossRef] [Green Version]
- Nieto, N.C.; Foley, J.E. Evaluation of squirrels (Rodentia: Sciuridae) as ecologically significant hosts for Anaplasma phagocytophilum in California. J. Med. Entomol. 2008, 45, 763–769. [Google Scholar] [CrossRef]
- Nieto, N.C.; Foley, J.E. Reservoir Competence of the Redwood Chipmunk (Tamias Ochrogenys) for Anaplasma Phagocytophilum. Vector Borne Zoonotic Dis. 2009, 9, 573–577. [Google Scholar] [CrossRef] [Green Version]
- Foley, J.E.; Nieto, N.C.; Adjemian, J.; Dabritz, H.; Brown, R.N. Anaplasma phagocytophilum Infection in Small Mammal Hosts of Ixodes Ticks, Western United States. Emerg. Infect. Dis. 2008, 14, 1147–1150. [Google Scholar] [CrossRef] [PubMed]
- Nieto, N.C.; Foley, J.E.; Bettaso, J.; Lane, R.S. Reptile infection with Anaplasma phagocytophilum, the causative agent of granulocytic anaplasmosis. J. Parasitol. 2009, 95, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Dingler, R.J.; Wright, S.A.; Donohue, A.M.; Macedo, P.A.; Foley, J.E. Surveillance for Ixodes pacificus and the tickborne pathogens Anaplasma phagocytophilum and Borrelia burgdorferi in birds from California’s Inner Coast Range. Ticks Tick Borne Dis. 2014, 5, 436–445. [Google Scholar] [CrossRef]
- Reubel, G.H.; Kimsey, R.B.; Barlough, J.E.; Madigan, J.E. Experimental Transmission of Ehrlichia equi to Horses through Naturally Infected Ticks (Ixodes pacificus) from Northern California. J. Clin. Microbiol. 1998, 36, 2131–2134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, V.L.; Gutierrez, A.G.; Hui, L.T.; Irwin, W.E.; Randolph, M.P.; Vugia, D.J. Detection of the agents of human ehrlichioses in ixodid ticks from California. Am. J. Trop. Med. Hyg. 1999, 60, 62–65. [Google Scholar] [CrossRef] [Green Version]
- Richter, P.J.; Kimsey, R.B.; Madigan, J.E.; Barlough, J.E.; Dumler, J.S.; Brooks, D.L. Ixodes pacificus (Acari: Ixodidae) as a Vector of Ehrlichia equi (Rickettsiales: Ehrlichieae). J. Med. Entomol. 1996, 33, 1–5. [Google Scholar] [CrossRef]
- Barlough, J.E.; Madigan, J.E.; Turoff, D.R.; Clover, J.R.; Shelly, S.M.; Dumler, J.S. An Ehrlichia strain from a llama (Lama glama) and Llama-associated ticks (Ixodes pacificus). J. Clin. Microbiol. 1997, 35, 1005–1007. [Google Scholar] [CrossRef] [Green Version]
- Nieto, N.C.; Leonhard, S.; Foley, J.E.; Lane, R.S. Coinfection of Western Gray Squirrel (Sciurus griseus) and other Sciurid Rodents with Borrelia burgdorferi sensu stricto and Anaplasma phagocytophilum in California. J. Wildl. Dis. 2010, 46, 291–296. [Google Scholar] [CrossRef] [Green Version]
- Larson, S.R.; Lee, X.; Paskewitz, S.M. Prevalence of Tickborne Pathogens in Two Species of Peromyscus Mice Common in Northern Wisconsin. J. Med. Entomol. 2018, 55, 1002–1010. [Google Scholar] [CrossRef]
- Holden, K.; Boothby, J.T.; Anand, S.; Massung, R.F. Detection of Borrelia burgdorferi, Ehrlichia chaffeensis, and Anaplasma phagocytophilum in Ticks (Acari: Ixodidae) from a Coastal Region of California. J. Med. Entomol. 2003, 40, 534–539. [Google Scholar] [CrossRef] [Green Version]
- Dykstra, E.A.; Oltean, H.N.; Kangiser, D.; Marsden-Haug, N.; Rich, S.M.; Xu, G.; Lee, M.K.; Morshed, M.G.; Graham, C.B.; Eisen, R.J. Ecology and Epidemiology of Tickborne Pathogens, Washington, USA, 2011–2016. Emerg. Infect. Dis. 2020, 26, 648–657. [Google Scholar] [CrossRef] [Green Version]
- Nwanade, C.F.; Wang, M.; Wang, T.; Yu, Z.; Liu, J. Botanical acaricides and repellents in tick control: Current status and future directions. Exp. Appl. Acarol. 2020, 81, 1–35. [Google Scholar] [CrossRef]
- Stafford, K.C., III; Williams, S.C.; Molaei, G. Integrated Pest Management in Controlling Ticks and Tick-Associated Diseases. J. Integr. Pest. Manag. 2017, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Sajid, A.; Matias, J.; Arora, G.; Kurokawa, C.; DePonte, K.; Tang, X.; Lynn, G.; Wu, M.-J.; Pal, U.; Strank, N.O.; et al. mRNA vaccination induces tick resistance and prevents transmission of the lyme disease agent. Sci. Transl. Med. 2021, 13, eabj9827. [Google Scholar] [CrossRef] [PubMed]
- Richer, L.M.; Brisson, D.; Melo, R.; Ostfeld, R.S.; Zeidner, N.; Gomes-Solecki, M. Reservoir Targeted Vaccine Against Borrelia burgdorferi: A New Strategy to Prevent Lyme Disease Transmission. J. Infect. Dis. 2014, 209, 1972–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kianersi, S.; Luetke, M.; Wolfe, C.G.; Clark, W.A.; Omodior, O. Associations Between Personal Protective Measures and Self-Reported Tick-Borne Disease Diagnosis in Indiana Residents. J. Community Health. 2020, 45, 739–750. [Google Scholar] [CrossRef]
- Smith, G.; Wileyto, E.P.; Hopkins, R.B.; Cherry, B.R.; Maher, J.P. Risk factors for lyme disease in Chester County, Pennsylvania. Public Health Rep. 2001, 116 (Suppl. 1), 146–156. [Google Scholar] [CrossRef] [Green Version]
- Connally, N.P.; Durante, A.J.; Yousey-Hindes, K.M.; Meek, J.I.; Nelson, R.S.; Heimer, R. Peridomestic Lyme Disease Prevention. Am. J. Prev. Med. 2009, 37, 201–206. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McVicar, M.; Rivera, I.; Reyes, J.B.; Gulia-Nuss, M. Ecology of Ixodes pacificus Ticks and Associated Pathogens in the Western United States. Pathogens 2022, 11, 89. https://doi.org/10.3390/pathogens11010089
McVicar M, Rivera I, Reyes JB, Gulia-Nuss M. Ecology of Ixodes pacificus Ticks and Associated Pathogens in the Western United States. Pathogens. 2022; 11(1):89. https://doi.org/10.3390/pathogens11010089
Chicago/Turabian StyleMcVicar, Molly, Isabella Rivera, Jeremiah B. Reyes, and Monika Gulia-Nuss. 2022. "Ecology of Ixodes pacificus Ticks and Associated Pathogens in the Western United States" Pathogens 11, no. 1: 89. https://doi.org/10.3390/pathogens11010089