
West Nile Virus in Common Wild Avian Species in Israel

 ,
, 
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez-Ramírez, E.; Llorente, F.; Jiménez-Clavero, M.Á. Experimental infections of wild birds with West Nile virus. Viruses 2014, 6, 752–781. [Google Scholar] [CrossRef] [Green Version]
- Smithburn, K.C.; Hughes, T.P.; Burke, A.W.; Paul, J.H.; African, A. A neurotropic virus isolated from the blood of a native of uganda1. Am. J. Trop. Med. Hyg. 1940, 20, 471–492. [Google Scholar] [CrossRef]
- Murgue, B.; Murri, S.; Triki, H.; Deubel, V.; Zeller, H.G. West Nile in the Mediterranean Basin: 1950–2000. Ann. N. Y. Acad. Sci. 2001, 951, 117–126. [Google Scholar] [CrossRef]
- Sambri, V.; Capobianchi, M.R.; Cavrini, F.; Charrel, R.; Donoso-Mantke, O.; Escadafal, C.; Franco, L.; Gaibani, P.; Gould, E.A.; Niedrig, M.; et al. Diagnosis of west nile virus human infections: Overview and proposal of diagnostic protocols considering the results of external quality assessment studies. Viruses 2013, 5, 2329–2348. [Google Scholar] [CrossRef] [Green Version]
- Marra, P.P.; Griffing, S.; Caffrey, C.; Kilpatrick, A.M.; McLean, R.; Brand, C.; Saito, E.; Dupuis, A.P.; Kramer, L.; Novak, R. West Nile virus and wildlife. Bioscience 2004, 54, 393–402. [Google Scholar] [CrossRef] [Green Version]
- CDC West Nile Virus Activity—United States, 2007; CDC: Atlanta, GA, USA, 2008.
- Ain-Najwa, M.Y.; Yasmin, A.R.; Omar, A.R.; Arshad, S.S.; Abu, J.; Mohammed, H.O.; Kumar, K.; Loong, S.K.; Rovie-Ryan, J.J.; Mohd-Kharip-Shah, A.K. Evidence of West Nile virus infection in migratory and resident wild birds in west coast of peninsular Malaysia. One Heal. 2020, 10, 100134. [Google Scholar] [CrossRef] [PubMed]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The Global Ecology and Epidemiology of West Nile Virus. Biomed. Res. Int. 2015, 2015, 376230. [Google Scholar] [CrossRef] [Green Version]
- Sejvar, J.J. West Nile virus: An historical overview. Ochsner J. 2003, 5, 6–10. [Google Scholar] [PubMed]
- Van Der Meulen, K.M.; Pensaert, M.B.; Nauwynck, H.J. West Nile virus in the vertebrate world. Arch. Virol. 2005, 150, 637–657. [Google Scholar] [CrossRef]
- Tolsá, M.J.; García-Peña, G.E.; Rico-Chávez, O.; Roche, B.; Suzán, G. Macroecology of birds potentially susceptible to West Nile virus. Proc. R. Soc. B Biol. Sci. 2018, 285. [Google Scholar] [CrossRef] [Green Version]
- Gamino, V.; Höfle, U. Pathology and tissue tropism of natural West Nile virus infection in birds: A review. Vet. Res. 2013, 44, 39. [Google Scholar] [CrossRef] [Green Version]
- Donadieu, E.; Bahuon, C.; Lowenski, S.; Zientara, S.; Coulpier, M.; Lecollinet, S. Differential virulence and pathogenesis of West Nile viruses. Viruses 2013, 5, 2856–2880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, L.R.; Brault, A.C.; Nasci, R.S. West Nile virus: Review of the literature. JAMA—J. Am. Med. Assoc. 2013, 310, 308–315. [Google Scholar] [CrossRef]
- Sejvar, J.J. Clinical manifestations and outcomes of West Nile virus infection. Viruses 2014, 6, 606–623. [Google Scholar] [CrossRef] [Green Version]
- Steinman, A.; Banet, C.; Sutton, G.; Yadin, H.; Hadar, S.; Brill, A. Clinical signs of West Nile virus encephalomyelitis in horses during the outbreak in Israel in 2000. Vet. Rec. 2002, 151, 47–49. [Google Scholar] [CrossRef]
- Brault, A.C.; Langevin, S.A.; Ramey, W.N.; Fang, Y.; Beasley, D.W.C.; Barker, C.M.; Sanders, T.A.; Reisen, W.K.; Barrett, A.D.T.; Bowen, R.A. Reduced avian virulence and viremia of West Nile virus isolates from Mexico and Texas. Am. J. Trop. Med. Hyg. 2011, 85, 758–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brault, A.C.; Langevin, S.A.; Bowen, R.A.; Panella, N.A.; Biggerstaff, B.J.; Miller, B.R.; Komar, N. Differential virulence of West Nile strains for American Crows. Emerg. Infect. Dis. 2004, 10, 2161–2168. [Google Scholar] [CrossRef]
- Bernkopf, H.; Levine, S.; Nerson, R. Isolation of West Nile virus in Israel. J. Infect. Dis. 1952, 93, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Noach Baneth, C.; Malkinson, M.; Brill, A.; Samina, I.; Yadin, H.; Weissman, Y.; Pokamunski, S.; King, R.; Deubel, V. Phylogenetic Relationships of West Nile Viruses Isolated from Birds and Horses in Israel from 1997 to 2001. Virus Genes 2003, 26, 135–141. [Google Scholar] [CrossRef]
- Banet-noach, C.; Simanov, L.; Malkinson, M. in geese Direct (non-vector) transmission of West Nile virus in geese. Avian Pathol. 2010, 32, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Chowers, M.Y.; Lang, R.; Nassar, F.; Ben-David, D.; Giladi, M.; Rubinshtein, E.; Itzhaki, A.; Mishal, J.; Siegman-Igra, Y.; Kitzes, R.; et al. Clinical characteristics of the West Nile fever outbreak, Israel, 2000. Emerg. Infect. Dis. 2001, 7, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Schvartz, G.; Farnoushi, Y.; Berkowitz, A.; Edery, N.; Hahn, S.; Steinman, A.; Lublin, A.; Erster, O. Molecular characterization of the re-emerging West Nile virus in avian species and equids in Israel, 2018, and pathological description of the disease. Parasites Vectors 2020, 13, 528. [Google Scholar] [CrossRef]
- Lustig, Y.; Gosinov, R.; Zuckerman, N.; Glazer, Y.; Orshan, L.; Sofer, D.; Schwartz, E.; Schvartz, G.; Farnoushi, Y.; Lublin, A.; et al. Epidemiologic and phylogenetic analysis of the 2018 west Nile virus (WNV) outbreak in Israel demonstrates human infection of WNV lineage I. Eurosurveillance 2019, 24. [Google Scholar] [CrossRef]
- Lustig, Y.; Hindiyeh, M.; Orshan, L.; Weiss, L.; Koren, R.; Katz-Likvornik, S.; Zadka, H.; Glatman-Freedman, A.; Mendelson, E.; Shulman, L.M. Mosquito surveillance for 15 years reveals high genetic diversity among West Nile Viruses in Israel. J. Infect. Dis. 2016, 213, 1107–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lustig, Y.; Kaufman, Z.; Mannasse, B.; Koren, R.; Katz-Likvornik, S.; Orshan, L.; Glatman-Freedman, A.; Mendelson, E. West Nile virus outbreak in Israel in 2015: Phylogenetic and geographic characterization in humans and mosquitoes. Clin. Microbiol. Infect. 2017, 23, 986–993. [Google Scholar] [CrossRef] [Green Version]
- Walsh, A.S.; Glass, G.E.; Lesser, C.R.; Curriero, F.C. Predicting seasonal abundance of mosquitoes based on off-season meteorological conditions. Environ. Ecol. Stat. 2008, 15, 279–291. [Google Scholar] [CrossRef]
- Grubaugh, N.D.; Ebel, G.D. Dynamics of West Nile virus evolution in mosquito vectors. Curr. Opin. Virol. 2016, 21, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Rios, L.M.V.; Sheu, J.J.; Day, J.F.; Maruniak, J.E.; Sein’O, K.; Zaretsky, H.; Long, M.T. Environmental risk factors associated with west nile virus clinical disease in florida horses. Med. Vet. Entomol. 2009, 23, 357–366. [Google Scholar] [CrossRef]
- Reisen, W.K.; Fang, Y.; Martinez, V.M. Effects of temperature on the transmission of West Nile virus by Culex tarsalis (Diptera: Culicidae). J. Med. Entomol. 2006, 43, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Langevin, S.A.; Brault, A.C.; Panella, N.A.; Bowen, R.A.; Komar, N. Variation in virulence of West Nile virus strains for house sparrows (Passer Domesticus). Am. J. Trop. Med. Hyg. 2005, 72, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Komar, N.; Burns, J.; Carrie, D.; Panella, N.A.; Dusza, S.; Bryan, C. Serologic Evidence for West Nile Virus Infection in Birds in Staten Island, New York, After an Outbreak in 2000. Vector Borne Zoonoyic Dis. 2001, 1, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Komar, N.; Panella, N.A.; Burns, J.E.; Dusza, S.W.; Mascarenhas, T.M.; Talbot, T.O. Serologic Evidence for West Nile Virus Infection in Birds in the New York City Vicinity During an Outbreak in 1999. Emerg. Infect. Dis. 2001, 7, 621–623. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Oh, H.; Song, J.; Hur, M.; Suh, J.H.; Jheong, W.H.; Kim, J.T.; Oh, H.S.; Park, J.H. First detection of West Nile virus in domestic pigeon in Korea. J. Vet. Sci. 2016, 17, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Allison, A.B.; Mead, D.G.; Gibbs, S.E.J.; Hoffman, D.M.; Stallknecht, D.E. West Nile virus viremia in wild rock pigeons. Emerg. Infect. Dis. 2004, 10, 2252–2255. [Google Scholar] [CrossRef] [PubMed]
- Haussig, J.M.; Young, J.J.; Gossner, C.M.; Mezei, E.; Bella, A.; Sirbu, A.; Pervanidou, D.; Drakulovic, M.B.; Sudre, B. Early start of the west nile fever transmission season 2018 in Europe. Eurosurveillance 2018, 23, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Hartley, D.M.; Barker, C.M.; Le Menach, A.; Niu, T.; Gaff, H.D.; Reisen, W.K. Effects of temperature on emergence and seasonality of West Nile virus in California. Am. J. Trop. Med. Hyg. 2012, 86, 884–894. [Google Scholar] [CrossRef] [Green Version]
- Eiden, M.; Vina-Rodriguez, A.; Hoffmann, B.; Ziegler, U.; Groschup, M.H. Two new real-time quantitative reverse transcription polymerase chain reaction assays with unique target sites for the specific and sensitive detection of lineages 1 and 2 West Nile virus strains. J. Vet. Diagnostic Investig. 2010, 22, 748–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savage, H.M.; Ceianu, C.; Nicolescu, G.; Karabatsos, N.; Lanciotti, R.; Vladimirescu, A.; Laiv, L.; Ungureanu, A.; Romanca, C.; Tsai, T.F. Entomologic and avian investigations of an epidemic of West Nile fever in Romania in 1996, with serologic and molecular characterization of a virus isolate from mosquitoes. Am. J. Trop. Med. Hyg. 1999, 61, 600–611. [Google Scholar] [CrossRef] [Green Version]


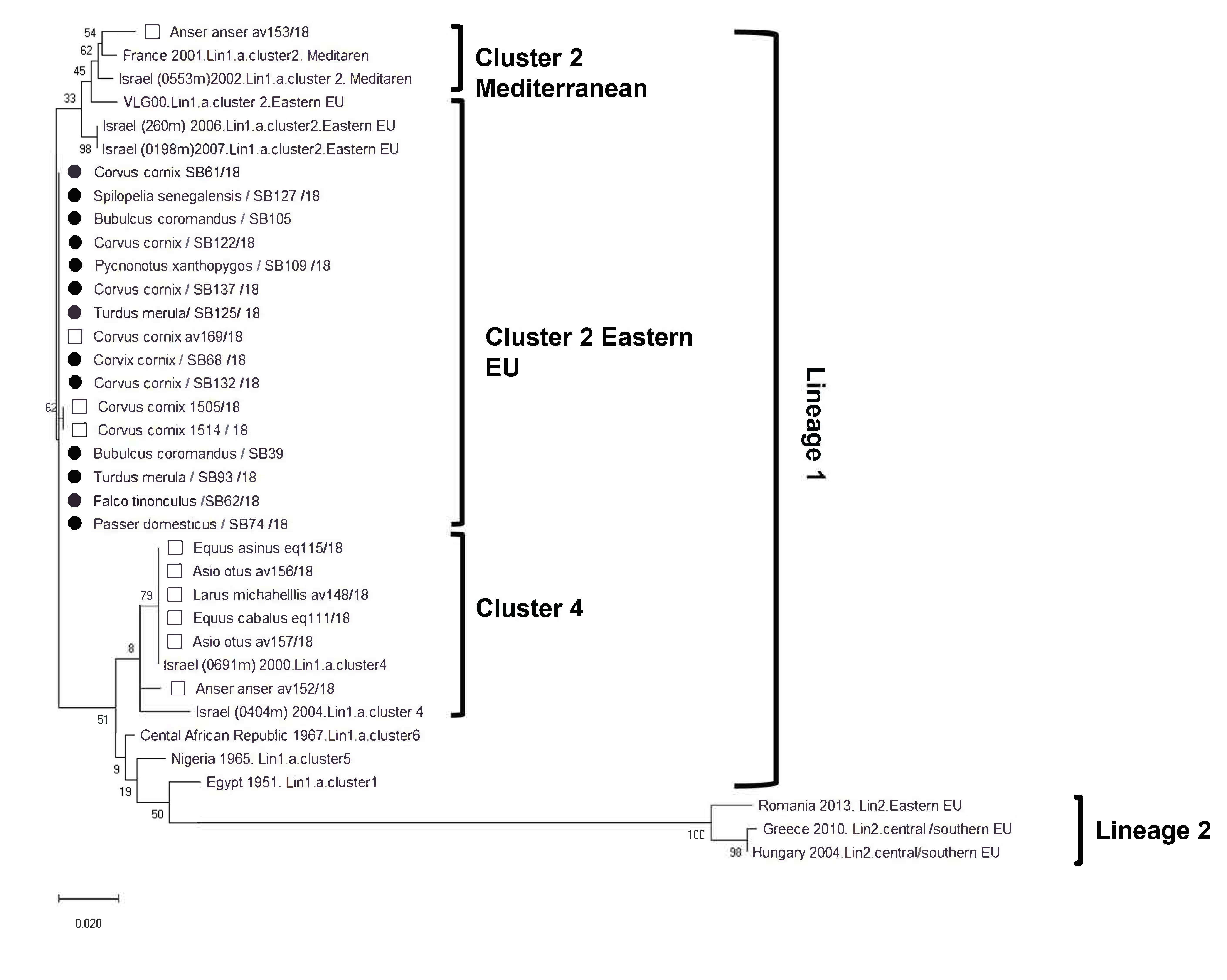{kind=link}
| Type (L/M/B) | Avian Species | n | WNV Positive |
|---|---|---|---|
| L | Acridotheres tristis | 3 | 0 |
| L | Bubulcus coromandus | 12 | 2 (16.7%) |
| L | Carduelis carduelis | 1 | 0 |
| L | Cinnyris | 1 | 0 |
| L | Columba domestica | 8 | 0 |
| L | Corvus cornix | 12 | 6 (50%) |
| L | Dendrocopos syriacus | 3 | 0 |
| L | Garrulus glandarius | 7 | 1 (14.3%) |
| L | Halcyon smyrnensis | 2 | 0 |
| L | Myiopsitta monachus | 2 | 0 |
| L | Parus major | 2 | 0 |
| L | Passer domesticus | 11 | 1 (9.1%) |
| L | Prinia gracilis | 2 | 0 |
| L | Psittacula krameri | 4 | 0 |
| L | Pycnonotus xanthopygos | 8 | 1 (12.5%) |
| L | Spilopelia senegalensis | 4 | 1 (25%) |
| B | Anas platyrhynchos | 1 | 0 |
| B | Corvus monedula | 1 | 0 |
| B | Falco tinnunculus | 19 | 1 (5.3%) |
| B | Hirundo rustica | 1 | 0 |
| B | Plegadis falcinellus | 1 | 0 |
| B | Turdus merula | 7 | 2 (28.6%) |
| B | Upopa epops | 7 | 0 |
| B | Venallus spinosus | 1 | 0 |
| M | Apus apus | 7 | 0 |
| M | Asio otus | 1 | 0 |
| M | Buteo buteo | 1 | 0 |
| M | Clamator glandarius | 1 | 0 |
| M | Coturnix coturnix | 1 | 0 |
| M | Otus scops | 3 | 0 |
| M | Streptopelia turtur | 1 | 0 |
| M | Sylvia melanocephala | 1 | 0 |
| Total | L: 60.6%, B: 27.7%, M: 11.7% | 136 | 15 (11.03%) |
| GenBank Accession No. | KVI/Avian Diseases No. | Sequence: |
|---|---|---|
| 2540881 | 327316 | Corvus cornix av169/18 |
| OL365014 | 1105/2 | Anser anser av153/18 |
| MK343718.1 | 1038 | Larus michahellis 148/18 |
| OL365013 | 1105/1 | Anser anser av152/18 |
| MK343719.1 | 1505 | Corvus cornix 1505/18 |
| OL450362 | 1514 | Corvus cornix 1514/18 |
| OL450363 | 1131 | Asio otus 157/18 |
| MT828576.1 | 324085 | Equus cabalus 111/18 |
| 2540897 | 1111 | Asio otus 156/18 |
| MT828577.1 | 325209 | Equus asinus 115/18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schvartz, G.; Tirosh-Levy, S.; Bider, S.; Lublin, A.; Farnoushi, Y.; Erster, O.; Steinman, A. West Nile Virus in Common Wild Avian Species in Israel. Pathogens 2022, 11, 107. https://doi.org/10.3390/pathogens11010107
Schvartz G, Tirosh-Levy S, Bider S, Lublin A, Farnoushi Y, Erster O, Steinman A. West Nile Virus in Common Wild Avian Species in Israel. Pathogens. 2022; 11(1):107. https://doi.org/10.3390/pathogens11010107
Chicago/Turabian StyleSchvartz, Gili, Sharon Tirosh-Levy, Shahar Bider, Avishai Lublin, Yigal Farnoushi, Oran Erster, and Amir Steinman. 2022. "West Nile Virus in Common Wild Avian Species in Israel" Pathogens 11, no. 1: 107. https://doi.org/10.3390/pathogens11010107