
Non-Penicillin-Susceptible Streptococcus suis Isolated from Humans

 , and
, and 
Abstract
:1. Introduction
2. Results
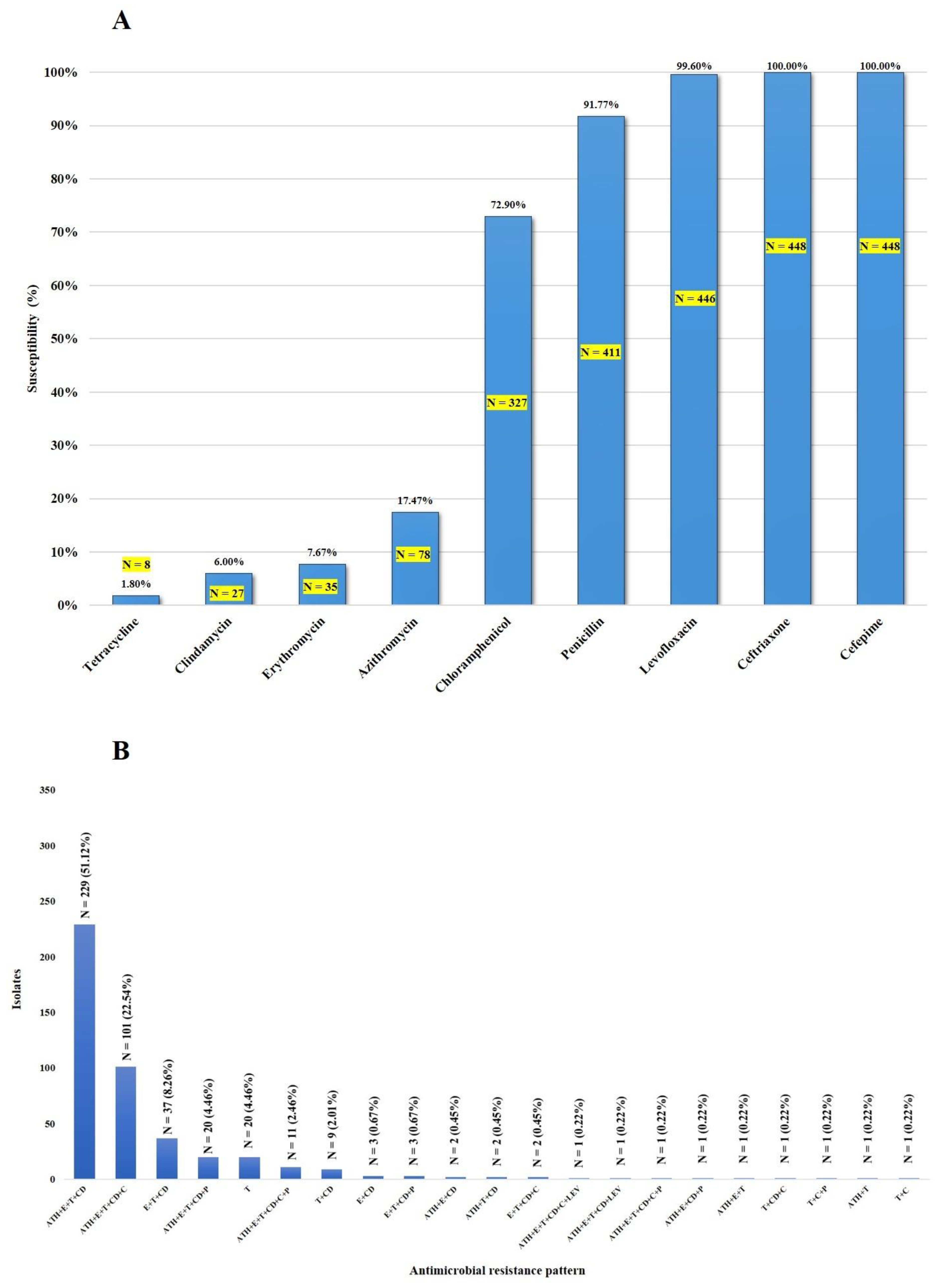
2.1. Antimicrobial Susceptibility
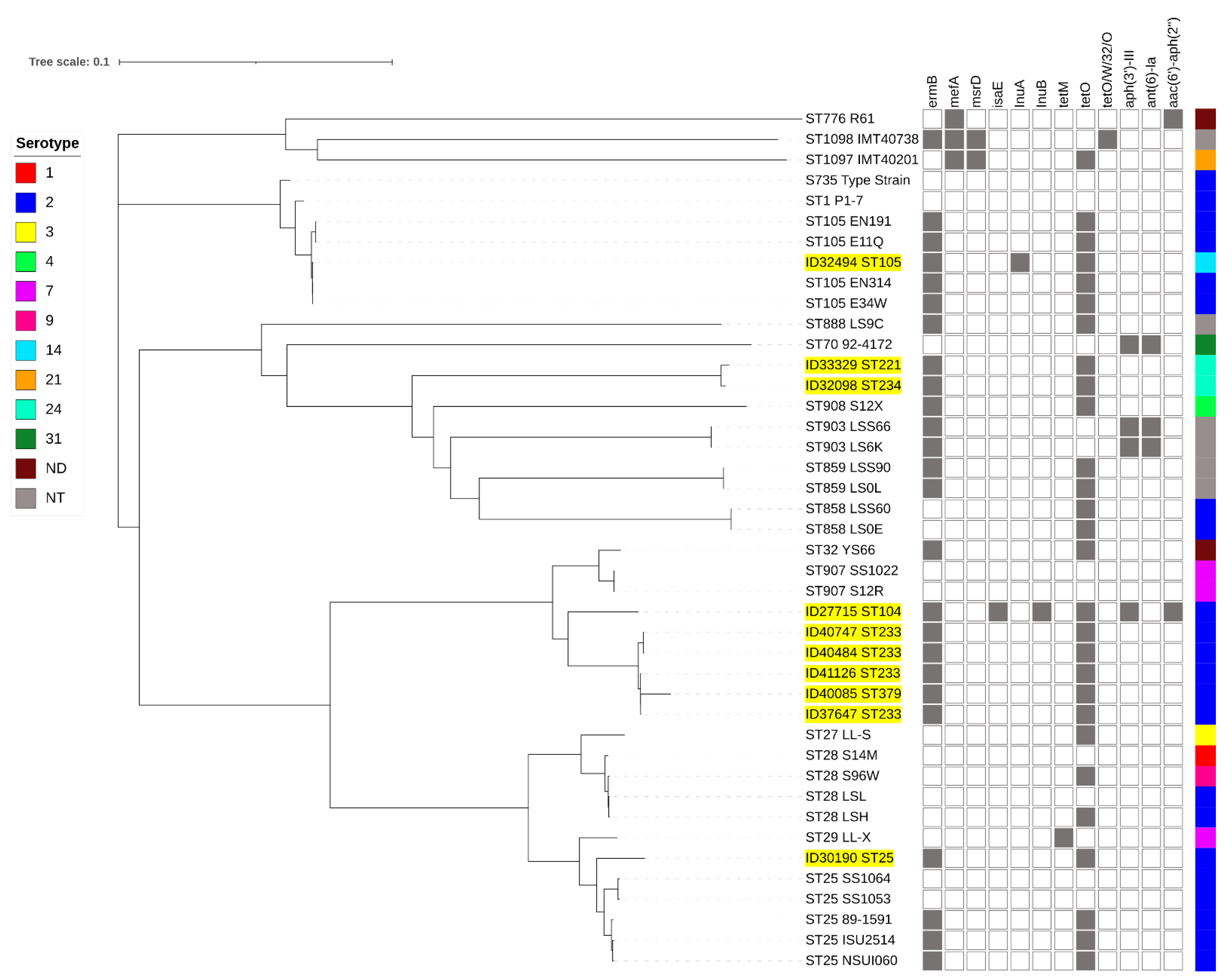
2.2. Macrolide- and Tetracycline-Resistance Genes
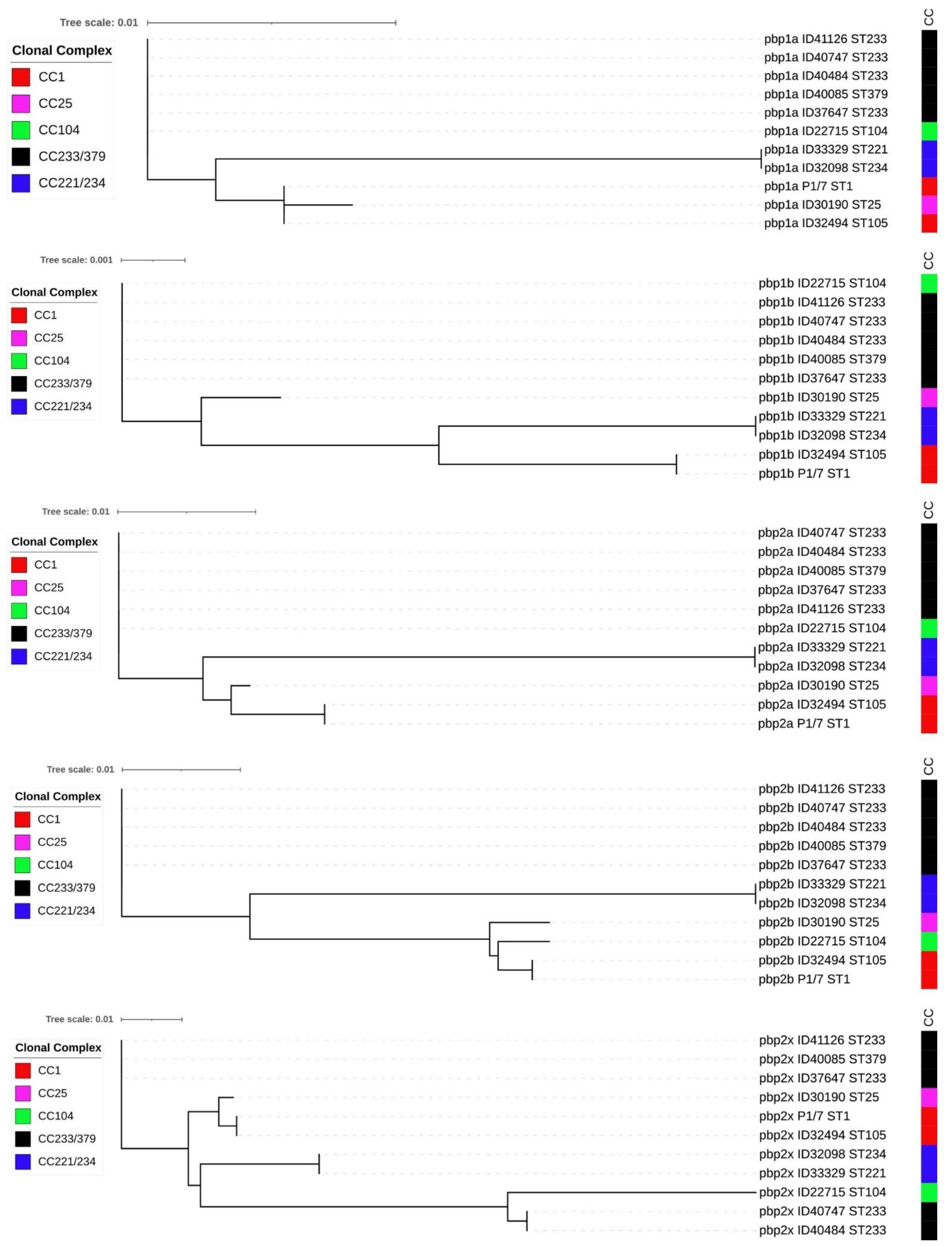
2.3. Whole-Genome Analysis of Non-penicillin-Susceptible S. suis Isolates
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Antimicrobial Susceptibility Testing
4.3. PCR Detection of Macrolide- and Tetracycline-Resistance Genes
4.4. Whole-Genome Sequencing (WGS)
4.5. Bioinformatics Analysis
4.6. Accession Numbers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent—An update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microbes. Infect. 2014, 3, e45. [Google Scholar] [CrossRef]
- Segura, M. Streptococcus suis research: Progress and challenges. Pathogens 2020, 9, 707. [Google Scholar] [CrossRef]
- Okura, M.; Osaki, M.; Nomoto, R.; Arai, S.; Osawa, R.; Sekizaki, T.; Takamatsu, D. Current Taxonomical situation of Streptococcus suis. Pathogens 2016, 24, E45. [Google Scholar] [CrossRef] [Green Version]
- Callejo, R.; Prieto, M.; Salamone, F.; Auger, J.P.; Goyette-Desjardins, G.; Gottschalk, M. Atypical Streptococcus suis in man, Argentina. Emerg. Infect. Dis. 2014, 20, 500–502. [Google Scholar] [CrossRef] [PubMed]
- Hatrongjit, R.; Kerdsin, A.; Gottschalk, M.; Takeuchi, D.; Hamada, S.; Oishi, K.; Akeda, Y. First human case report of sepsis due to infection with Streptococcus suis serotype 31 in Thailand. BMC Infect. Dis. 2015, 15, 392. [Google Scholar] [CrossRef]
- Kerdsin, A.; Oishi, K.; Sripakdee, S.; Boonkerd, N.; Polwichai, P.; Nakamura, S.; Uchida, R.; Sawanpanyalert, P.; Dejsirilert, S. Clonal dissemination of Streptococcus suis serotype 14 in Thailand. J. Med. Microbiol. 2009, 58, 1508–1513. [Google Scholar] [CrossRef] [PubMed]
- Kerdsin, A.; Dejsirilert, S.; Sawanpanyalert, P.; Boonnark, A.; Noithachang, W.; Sriyakum, D.; Simkum, S.; Chokngam, S.; Gottschalk, M.; Akeda, Y.; et al. Sepsis and spontaneous bacterial peritonitis in Thailand. Lancet 2011, 378, 960. [Google Scholar] [CrossRef]
- Kerdsin, A.; Hatrongjit, R.; Gottschalk, M.; Takeuchi, D.; Hamada, S.; Akeda, Y.; Oishi, K. Emergence of Streptococcus suis serotype 9 infection in humans. J. Microbiol. Immunol. Infect. 2017, 50, 545–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nghia, H.D.; Hoa, N.T.; le Linh, D.; Campbell, J.; Diep, T.S.; Chau, N.V.; Mai, N.T.; Hien, T.T.; Spratt, B.; Farrar, J.; et al. Human case of Streptococcus suis serotype 16 infection. Emerg. Infect. Dis. 2008, 14, 155–157. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, C.; Varaldo, P.E.; Facinelli, B. Streptococcus suis, an emerging drug-resistant animal and human pathogen. Front. Microbiol. 2011, 2, 235. [Google Scholar] [CrossRef] [Green Version]
- Segura, M.; Aragon, V.; Brockmeier, S.L.; Gebhart, C.; Greeff, A.; Kerdsin, A.; O’Dea, M.A.; Okura, M.; Saléry, M.; Schultsz, C.; et al. Update on Streptococcus suis research and prevention in the era of antimicrobial restriction: 4th International Workshop on S. suis. Pathogens 2020, 9, 374. [Google Scholar] [CrossRef]
- Gurung, M.; Tamang, M.D.; Moon, D.C.; Kim, S.R.; Jeong, J.H.; Jang, G.C.; Jung, S.C.; Park, Y.H.; Lim, S.K. Molecular basis of resistance to selected antimicrobial agents in the emerging zoonotic pathogen Streptococcus suis. J. Clin. Microbiol. 2015, 53, 2332–2336. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Garcia, J.; Wang, J.; Restif, O.; Holmes, M.A.; Mather, A.E.; Weinert, L.A.; Wileman, T.M.; Thomson, J.R.; Langford, P.R.; Wren, B.W.; et al. Patterns of antimicrobial resistance in Streptococcus suis isolates from pigs with or without streptococcal disease in England between 2009 and 2014. Vet Microbiol. 2017, 207, 117–124. [Google Scholar] [CrossRef]
- Ichikawa, T.; Oshima, M.; Yamagishi, J.; Muramatsu, C.; Asai, T. Changes in antimicrobial resistance phenotypes and genotypes in Streptococcus suis strains isolated from pigs in the Tokai area of Japan. J. Vet. Med. Sci. 2020, 82, 9–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matajira, C.E.C.; Moreno, L.Z.; Poor, A.P.; Gomes, V.T.M.; Dalmutt, A.C.; Parra, B.M.; Oliveira, C.H.; Barbosa, M.R.F.; Sato, M.I.Z.; Calderaro, F.F.; et al. Streptococcus suis in Brazil: Genotypic, virulence, and resistance profiling of strains isolated from pigs between 2001 and 2016. Pathogens 2020, 9, 31. [Google Scholar] [CrossRef] [Green Version]
- Yongkiettrakul, S.; Maneerat, K.; Arechanajan, B.; Malila, Y.; Srimanote, P.; Gottschalk, M.; Visessanguan, W. Antimicrobial susceptibility of Streptococcus suis isolated from diseased pigs, asymptomatic pigs, and human patients in Thailand. BMC Vet. Res. 2019, 15, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhang, P.; Wang, Y.; Fu, L.; Liu, L.; Xu, D.; Hou, Y.; Li, Y.; Fu, M.; Ding, S.; et al. Capsular serotypes, antimicrobial susceptibility, and the presence of transferable oxazolidinone resistance genes in Streptococcus suis isolated from healthy pigs in China. Vet. Microbiol. 2020, 247, 108750. [Google Scholar] [CrossRef] [PubMed]
- Bojarska, A.; Molska, E.; Janas, K.; Skoczyńska, A.; Stefaniuk, E.; Hryniewicz, W.; Sadowy, E. Streptococcus suis in invasive human infections in Poland: Clonality and determinants of virulence and antimicrobial resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 917–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoa, N.T.; Chieu, T.T.; Nghia, H.D.; Mai, N.T.; Anh, P.H.; Wolbers, M.; Baker, S.; Campbell, J.I.; Chau, N.V.; Hien, T.T.; et al. The antimicrobial resistance patterns and associated determinants in Streptococcus suis isolated from humans in southern Vietnam, 1997–2008. BMC Infect. Dis. 2011, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Marie, J.; Morvan, H.; Berthelot-Hérault, F.; Sanders, P.; Kempf, I.; Gautier-Bouchardon, A.V.; Jouy, E.; Kobisch, M. Antimicrobial susceptibility of Streptococcus suis isolated from swine in France and from humans in different countries between 1996 and 2000. J. Antimicrob. Chemother. 2002, 50, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Rayanakorn, A.; Ademi, Z.; Liew, D.; Lee, L.H. Burden of disease and productivity impact of Streptococcus suis infection in Thailand. PLoS Negl. Trop. Dis. 2021, 15, e0008985. [Google Scholar] [CrossRef] [PubMed]
- Dechêne-Tempier, M.; Marois-Créhan, C.; Libante, V.; Jouy, E.; Leblond-Bourget, N.; Payot, S. Update on the Mechanisms of Antibiotic Resistance and the Mobile Resistome in the Emerging Zoonotic Pathogen Streptococcus suis. Microorganisms 2021, 9, 1765. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, Z.; Song, L.; Fan, X.; Wen, F.; Xu, S.; Ning, Y. Antimicrobial resistance profile and genotypic characteristics of Streptococcus suis capsular type 2 isolated from clinical carrier sows and diseased pigs in China. Biomed. Res. Int. 2015, 2015, 284303. [Google Scholar] [CrossRef] [PubMed]
- van Hout, J.; Heuvelink, A.; Gonggrijp, M. Monitoring of antimicrobial susceptibility of Streptococcus suis in the Netherlands, 2013–2015. Vet. Microbiol. 2016, 194, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.P.; Saegerman, C.; Douny, C.; Dinh, T.V.; Xuan, B.H.; Vu, B.D.; Hong, N.P.; Scippo, M.L. First survey on the use of antibiotics in pig and poultry production in the Red Delta region of Vietnam. Food Pub. Health 2013, 3, 247–256. [Google Scholar] [CrossRef]
- Pyörälä, S.; Baptiste, K.E.; Catry, B.; van Duijkeren, E.; Greko, C.; Moreno, M.A.; Pomba, M.C.; Rantala, M.; Ružauskas, M.; Sanders, P.; et al. Macrolides and lincosamides in cattle and pigs: Use and development of antimicrobial resistance. Vet. J. 2014, 200, 230–239. [Google Scholar] [CrossRef]
- Chen, L.; Song, Y.; Wei, Z.; He, H.; Zhang, A.; Jin, M. Antimicrobial susceptibility, tetracycline and erythromycin resistance genes, and multilocus sequence typing of Streptococcus suis isolates from diseased pigs in China. J. Vet. Med. Sci. 2013, 75, 583–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yongkiettrakul, S.; Wongsurawat, T.; Jenjaroenpun, P.; Acheampong, D.A.; Srimanote, P.; Maneerat, K.; Visessanguan, W.; Nookaew, I. Genome sequences of antibiotic-resistant Streptococcus suis strains isolated from human patients and diseased and asymptomatic pigs in Thailand. Infect. Genet. Evol. 2021, 87, 104674. [Google Scholar] [CrossRef] [PubMed]
- Werinder, A.; Aspán, A.; Backhans, A.; Sjölund, M.; Guss, B.; Jacobson, M. Streptococcus suis in Swedish grower pigs: Occurrence, serotypes, and antimicrobial susceptibility. Acta Vet. Scand. 2020, 62, 36. [Google Scholar] [CrossRef]
- Niemann, L.; Eichhorn, I.; Müller, P.; Brauns, J.; Nathaus, R.; Schäkel, F.; Höltig, D.; Wendt, M.; Kadlec, K.; Schwarz, S. Draft genome sequences of three porcine Streptococcus suis isolates which differ in their susceptibility to penicillin. Microbiol. Resour. Announc. 2019, 8, e01711-18. [Google Scholar] [CrossRef] [Green Version]
- Hu, P.; Yang, M.; Zhang, A.; Wu, J.; Chen, B.; Hua, Y.; Yu, J.; Chen, H.; Xiao, J.; Jin, M. Comparative genomics study of multi-drug-resistance mechanisms in the antibiotic-resistant Streptococcus suis R61 strain. PLoS ONE 2011, 6, e24988. [Google Scholar] [CrossRef] [Green Version]
- Hakenbeck, R.; Grebe, T.; Zähner, D.; Stock, J.B. β-lactam resistance in Streptococcus pneumoniae: Penicillin-binding proteins and non-penicillin-binding proteins. Mol. Microbiol. 1999, 33, 673–678. [Google Scholar] [CrossRef]
- Haenni, M.; Moreillon, P. Mutations in penicillin-binding protein (PBP) genes and in non-PBP genes during selection of penicillin-resistant Streptococcus gordonii. Antimicrob. Agents Chemother. 2006, 50, 4053–4061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fani, F.; Leprohon, P.; Légaré, D.; Ouellette, M. Whole genome sequencing of penicillin-resistant Streptococcus pneumoniae reveals mutations in penicillin-binding proteins and in a putative iron permease. Genome. Biol. 2011, 12, R115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okura, M.; Auger, J.P.; Shibahara, T.; Goyette-Desjardins, G.; Van Calsteren, M.R.; Maruyama, F.; Kawai, M.; Osaki, M.; Segura, M.; Gottschalk, M.; et al. Capsular polysaccharide switching in Streptococcus suis modulates host cell interactions and virulence. Sci. Rep. 2021, 11, 6513. [Google Scholar] [CrossRef] [PubMed]
- Kerdsin, A.; Dejsirilert, S.; Puangpatra, P.; Sripakdee, S.; Chumla, K.; Boonkerd, N.; Polwichai, P.; Tanimura, S.; Takeuchi, D.; Nakayama, T.; et al. Genotypic profile of Streptococcus suis serotype 2 and clinical features of infection in humans, Thailand. Emerg. Infect. Dis. 2011, 17, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Kerdsin, A.; Akeda, Y.; Takeuchi, D.; Dejsirilert, S.; Gottschalk, M.; Oishi, K. Genotypic diversity of Streptococcus suis strains isolated from humans in Thailand. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 917–925. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Document M100-S30; Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2020. [Google Scholar]
- Kang, Y.; Li, Q.; Yin, Z.; Shen, M.; Zhao, H.; Bai, Y.; Mei, L.; Hu, J. High diversity and abundance of cultivable tetracycline-resistant bacteria in soil following pig manure application. Sci. Rep. 2018, 8, 1489. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic. Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vielva, L.; de Toro, M.; Lanza, V.F.; de la Cruz, F. PLACNETw: A web-based tool for plasmid reconstruction from bacterial genomes. Bioinformatics 2017, 33, 3796–3798. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic. Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Feng, Y.; Zou, S.; Chen, H.; Yu, Y.; Ruan, Z. BacWGSTdb 2.0: A one-stop repository for bacterial whole-genome sequence typing and source tracking. Nucleic. Acids Res. 2021, 49, D644–D650. [Google Scholar] [CrossRef]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the problem of comparing whole bacterial genomes across different sequencing platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Serotypes | CC | ST | MIC (µg/mL) | No. of Isolates. |
|---|---|---|---|---|
| 2 | 1 | 1 | 0.38 | 2 |
| 25 | 25 | 0.38 | 1 | |
| 104 | 104 | 0.25 | 3 | |
| 1 | 105 | 0.38 | 1 | |
| 233/379 | 233 | 0.25 | 1 | |
| 0.38 | 7 | |||
| 0.5 | 9 | |||
| 0.75 | 3 | |||
| 1 | 1 | |||
| 1.5 | 1 | |||
| 379 | 0.25 | 1 | ||
| 0.5 | 1 | |||
| 0.75 | 1 | |||
| 104 | 393 | 0.5 | 1 | |
| 5 | - | 235 | 1 | 1 |
| 14 | 1 | 105 | 0.5 | 1 |
| 24 | 221/234 | 221 | 0.75 | 1 |
| 234 | 0.38 | 1 | ||
| Total | 37 | |||
| Serotype | No. of Isolates | Macrolide-Resistance Genes | Tetracycline-Resistance Genes | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| ermB | mefA | msrD | Unknown | tetO | tetM | tetM+ tetL | tetW | tetO+ tetW | Unknown | ||
| 1 | 2 (0.45%) | 2 (0.45%) | - | - | - | 2 (0.45%) | 0 | 0 | 0 | 0 | 0 |
| 2 | 392 (87.50%) | 334 (74.55%) | - | - | 58 (12.95%) | 357 (79.69%) | 1 (0.22%) | 1 (0.22%) | - | 26 (5.80%) | 7 (1.56%) |
| 4 | 1 (0.22%) | 1 (0.22%) | - | - | - | 1 (0.22%) | 0 | 0 | 0 | 0 | 0 |
| 5 | 2 (0.45%) | 2 (0.45%) | - | - | - | 2 (0.45%) | 0 | 0 | 0 | 0 | 0 |
| 9 | 1 (0.22%) | - | - | - | 1 (0.22%)- | 0 | 0 | 0 | 1 (0.22%) | 0 | 0 |
| 14 | 47 (10.49%) | 42 (9.37%) | - | - | 5 (1.12%) | 44 (9.82%) | - | 1 (0.22%) | - | - | 2 (0.45%) |
| 24 | 3 (0.67%) | 3 (0.67%) | - | - | - | 3 (0.67%) | 0 | 0 | 0 | 0 | 0 |
| Total | 448 (100%) | 384 (85.71%) | 0 | 0 | 64 (14.29%) | 409 (91.29%) | 1 (0.22%) | 2 (0.45%) | 1 (0.22%) | 26 (5.8%) | 9 (2.0%) |
| Serotype | CC | ST | Strain ID No. | Plasmid Replicon Types | Acquired Antimicrobial-Resistance Genes | Substitutions in pbp Amino Acid | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ML | TET | AMG | pbp1a | pbp1b | pbp2a | pbp2b | pbp2x | |||||
| 2 | 25 | 25 | 30190 | MOBV, MOBT | ermB | tetO | - | T249P, A365S | G18V, I267V, K372G, A419S, V492I, T578A A765P | H27R, I121V, N233D, A275T, A561S, S660T | I414V, V637M, V641I | I72T, D136A, L204P, A468S, |
| 104 | 104 | 27715 | MOBP, MOBT | ermB, InuB, IsaE | tetO | aac(6′)-aph(2″), aph(3′)-III | E447A, A493T, T689A, A703S | G18V, I267V, K372G, A419S, G476R, T578A A765P | H19N, R22Q, H27R, I34V, I121V, N233D V390I, A434S, A561S, S637N, S660T | K143E, I414V, V641I, A659V | I72T, L109F, K110Q, K111E, V112I, Q116E, S121H, E125D, F127L, K128N, H130Y, M139N, E140K, E146D, S165T, E177K, V205I, V217M, L220M, V245I, K251S, T254Q, T276I, N284T, R288K, Q321E, Y382Q, Y389F, Q407E, T418A, F422Y, S450T, I474V, V494I, S495A, S496A, Y525F, L533V, D541E, V547M, T551S, S556G, Q560L, G565D, I568T, N595S, D601T, S621V, P624S, A625N, A627S, R628L, V629L, A630T, Q636K, L640I, I641V, S648I, Q655N, A666S, K670Q, V671A, S677T, K678A, I680S, I701V, I708L, F709I, K720E, T722Q, S724A, V727I, A736K, K742N, I745V | |
| 233/379 | 233 | 40747 | MOBP, MOBT | ermB | tetO | - | E447A, A493T, T689A, A703S | G18V, I267V, K372G, A419S, G476R, T578A A765P | H19N, R22Q, H27R, I34V, I121V, N233D V390I, A434S, A561S, S637N, S660T | P92S, K143E, N179S, S206A, I300V, R332K N356E, S374A, S375G, T376S, I414V, Y432W I452A, K479T, T507I, D512E, K513E, T515S D587E, T625R, K674N | I72T, L109F, K110Q, K111E, V112I, Q116E, S121H, E125D, F127L, K128N, H130Y, M139N, E140K, E146D, S165T, E177K, V205I, V217M, L220M, V245I, K251S, T254Q, N284T, R288K, Q321E, Y382Q, Y389F, Q407E, T418A, F422Y, S450T, I474V, V494I, S495A, S496A, Y525F, L533V, D541E, V547M, T551S, S556G, Q560L, G565D, I568T, R628Q, K678N, K741E | |
| 41126 | MOBP, MOBT | ermB | tetO | - | E447A, A493T, T689A, A703S | G18V, I267V, K372G, A419S, G476R, T578A A765P | H19N, R22Q, H27R, I34V, I121V, N233D V390I, A434S, A561S, S637N, S660T | P92S, K143E, N179S, S206A, I300V, R332K N356E, S374A, S375G, T376S, I414V, Y432W I452A, K479T, T507I, D512E, K513E, T515S D587E, T625R, K674N | V18I, I72T, E146K, N182D, P190S, L204P, H212Y, S450T, D486N, T551S, I568T, R628Q, K678N, K741E | |||
| 40484 | MOBP, MOBT | ermB | tetO | - | E447A, A493T, T689A, A703S | G18V, I267V, K372G, A419S, G476R, T578A A765P | H19N, R22Q, H27R, I34V, I121V, N233D V390I, A434S, A561S, S637N, S660T | P92S, K143E, N179S, S206A, I300V, R332K N356E, S374A, S375G, T376S, I414V, Y432W I452A, K479T, T507I, D512E, K513E, T515S D587E, T625R, K674N | I72T, L109F, K110Q, K111E, V112I, Q116E, S121H, E125D, F127L, K128N, H130Y, M139N, E140K, E146D, S165T, E177K, V205I, V217M, L220M, V245I, K251S, T254Q, N284T, R288K, Q321E, Y382Q, Y389F, Q407E, T418A, F422Y, S450T, I474V, V494I, S495A, S496A, Y525F, L533V, D541E, V547M, T551S, S556G, Q560L, G565D, I568T, R628Q, K678N, K741E | |||
| 37647 | MOBP, MOBT | ermB | tetO | - | E447A, A493T, T689A, A703S | G18V, I267V, K372G, A419S, G476R, T578A A765P | H19N, R22Q, H27R, I34V, I121V, N233D V390I, A434S, A561S, S637N, S660T | P92S, K143E, N179S, S206A, I300V, R332K N356E, S374A, S375G, T376S, I414V, Y432W I452A, K479T, T507I, D512E, K513E, T515S D587E, T625R, K674N | V18I, I72T, E146K, N182D, P190S, L204P, H212Y, S450T, D486N, T551S, I568T, R628Q, K678N, K741E | |||
| 379 | 40085 | MOBP, MOBT | ermB | tetO | - | E447A, A493T, T689A, A703S | G18V, I267V, K372G, A419S, G476R, T578A A765P | H19N, R22Q, H27R, I34V, I121V, N233D V390I, A434S, A561S, S637N, S660T | P92S, K143E, N179S, S206A, I300V, R332K N356E, S374A, S375G, T376S, I414V, Y432W I452A, K479T, T507I, D512E, K513E, T515S D587E, T625R, K674N | V18I, I72T, E146K, N182D, P190S, L204P, H212Y, S450T, D486N, T551S, I568T, R628Q, K678N, K741E | ||
| 14 | 1 | 105 | 32494 | Rep21, MOBT | ermB, InuA | tetO | - | - | - | - | - | - |
| 24 | 221/234 | 221 | 33329 | MOBP, MOBV | ermB | tetO | - | A231V, A289E, P409T, V412F, N459D, H464Y S477G, K479Q, K522Q, K525Q, N550D, S578A S675Y, T689A, A703S | S259A, K317E, K338E, K372G, T578A, A765P S779T | H27R, I34V, I121V, N233D, R379T, S430A M433T, A434P, L479F, Y487F, V504A, A522S T523A, K527D, E549Q, A561S, A568S, T584A A585T, T578L, T581S, H588Y, I589L, H610Y S638G, S644T, Q647R, N651D, S652A, Y656N S660T, K661S, E663D, A669V, E698D | P92S, K143E, N179S, S206A, T231N, I240V S266P, Q291E, I300V, E304K, R332K, A340S V408I, I414V, Q415L, Y432W, I452V, K479T D512E, K513E, T515S, T521M, A539V, I542V T547K, A562P, E568D, E586D, D587E, V598L S602G, A633P, D636E, Q668E, S669N, T675A Q678T, Q685N, H688Y | I72T, D136A, A195S, L204P, T418A, A468S, T551S, I568T, D579N, N595S, D601S, V603I, S621A, A625S, A627T, R628Y, A630E, N631K, E634D, Q636K, K678N |
| 234 | 32098 | MOBP, MOBV | ermB | tetO | - | A231V, A289E, P409T, V412F, N459D, H464Y S477G, K479Q, K522Q, K525Q, N550D, S578A S675Y, T689A, A703S | S259A, K317E, K338E, K372G, T578A, A765P S779T | H27R, I34V, I121V, N233D, R379T, S430A M433T, A434P, L479F, Y487F, V504A, A522S T523A, K527D, E549Q, A561S, A568S, T584A A585T, T578L, T581S, H588Y, I589L, H610Y S638G, S644T, Q647R, N651D, S652A, Y656N S660T, K661S, E663D, A669V, E698D | P92S, K143E, N179S, S206A, T231N, I240V S266P, Q291E, I300V, E304K, R332K, A340S V408I, I414V, Q415L, Y432W, I452V, K479T D512E, K513E, T515S, T521M, A539V, I542V T547K, A562P, E568D, E586D, D587E, V598L S602G, A633P, D636E, Q668E, S669N, T675A Q678T, Q685N, H688Y | I72T, D136A, A195S, L204P, T418A, A468S, T551S, I568T, D579N, N595S, D601S, V603I, S621A, A625S, A627T, R628Y, A630E, N631K, E634D, Q636K, K678N | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bamphensin, N.; Chopjitt, P.; Hatrongjit, R.; Boueroy, P.; Fittipaldi, N.; Gottschalk, M.; Kerdsin, A. Non-Penicillin-Susceptible Streptococcus suis Isolated from Humans. Pathogens 2021, 10, 1178. https://doi.org/10.3390/pathogens10091178
Bamphensin N, Chopjitt P, Hatrongjit R, Boueroy P, Fittipaldi N, Gottschalk M, Kerdsin A. Non-Penicillin-Susceptible Streptococcus suis Isolated from Humans. Pathogens. 2021; 10(9):1178. https://doi.org/10.3390/pathogens10091178
Chicago/Turabian StyleBamphensin, Nichari, Peechanika Chopjitt, Rujirat Hatrongjit, Parichart Boueroy, Nahuel Fittipaldi, Marcelo Gottschalk, and Anusak Kerdsin. 2021. "Non-Penicillin-Susceptible Streptococcus suis Isolated from Humans" Pathogens 10, no. 9: 1178. https://doi.org/10.3390/pathogens10091178