
Evidence of Extensive Circulation of Yersinia enterocolitica in Rodents and Shrews in Natural Habitats from Retrospective and Perspective Studies in South Caucasus

 ,
,
Abstract
:1. Introduction
2. Material & Methods
2.1. Archived Documents
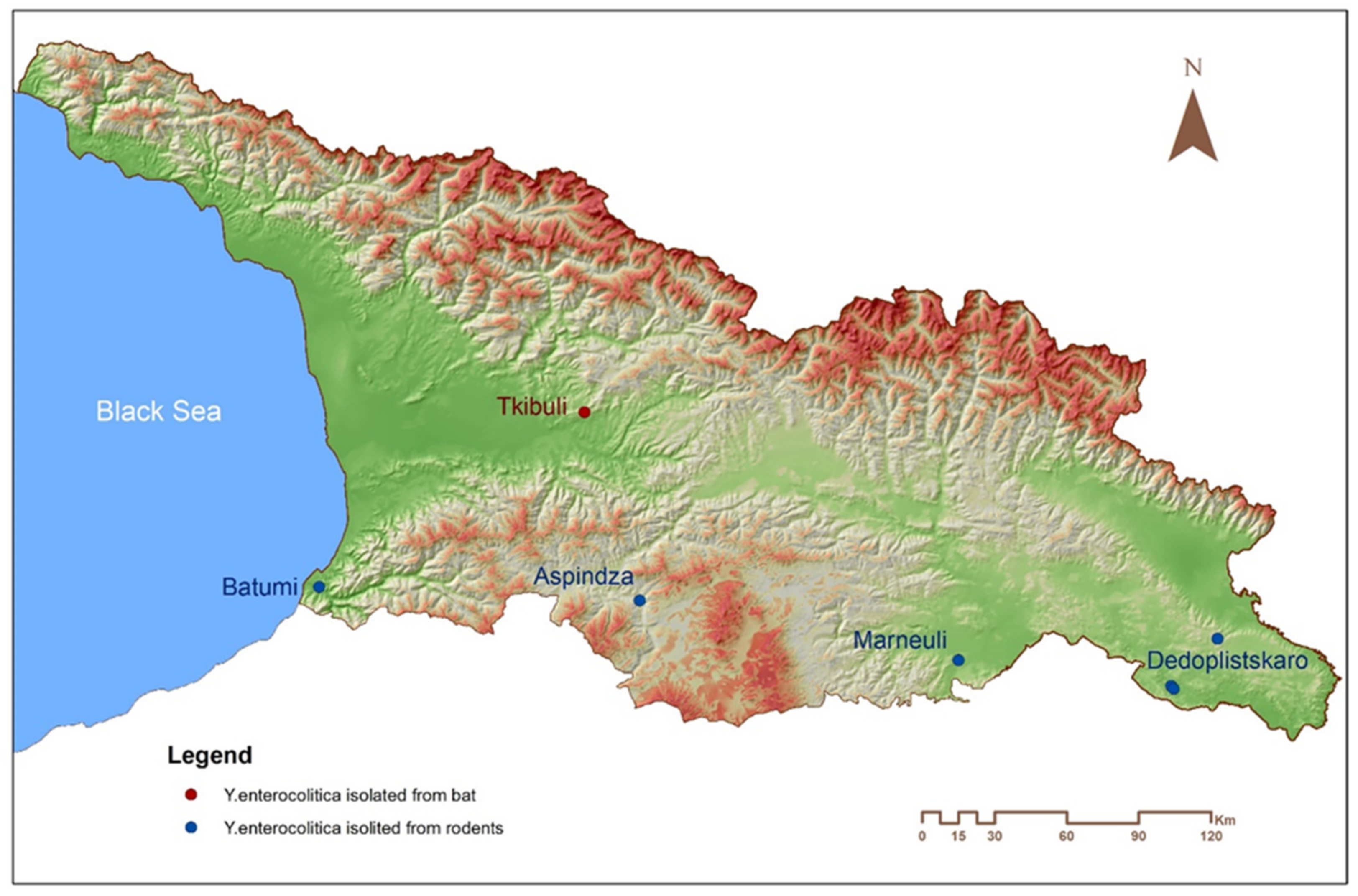
2.2. Field Investigations
2.3. Microbiology
2.4. Determination of Plasmid Content
2.5. Selection of Y. enterocolitica Isolates for Sequencing
2.6. Genome Sequencing and Assembly
2.7. Genome Sequence Processing and Phylogenetic Analyses
3. Results
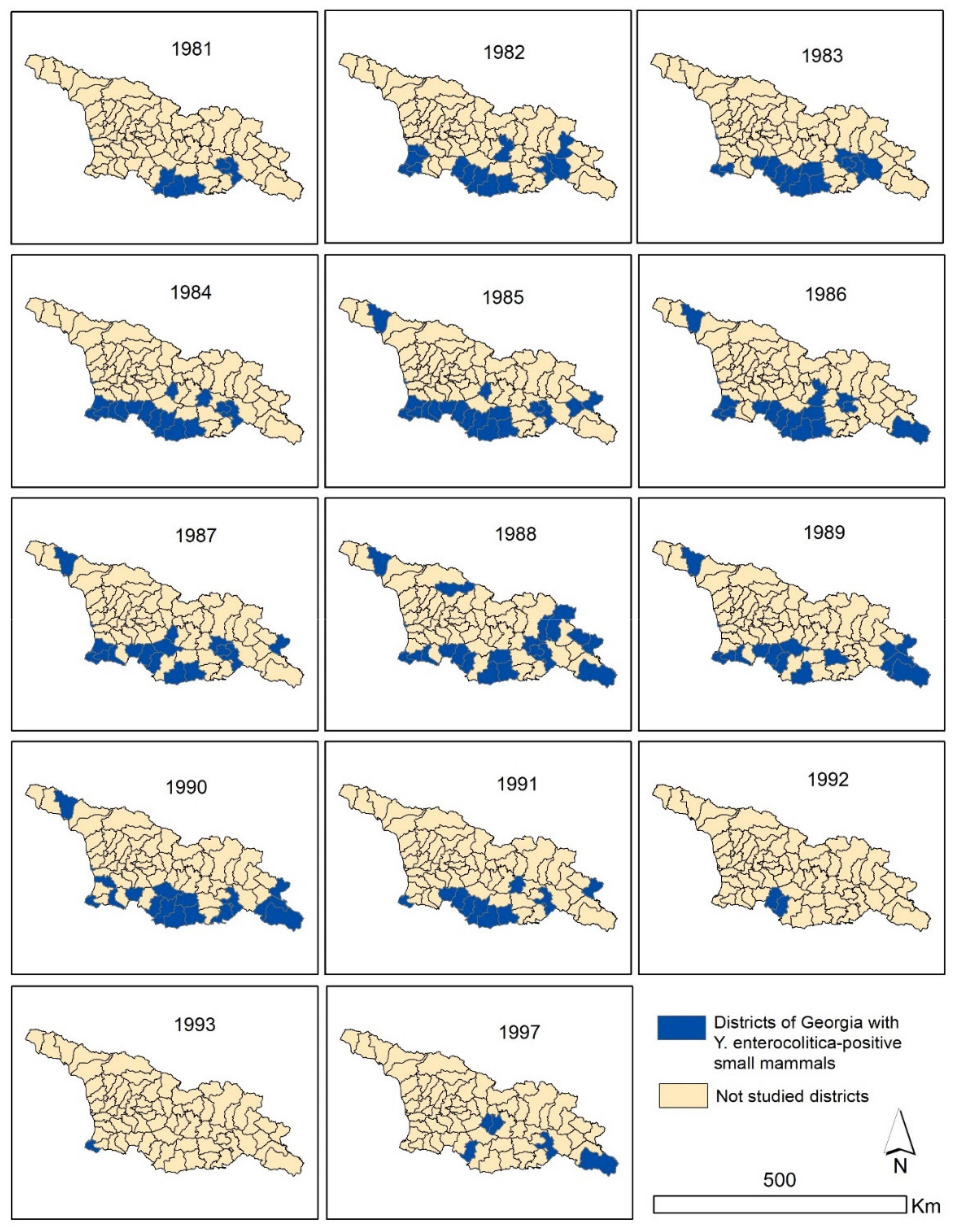
3.1. Prevalence of Y. entercolitica in Rodents and Shrews Based on Archived Records
3.2. Isolation of Y. enterocolitica and Y. enterocolitica-Like Organisms during the Perspective Study
3.3. Phenotypical Characterization of Freshly Isolated Y. enterocolitica Strains
3.4. Genome Size and Sequences of Y. entercolitica Strains Isolated from Small Mammals
3.5. Virulence Genes of Sequenced Y. entercolitica Isolates
3.6. Plasmids
3.7. Phylogenetic Relationship
4. Discussions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sulakvelidze, A. Yersiniae other than Y. enterocolitica, Y. pseudotuberculosis, and Y. pestis: The ignored species. Microbes Infect. 2000, 2, 497–513. [Google Scholar] [CrossRef]
- Wren, B.W. The yersiniae—A model genus to study the rapid evolution of bacterial pathogens. Nat. Rev. Microbiol. 2003, 1, 55–64. [Google Scholar] [CrossRef]
- Bancerz-Kisiel, A.; Pieczywek, M.; Łada, P.; Szweda, W. The most important virulence markers of Yersinia enterocolitica and their role during infection. Genes 2018, 9, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottone, E.J. Yersinia enterocolitica: The charisma continues. Clin. Microbiol. Rev. 1997, 10, 257–276. [Google Scholar] [CrossRef] [PubMed]
- Batzilla, J.; Heesemann, J.; Rakin, A. The pathogenic potential of Yersinia enterocolitica 1A. Int. J. Med. Microbiol. 2011, 301, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Gage, K.L.; Kosoy, M.Y. Natural history of plague: Perspectives from more than a century of research. Annu. Rev. Entomol. 2005, 50, 505–528. [Google Scholar] [CrossRef] [Green Version]
- Wimsatt, J.; Biggins, D.E. A review of plague persistence with special emphasis on fleas. J. Vector. Borne Dis. 2009, 46, 85–99. [Google Scholar]
- Bancerz-Kisiel, A.; Szweda, W. Yersiniosis—A zoonotic foodborne disease of relevance to public health. Ann. Agric. Environ. Med. 2015, 22, 397–402. [Google Scholar] [CrossRef]
- Arrausi-Subiza, M.; Gerrikagoitia, X.; Alvarez, V.; Ibabe, J.C.; Barral, M. Prevalence of Yersinia enterocolitica and Yersinia pseudotuberculosis in wild boars in the Basque Country, northern Spain. Acta Vet. Scand. 2016, 58, 4. [Google Scholar] [CrossRef] [Green Version]
- Muhldorfer, K.; Wibbelt, G.; Haensel, J.; Riehm, J.; Speck, S. Yersinia species isolated from bats, Germany. Emerg. Infect. Dis. 2010, 16, 578–580. [Google Scholar] [CrossRef]
- Nikolova, S.; Tzvetkov, Y.; Najdenski, H.; Vesselinova, A. Isolation of pathogenic yersiniae from wild animals in Bulgaria. J. Vet. Med. Ser. B 2001, 48, 203–209. [Google Scholar] [CrossRef]
- Shayegani, M.; Stone, W.B.; DeForge, I.; Root, T.; Parsons, L.M.; Maupin, P. Yersinia enterocolitica and related species isolated from wildlife in New York State. Appl. Environ. Microbiol. 1986, 52, 420–424. [Google Scholar] [CrossRef] [Green Version]
- Syczyło, K.; Platt-Samoraj, A.; Bancerz-Kisiel, A.; Szczerba-Turek, A.; Pajdak-Czaus, J.; Łabuć, S.; Procajło, Z.; Socha, P.; Chuzhebayeva, G.; Szweda, W. The prevalence of Yersinia enterocolitica in game animals in Poland. PLoS ONE 2018, 13, e0195136. [Google Scholar] [CrossRef]
- Backhans, A.; Fellström, C.; Lambertz, S.T. Occurrence of pathogenic Yersinia enterocolitica and Yersinia pseudotuberculosis in small wild rodents. Epidemiol. Infect. 2011, 139, 1230–1238. [Google Scholar] [CrossRef] [PubMed]
- Aldová, E.; Lím, D. Yersinia enterocolitica in small rodents. Pilot study in two wildlife areas. Zentralbl. Bakteriol. Orig. A 1974, 226, 491–496. [Google Scholar]
- Fukushima, H.; Hao, Q.; Wu, K.; Hu, X.; Chen, J.; Guo, Z.; Dai, H.; Qin, C.; Lu, S.; Gomyoda, M. Yersinia enterocolitica O9 as a possible barrier against Y. pestis in natural plague foci in Ningxia, China. Curr. Microbiol. 2001, 42, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Iinuma, Y.; Hayashidani, H.; Kaneko, K.; Ogawa, M.; Hamasaki, S. Isolation of Yersinia enterocolitica serovar O8 from free-living small rodents in Japan. J. Clin. Microbiol. 1992, 30, 240–242. [Google Scholar] [CrossRef] [Green Version]
- Joutsen, S.; Laukkanen-Ninios, R.; Henttonen, H.; Niemimaa, J.; Voutilainen, L.; Kallio, E.R.; Helle, H.; Korkeala, H.; Fredriksson-Ahomaa, M. Yersinia spp. in wild rodents and shrews in Finland. Vector Borne Zoonotic Dis. 2017, 17, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Kapperud, G. Yersinia enterocolitica in small rodents from Norway, Sweden and Finland. Acta Pathol. Microbiol. Scand. B 1975, 83, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Evstafiev, I.; Tovpinets, N. Small mammals as reservoirs and vectors of yersiniosis pathogens (Yersinia enterocolitica and Y. pseudotuberculosis). Theriol. Ukrain. 2020, 19, 115–121. [Google Scholar] [CrossRef]
- Bakanidze, L.; Imnadze, P.; Chubinidze, S.; Tsertsvadze, N.; Mgeladze, G.; Shalutashvili, I.; Tsanava, S.; Shavishvili, M.; Manvelyan, J.; Ninashvili, N.; et al. Surveillance on Plague in Natural Foci in Georgia. In Emerging and Endemic Pathogens. NATO Science for Peace and Security Series A: Chemistry and Biology; O’Connell, K., Skowronski, E., Sulakvelidze, A., Bakanidze, L., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 21–28. [Google Scholar]
- Shidlovsky, M. Guide to Rodents of Transcaucasia, 2nd ed.; Metsniereba: Tbilisi, Georgia, 1976; p. 255. [Google Scholar]
- Bukhnikashvili, A.; Kandaurov, A. The Annotated List of Mammals of Georgia. In Proceedings of the Institute of Zoology of Academy of Sciences of the Georgia; Metsniereba: Tbilisi, Georgia, 2002; Volume 21, pp. 319–340. [Google Scholar]
- Wilson, D.E.; Reeder, D.M. Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2005; pp. 1–2142. [Google Scholar]
- Kado, C.I.; Liu, S.T. Rapid procedure for detection and isolation of large and small plasmids. J. Bacteriol. 1981, 145, 1365–1373. [Google Scholar] [CrossRef] [Green Version]
- Imnadze, T.; Natradze, I.; Zhgenti, E.; Malania, L.; Abazashvili, N.; Sidamonidze, K.; Khmaladze, E.; Zakalashvili, M.; Imnadze, P.; Arner, R.J.; et al. Identification of a novel Yersinia enterocolitica strain from bats in association with a bat die-off that occurred in Georgia (Caucasus). Microorganisms 2020, 8, 1000. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, A.S.; Gunnison, J.N. The action of Pasteurella pestis bacteriophage on Pasteurella, Salmonella and Shigella. J. Bacteriol. 1947, 54, 70. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Duan, R.; Xia, S.; Hao, Q.; Yang, J.; Xiao, Y.; Qiu, H.; Shi, G.; Wang, S.; Gu, W.; et al. Ecology and geographic distribution of Yersinia enterocolitica among livestock and wildlife in China. Vet. Microbiol. 2015, 178, 125–131. [Google Scholar] [CrossRef]
- Urgeová, E.; Lysý, J. Ecology study of Yersiniae isolated from small terrestrial mammals captured in the territory of the Barrage System Gabcikovo 1990–1994. Contrib. Microbiol. Immunol. 1995, 13, 46–47. [Google Scholar] [PubMed]
- Tennant, S.M.; Grant, T.H.; Robins-Browne, R.M. Pathogenicity of Yersinia enterocolitica biotype 1A. FEMS Immunol. Med. Microbiol. 2003, 38, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Duan, Q.; Xia, P.; Nandre, R.; Zhang, W.; Zhu, G. Review of newly identified functions associated with the heat-labile toxin of enterotoxigenic Escherichia coli. Front. Cell. Infect. Microbiol. 2019, 9, 292. [Google Scholar] [CrossRef] [Green Version]
- Platt-Samoraj, A.; Żmudzki, J.; Pajdak-Czaus, J.; Szczerba-Turek, A.; Bancerz-Kisiel, A.; Procajło, Z.; Łabuć, S.; Szweda, W. The Prevalence of Yersinia enterocolitica and Yersinia pseudotuberculosis in small wild rodents in Poland. Vector Borne Zoonotic Dis. 2020, 20, 586–592. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| ID. | Tissue | Host Species | Location | Year |
|---|---|---|---|---|
| 18ENCDC AsAtc Rint-11-12 | Intestine | Rattus norvegicus | Aspindza | 2018 |
| LcENCDC Rint 5 | Intestine | Rattus norvegicus | Marneuli | 2018 |
| 18E17 065 D I-33 | Intestine | Microtus socialis | Dedoplistskaro | 2018 |
| 18E17 065 D I Rint-95 | Intestine | Apodemus witherbyi | Dedoplistskaro | 2018 |
| 18E17 065 D I Rint-91 | Intestine | Mus macedonicus | Dedoplistskaro | 2018 |
| B18 Rint-231 | Intestine | Shrew (Crocidura sp.) | Batumi | 2018 |
| 1981 | 1982 | 1983 | 1984 | 1985 | 1986 | 1987 | 1988 | 1989 | 1990 | 1991 | 1992 | 1993 | 1997 | Total | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Apodemus spp. Arvicola terrestris | 1 | 8 | 25 4 | 21 1 | 32 3 | 69 | 13 | 2 | 3 3 | 5 | 1 | 180 11 | |||
| Chionomys nivalis | 4 | 4 | |||||||||||||
| Chionomys roberti | 7 | 7 | |||||||||||||
| Cricetulus migratorius | 1 | 1 | 1 | 3 | |||||||||||
| Dryomys nitedula | 1 | 1 | |||||||||||||
| Meriones libycus | 1 | 2 | 8 | 1 | 2 | 1 | 15 | ||||||||
| Mesocricetus brandti | 5 | 1 | 2 | 2 | 2 | 12 | |||||||||
| Microtus arvalis | 8 | 76 | 203 | 165 | 266 | 130 | 116 | 88 | 123 | 73 | 121 | 17 | 1386 | ||
| Microtus majori | 10 | 1 | 2 | 3 | 1 | 5 | 1 | 23 | |||||||
| Microtus socialis | 14 | 3 | 2 | 39 | 7 | 5 | 3 | 73 | |||||||
| Mus musculus | 1 | 1 | 11 | 21 | 11 | 16 | 11 | 18 | 8 | 15 | 7 | 1 | 10 | 131 | |
| Rattus rattus | 4 | 2 | 1 | 1 | 8 | ||||||||||
| Rattus norvegicus | 3 | 6 | 19 | 20 | 19 | 51 | 38 | 10 | 4 | 17 | 11 | 198 | |||
| Soricidae spp. | 1 | 4 | 1 | 6 | 5 | 1 | 5 | 5 | 3 | 1 | 32 | ||||
| Total | 13 | 107 | 245 | 252 | 342 | 243 | 247 | 186 | 147 | 120 | 149 | 17 | 1 | 15 | 2084 |
| Genomic Properties | 18ENCDC AsAtc Rint-11-12 | LcENCDC Rint 5 | 18E17 065 D I-33 | 18E17 065 D I Rint-95 | 18E17 065 D I Rint-91 | B18 Rint-231 |
|---|---|---|---|---|---|---|
| Assembly size (bp) | 4,673,420 | 4,696,602 | 4,899,686 | 4,872,315 | 4,871,034 | 4,775,715 |
| N contigs | 40 | 45 | 83 | 75 | 95 | 124 |
| G+C content (%) | 47.06 | 47.29 | 46.41 | 46.5 | 46.46 | 46.96 |
| Number of protein-coding genes (CDS) | 4218 | 4258 | 4549 | 4455 | 4453 | 4392 |
| CDS (Function assigned) | 2894 | 2916 | 2920 | 2940 | 2938 | 2789 |
| CDS (Hypothetical/putative) | 1324 | 1342 | 1629 | 1515 | 1515 | 1603 |
| Genes | 3103 | 3132 | 3124 | 3144 | 3142 | 3001 |
| Number of tRNAs | 66 | 63 | 69 | 69 | 69 | 72 |
| rRNAs (23S, 16S, and 5S) | 6 | 4 | 9 | 5 | 5 | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imnadze, T.; Malania, L.; Chakvetadze, N.; Burjanadze, I.; Abazashvili, N.; Zhgenti, E.; Sidamonidze, K.; Khmaladze, E.; Martashvili, V.; Tsertsvadze, N.; et al. Evidence of Extensive Circulation of Yersinia enterocolitica in Rodents and Shrews in Natural Habitats from Retrospective and Perspective Studies in South Caucasus. Pathogens 2021, 10, 939. https://doi.org/10.3390/pathogens10080939
Imnadze T, Malania L, Chakvetadze N, Burjanadze I, Abazashvili N, Zhgenti E, Sidamonidze K, Khmaladze E, Martashvili V, Tsertsvadze N, et al. Evidence of Extensive Circulation of Yersinia enterocolitica in Rodents and Shrews in Natural Habitats from Retrospective and Perspective Studies in South Caucasus. Pathogens. 2021; 10(8):939. https://doi.org/10.3390/pathogens10080939
Chicago/Turabian StyleImnadze, Tata, Lile Malania, Neli Chakvetadze, Irma Burjanadze, Natalia Abazashvili, Ekaterine Zhgenti, Ketevan Sidamonidze, Ekaterine Khmaladze, Vakhtang Martashvili, Nikoloz Tsertsvadze, and et al. 2021. "Evidence of Extensive Circulation of Yersinia enterocolitica in Rodents and Shrews in Natural Habitats from Retrospective and Perspective Studies in South Caucasus" Pathogens 10, no. 8: 939. https://doi.org/10.3390/pathogens10080939