
Evaluation of the Correlation between the mRNA Expression Levels of ystA and ymoA Genes in Y. enterocolitica Strains with Different Enterotoxic Properties


Abstract
:1. Introduction
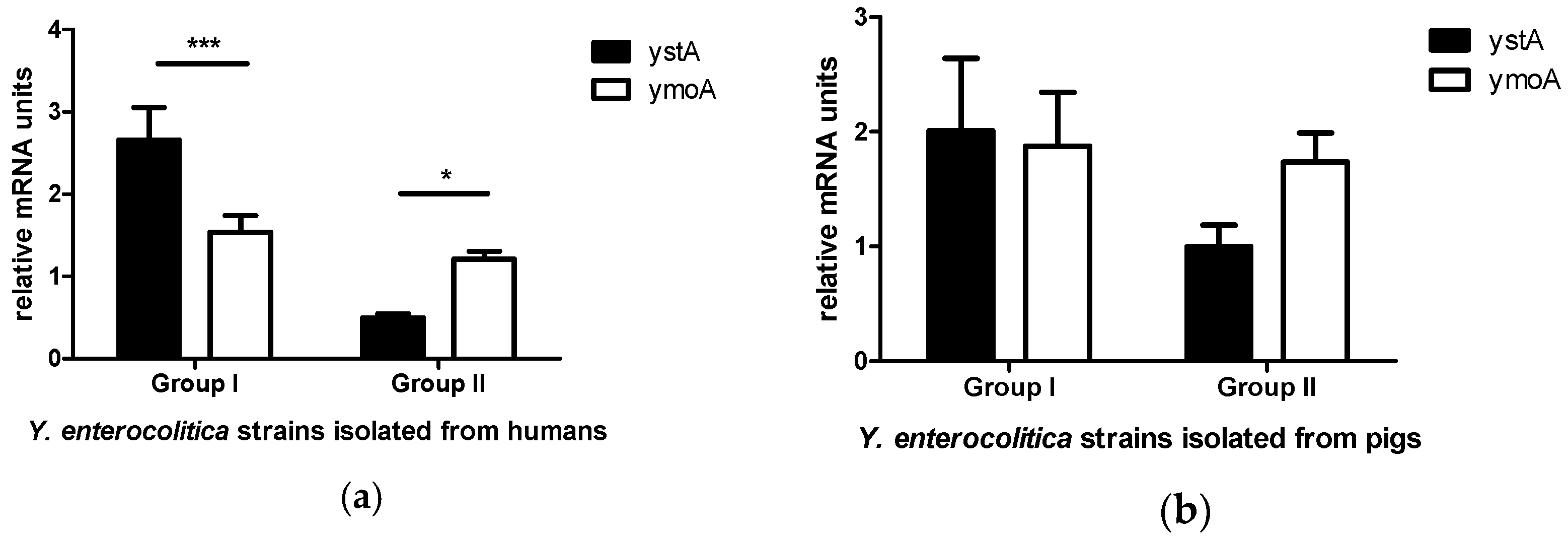
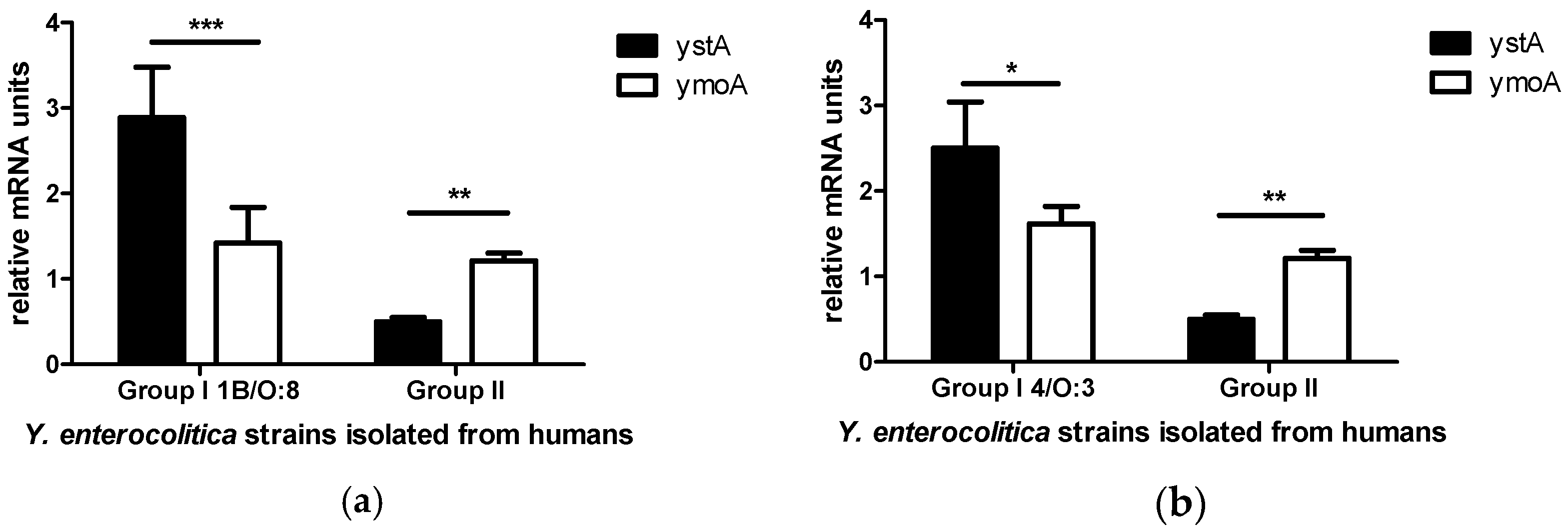
2. Results
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Yersinia enterocolitica Strains Isolated from Humans
4.3. Yersinia enterocolitica Strains Isolated from Pigs
4.4. RNA Preparation and Reverse Transcription
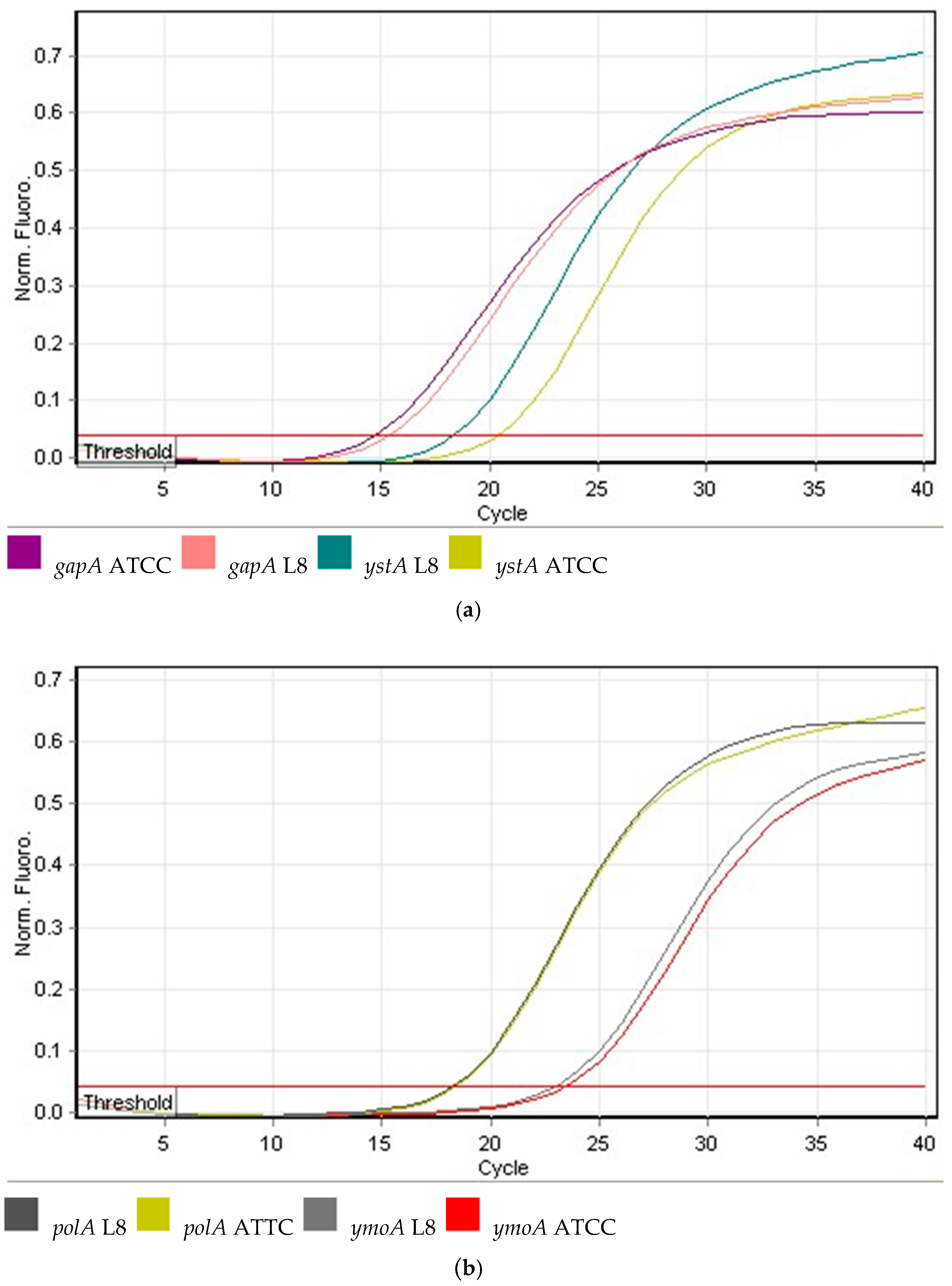
4.5. Gene Expression Analysis Using qPCR
4.6. Data Analysis and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bottone, E.J. Yersinia enterocolitica: Revisitation of an Enduring Human Pathogen. Clin. Microbiol. Newsl. 2015, 37, 1–8. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority and European Centre for Disease Prevention and Control). The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, 6406. [Google Scholar]
- Saraka, D.; Savin, C.; Kouassi, S.; Cissé, B.; Koffi, E.; Cabanel, N.; Brémont, S.; Faye-Kette, H.; Dosso, M.; Carniel, E. Yersinia enterocolitica, a Neglected Cause of Human Enteric Infections in Côte d’Ivoire. PLoS Negl. Trop. Dis. 2017, 11, e0005216. [Google Scholar]
- Chlebicz, A.; Śliżewska, K. Campylobacteriosis, Salmonellosis, Yersiniosis, and Listeriosis as Zoonotic Foodborne Diseases: A Review. Int. J. Environ. Res. Public Health 2018, 15, 863. [Google Scholar] [CrossRef] [Green Version]
- Savin, C.; Le Guern, A.S.; Lefranc, M.; Brémont, S.; Carniel, E.; Pizarro-Cerdá, J. Isolation of a Yersinia enterocolitica biotype 1B strain in France, and evaluation of its genetic relatedness to other European and North American biotype 1B strains. Emerg. Microbes Infect. 2018, 7, 121. [Google Scholar] [CrossRef] [Green Version]
- Cilia, G.; Turchi, B.; Fratini, F.; Bilei, S.; Bossù, T.; De Marchis, M.L.; Cerri, D.; Pacini, M.I.; Bertelloni, F. Prevalence, Virulence and Antimicrobial Susceptibility of Salmonella spp., Yersinia enterocolitica and Listeria monocytogenes in European Wild Boar (Sus scrofa) Hunted in Tuscany (Central Italy). Pathogens 2021, 10, 93. [Google Scholar] [CrossRef]
- Modesto, P.; De Ciucis, C.G.; Vencia, W.; Pugliano, M.C.; Mignone, W.; Berio, E.; Masotti, C.; Ercolini, C.; Serracca, L.; Andreoli, T.; et al. Evidence of Antimicrobial Resistance and Presence of Pathogenicity Genes in Yersinia enterocolitica Isolate from Wild Boars. Pathogens 2021, 10, 398. [Google Scholar] [CrossRef]
- Råsbäck, T.; Rosendal, T.; Stampe, M.; Sannö, A.; Aspán, A.; Järnevi, K.; Lahti, E.T. Prevalence of human pathogenic Yersinia enterocolitica in Swedish pig farms. Acta Vet. Scand. 2018, 25, 39. [Google Scholar] [CrossRef]
- Morka, K.; Wałecka-Zacharska, E.; Schubert, J.; Dudek, B.; Woźniak-Biel, A.; Kuczkowski, M.; Wieliczko, A.; Bystroń, J.; Bania, J.; Bugla-Płoskońska, G. Genetic Diversity and Distribution of Virulence-Associated Genes in Y. enterocolitica and Y. enterocolitica-Like Isolates from Humans and Animals in Poland. Pathogens 2021, 10, 65. [Google Scholar] [CrossRef]
- Farzan, A.; Friendship, R.M.; Cook, A.; Pollari, F. Occurrence of Salmonella, Campylobacter, Yersinia enterocolitica, Escherichia coli O157 and Listeria monocytogenes in Swine. Zoonoses Public Health 2010, 57, 388–396. [Google Scholar] [CrossRef]
- Singh, I.; Virdi, J.S. Production of Yersinia stable toxin (YST) and distribution of yst genes in biotype 1A strains of Yersinia enterocolitica. J. Med. Microbiol. 2004, 53, 1065–1068. [Google Scholar] [CrossRef] [Green Version]
- Delor, I.; Kaeckenbeeck, A.; Wauters, G.; Cornelis, G.R. Nucleotide sequence of yst, the Yersinia enterocolitica gene encoding the heat-stable enterotoxin, and prevalence of the gene among pathogenic and nonpathogenic yersiniae. Infect. Immun. 1990, 58, 2983–2988. [Google Scholar] [CrossRef] [Green Version]
- Revell, P.A.; Miller, V.L. Yersinia virulence: More than a plasmid. FEMS Microbiol. Lett. 2001, 205, 159–164. [Google Scholar] [CrossRef]
- Bancerz-Kisiel, A.; Szczerba-Turek, A.; Platt-Samoraj, A.; Szweda, W. Distribution of the ymoA and ystA genes and enterotoxins Yst production by Yersinia enterocolitica strains isolated from humans and pigs. Pol. J. Vet. Sci. 2012, 15, 609–614. [Google Scholar] [CrossRef] [Green Version]
- Mikulskis, A.V.; Delor, I.; Ha Thi, V.; Cornelis, G.R. Regulation of the Yersinia enterocolitica enterotoxin Yst gene. Influence of growth phase, temperature, osmolarity, pH and bacterial host factors. Mol. Microbiol. 1994, 14, 905–915. [Google Scholar] [CrossRef]
- Huovinien, E.; Shivonen, L.M.; Virtanen, M.J.; Haukka, K.; Siitinen, A.; Kuusi, M. Symptoms and sources of Yersinia enterocolitica—Infection: A case control study. BMC Infect. Dis. 2010, 10, 122. [Google Scholar] [CrossRef] [Green Version]
- Böhme, K.; Steinmann, R.; Kortmann, J.; Seekircher, S.; Heroven, A.K.; Berger, E.; Pisano, F.; Thiermann, T.; Wolf-Watz, H.; Narberhaus, F.; et al. Concerted Actions of a Thermo-labile Regulator and a Unique Intergenic RNA Thermosensor Control Yersinia Virulence. PLoS Pathog. 2012, 8, 2. [Google Scholar] [CrossRef] [Green Version]
- Mourino, M.; Madrid, C.; Balsalobre, C.; Prenafeta, A.; Munoa, F.; Blanco, J.; Blanco, M.; Blanco, J.E.; Juarez, A. The Hha protein as a modulator of expression of virulence factors in Escherichia coli. Infect. Immun. 1996, 64, 2881–2884. [Google Scholar] [CrossRef] [Green Version]
- Carmona, M.; Balsalobre, C.; Munoa, F.; Mourino, M.; Jubete, Y.; De la Cruz, F.; Juárez, A. Escherichia coli hha mutants, dna supercoiling and expression of the haemolysin genes from the recombinant plasmid pann202–312. Mol. Microbiol. 1993, 9, 1011–1018. [Google Scholar] [CrossRef]
- Madrid, C.; Balsalobre, C.; Garcia, J.; Juarez, A. The novel Hha/YmoA family of nucleoid-associated proteins: Use of structural mimicry to modulate the activity of the H-NS family of proteins. Mol. Microbiol. 2007, 63, 7–14. [Google Scholar] [CrossRef]
- Nieto, J.M.; Madrid, C.; Miquelay, E.; Parra, J.L.; Rodriguez, S.; Juarez, A. Evidence for direct protein-protein interaction between members of the enterobacterial Hha/YmoA and H-NS families of proteins. J. Bacteriol. 2002, 184, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Dorman, C.J. H-NS: A universal regulator for a dynamic genome. Nat. Rev. Microbiol. 2004, 2, 391–400. [Google Scholar] [CrossRef]
- Rimsky, S. Structure of the histone-like protein H-NS and its role in regulation and genome superstructure. Curr. Opin. Microbiol. 2004, 7, 109–114. [Google Scholar] [CrossRef]
- Peruzy, M.F.; Murru, N.; Perugini, A.G.; Capuano, F.; Delibato, E.; Mercogliano, R.; Korkeala, H.; Proroga, Y.T.R. Evaluation of virulence genes in Yersinia enterocolitica strains using SYBR Green real-time PCR. Food Microbiol. 2017, 65, 231–235. [Google Scholar] [CrossRef]
- Baños, R.C.; Pons, J.I.; Madrid, C.; Juárez, A. A global modulatory role for the Yersinia enterocolitica H-NS protein. Microbiol. 2008, 154, 1281–1289. [Google Scholar] [CrossRef] [Green Version]
- Madrid, C.; Nieto, J.M.; Juárez, A. Role of the Hha/YmoA family of proteins in the thermoregulation of the expression of virulence factors. Int. J. Med. Microbiol. 2001, 291, 425–432. [Google Scholar] [CrossRef]
- Cornelis, G.R.; Sluiters, C.; Delor, I.; Geib, D.; Kaniga, K.; De Rouvroit, C.L.; Sory, M.-P.; VanOoteghem, J.-C.; Michiels, T.; Cornells, G.R. ymoA, a Yersinia enterocolitica chromosomal gene modulating the expression of virulence functions. Mol. Microbiol. 1991, 5, 1023–1034. [Google Scholar] [CrossRef] [PubMed]
- Ellison, D.W.; Young, B.; Nelson, K.; Miller, V.L. YmoA Negatively Regulates Expression of Invasin from Yersinia enterocolitica. J. Bacteriol. 2003, 185, 7153–7159. [Google Scholar] [CrossRef] [Green Version]
- Bancerz-Kisiel, A.; Lipczyńska, K.; Szczerba-Turek, A.; Gospodarek, E.; Platt-Samoraj, A.; Szweda, W. The use of the HRM method for identifying possible mutations in the ymoA gene region and evaluating their influence on the enterotoxic properties of Y. enterocolitica strains. BMC Vet. Res. 2014, 10, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Grant, T.; Bennett-Wood, V.; Robins-Browne, R.M. Identification of Virulence-Associated Characteristics in Clinical Isolates of Yersinia enterocolitica Lacking Classical Virulence Markers. Infect. Immun. 1998, 66, 1113–1120. [Google Scholar] [CrossRef] [Green Version]
- Starke, M.; Fuchs, T.M. YmoA negatively controls the expression of insecticidal genes inYersinia enterocolitica. Mol. Microbiol. 2014, 92, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Heroven, A.K.; Dersch, P.; Tran-Winkler, H.; Böhme, K. Regulatory Elements Implicated in the Environmental Control of Invasin Expression in Enteropathogenic Yersinia. Adv. Exp. Med. Biol. 2007, 603, 156–166. [Google Scholar] [CrossRef]
- Ellison, D.W.; Miller, V.L. H-NS Represses inv Transcription in Yersinia enterocolitica through Competition with RovA and Interaction with YmoA. J. Bacteriol. 2006, 188, 5101–5112. [Google Scholar] [CrossRef] [Green Version]
- Giannella, R.A. Suckling mouse model for detection of heat-stable Escherichia coli enterotoxin: Characteristics of the model. Infect. Immun. 1976, 14, 95–99. [Google Scholar] [CrossRef] [Green Version]
- Brzostek, K.; Brzóstkowska, M.; Bukowska, I.; Karwicka, E.; Raczkowska, A. OmpR negatively regulates expression of invasion in Yersinia enterocolitica. Microbiology 2007, 153, 2416–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platt-Samoraj, A.; Ugorski, M.; Szweda, W.; Szczerba-Turek, A.; Wojciech, Ł.; Procajło, Z. Analysis of the presence of ail, ystA and ystB genes in Yersinia enterocolitica strains isolated from aborting sows and aborted fetuses. J. Vet. Med. B 2007, 53, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Townsend, M.K.; Carr, N.J.; Iyer, J.G.; Horne, S.M.; GibbS, P.S.; Prüß, B.M. Pleiotropic phenotypes of a Yersinia enterocolitica flhD mutant include reduced lethality in a chicken embryo model. BMC Microbiol. 2008, 8, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Pleiotropic phenotypes of a Yersinia enterocolitica flhD mutant include reduced lethality in a chicken embryo model. Nat. Protocols 2008, 3, 1101–1108. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Reference |
|---|---|---|---|
| ystA | 5′GTCTTCATTTGGAGGATTCGGC3′ | 5′AATCACTACTGACTTCGGCTGG3′ | Platt-Samoraj et al. [36] |
| ymoA | 5′GACTTTTCTCAGGGGAATAC3′ | 5′GCTCAACGTTGTGTGTCT3′ | Grant et al. [30] |
| polA | 5′-GCTGGCTTGCGGATGTAGAT-3′ | 5′-AGCACGGCGGTCACTTCA-3′ | Townsend et al. [37] |
| gapA | 5′-CCATCCGTGTTACCGCAGAG-3′ | 5′-TCTTAGCACCAGCAGCAATGT-3′ | Townsend et al. [37] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bancerz-Kisiel, A.; Lipczyńska-Ilczuk, K. Evaluation of the Correlation between the mRNA Expression Levels of ystA and ymoA Genes in Y. enterocolitica Strains with Different Enterotoxic Properties. Pathogens 2021, 10, 1136. https://doi.org/10.3390/pathogens10091136
Bancerz-Kisiel A, Lipczyńska-Ilczuk K. Evaluation of the Correlation between the mRNA Expression Levels of ystA and ymoA Genes in Y. enterocolitica Strains with Different Enterotoxic Properties. Pathogens. 2021; 10(9):1136. https://doi.org/10.3390/pathogens10091136
Chicago/Turabian StyleBancerz-Kisiel, Agata, and Karolina Lipczyńska-Ilczuk. 2021. "Evaluation of the Correlation between the mRNA Expression Levels of ystA and ymoA Genes in Y. enterocolitica Strains with Different Enterotoxic Properties" Pathogens 10, no. 9: 1136. https://doi.org/10.3390/pathogens10091136