
Helicobacter pylori-Induced Inflammation: Possible Factors Modulating the Risk of Gastric Cancer


1
Department of Zoology, Deen Dayal Upadhyaya, Gorakhpur University, Gorakhpur 273009, India
2
Health Sciences Center, Department of Cell Biology and Biochemistry, Texas Tech University, 3601 4th Street, Lubbock, TX 79430, USA
3
Department of Gastroenterology, Sanjay Gandhi Postgraduate Institute of Medical Sciences, Lucknow 226014, India
*
Author to whom correspondence should be addressed.
Pathogens 2021, 10(9), 1099; https://doi.org/10.3390/pathogens10091099
Submission received: 13 July 2021
/
Revised: 21 August 2021
/
Accepted: 25 August 2021
/
Published: 29 August 2021
(This article belongs to the Special Issue Inflammation and Gastrointestinal/Hepatobiliary Cancers)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Chronic inflammation and long-term tissue injury are related to many malignancies, including gastric cancer (GC). Helicobacter pylori (H. pylori), classified as a class I carcinogen, induces chronic superficial gastritis followed by gastric carcinogenesis. Despite a high prevalence of H. pylori infection, only about 1–3% of people infected with this bacterium develop GC worldwide. Furthermore, the development of chronic gastritis in some, but not all, H. pylori-infected subjects remains unexplained. These conflicting findings indicate that clinical outcomes of aggressive inflammation (atrophic gastritis) to gastric carcinogenesis are influenced by several other factors (in addition to H. pylori infection), such as gut microbiota, co-existence of intestinal helminths, dietary habits, and host genetic factors. This review has five goals: (1) to assess our current understanding of the process of H. pylori-triggered inflammation and gastric precursor lesions; (2) to present a hypothesis on risk modulation by the gut microbiota and infestation with intestinal helminths; (3) to identify the dietary behavior of the people at risk of GC; (4) to check the inflammation-related genetic polymorphisms and role of exosomes together with other factors as initiators of precancerous lesions and gastric carcinoma; and (5) finally, to conclude and suggest a new direction for future research.
1. Introduction
Gastric cancer (GC) is the most common cancer contributing to 5.5% of all new cases of cancers. Moreover, it is the fourth most lethal cancer, resulting in 7.7% of all deaths worldwide [1]. It has a poor prognosis, with a survival rate of less than 5 years among 80% of cases [2]. Although the incidence of GC has decreased, it remains a significant global health burden, with the highest burden in Asia [1]. Helicobacter pylori (H. pylori) infects over half of the world’s population, and this bacterium is the main cause of non-cardia GC [3,4,5]. However, geographical variations of bacterial virulence, age of acquisition of infection, host genetics, and environmental factors may lead to variation in the incidence of GC [6]. Paleo Correa’s hypothesis suggests that gastric carcinogenesis progresses in multiple stages and is caused by various factors. The histological cascade associated with GC is well characterized. It proceeds from normal mucosa infected with H. pylori to chronic active gastritis, atrophic gastritis (AG), intestinal metaplasia (complete at first, then incomplete later), dysplasia, hyperplasia, and adenocarcinoma [7,8,9,10].
A chronic inflammatory state is known to be critical for H. pylori-induced GC; the molecular mechanisms underlying how H. pylori communicate with gastric epithelial cells directly or indirectly to trigger gastric carcinogenesis remain unknown. GC exhibits a multi-factorial etiology. Moreover, owing to some paradoxical observations, it is now speculated that in addition to H. pylori infection, diet and host genetic factors are responsible for the progress from H. pylori-induced inflammation to GC [9,11,12]. A few recent studies also proposed that the increasing prevalence of autoimmune gastritis and dysbiosis of gut microbiota and increased use of antibiotics and acid suppressants, might have led to the variation in the risk of GC, primary gastric lymphoma (PGL), and neuroendocrine carcinoma [13,14,15]. Inter-individual variations on disease susceptibility of GC can also be influenced by other infections such as Epstein-Barr virus, intestinal helminths, and host genetic differences in cytokine genes [9,16,17]. Therefore, this review may give a new insight into geographical and inter-individual variation in inflammation-related precancerous lesions and mechanisms of how H. pylori infection might alter the disease. This may be helpful for preventive strategies in subjects with high risk for the development of GC, a potentially fatal malignancy, and may further givea new, more stringent concept for clinical research.
2. Mechanism of Chronic Inflammation and Multi-Step Sequel of Gastric Carcinogenesis
GC is an inflammation-associated carcinoma promoted by H. pylori infection, characterized by ongoing chronic gastritis, formation of metaplastic epithelia, and finally genetic instability in the gastric mucosal epithelium [18,19]. The relationship between chronic inflammation and cancer dates back to Virchow, who, in 1863, hypothesized that the origin of cancer was at the sites of chronic inflammation [20]. Many shreds of evidence proved that the inflammation might result from persistent mucosal or epithelial cell colonization by H. pylori, which may cause GC [11,21,22]. Persistent inflammation leads to increased cellular turnover, especially in the epithelium, and provides selection pressure, which may result in the emergence of cells at high risk for malignant transformation [23]. The association of inflammatory signals with intracellular pathways in gastric epithelial cells eventually leads to uncontrolled cell division, and differentiation remains inadequate. Whereas acute injury and inflammation associated with healing are usually self-limited, chronic injuries or inflammation over decade’s leads to a sustained expansion of proliferative tissue zones that are predisposed to neoplastic progression [24].
The precancerous cascade advances slowly and steadily, and it may take years or several decades to develop malignancy since the initiation of the cascade. However, the rate of progression from metaplasia to dysplasia is not the same in all individuals. A study revealed that the progression rate was two times higher in subjects more than 40 years of age than their younger counterparts [25]. In a subset of patients, this inflammatory process leads to loss of parietal cells and development of AG, followed by intestinal metaplasia (IM), dysplasia, and cancer [26]. Surprisingly, the majority of patients infected with H. pylori not only incline towards the development of pre-malignant lesions or GC, but also towards other gastroduodenal diseases (Figure 1). It is also speculated that all the stages before the development of dysplasia are reversible, although this is still somewhat controversial [24,25,26].
3. H. pylori-Triggered Inflammation and Gastric Precursor Lesions
3.1. The Link between H. pylori-Induced Inflammation and Gastric Carcinogenesis
The persistent inflammation caused by H. pylori recruits immune cells through steady generation of cytokines, resulting in histological changes, including pre-neoplastic gastric lesions (AG and IM), as well as the production of reactive radicals that might disrupt the host’s DNA, favoring the onset of carcinoma [27]. The International Agency for Research on Cancer (a branch of the World Health Organization) has previously approved the link between H. pylori and GC [28]. In the previous three decades, H. pylori has been implicated in the development of gastroduodenal diseases, including GC. Several case-control studies retrospectively examined the association between H. pylori and GC and revealed up to six-fold increased risk of GC in the presence of H. pylori infection [4,29,30,31]. The virulence of the infecting strain is a key predictor determining who would develop disease in people infected with H. pylori. Cytotoxin-associated antigen A is a marker for the Cag pathogenicity island, which contains genes required for pathogenic strains to cause increased inflammation [32,33].
Gastric inflammation not only results from H. pylori infection; dietary and environmental factors also influence the development of gastritis [34]. H. pylori (or its products) may produce gastric inflammation by two main mechanisms. Firstly, the organism may interact with epithelial cells on the surface, causing direct cell injury or the release of epithelial-derived pro-inflammatory mediators (chemokines). Secondly, H. pylori-derived products may obtain access to the mucosa under the surface, triggering non-specific and specific immune responses in the host, including the production of cytokine messengers. Thus, H. pylori infection induces both humoral and cellular immune responses.
H. pylori-induced inflammation, which includes neutrophil and macrophage activation and induction of Th1 response as well as alterations in host physiological responses associated with infection, contributes to mucosal damage [35]. Since H. pylori induces a strong humoral immune response, the Th2-cell response is expected. Paradoxically, H. pylori-specific stomach mucosal T-lymphocytes are mostly Th1 in nature [36]. This Th1 phenotype may be related to increased antral IL-18 production, and when combined with Fas-mediated apoptosis of H. pylori-specific T-cell clones, may favor H. pylori persistence [37,38]. Studies in gene-targeted mice have further shown that Th1 cytokines cause gastritis, but Th2 cytokines protect against inflammation [39]. The H. pylori-secreted peptidyl-prolyl cis, trans-isomerase, also drives the Th17 response, which promotes pro-inflammatory, low cytotoxic, gastric, tumor-infiltrating lymphocytes, matrix breakdown, and pro-angiogenic pathways, ultimately leading to the development of GC [40]. However, the enigma of why only a subset of H. pylori-infected individuals develops GC still continues.
3.2. The Link between H. pylori-Triggered AG, IM, and GC
Most individuals infected with H. pylori have infiltrating polymorphonuclear leucocytes in the antrum and the fundus of the stomach in the acute stage and lymphocytic inflammatory response later [41]. It is very slow to disappear after eradicating H. pylori [42]. In the treatment of peptic ulcers, early precancerous lesions of GC, and PGL, eradication of H. pylori infection is recommended [43,44]. In some cases, chronic gastritis is associated with gastric ulcers, which may increase the risk of GC, though duodenal ulcers caused by H. pylori are negatively associated with GC development [4,45,46].
Atrophic gastritis (AG), which is defined as the loss of specific glandular tissues, is a well-known early morphologic alteration that occurs before the onset of GC. However, the presence of AG is associated with a 0.5% to 1% annual risk of GC advancement [47,48]. Two types of AG exist in stomach mucosa: the first one is H. pylori-associated multifocal AG and the second form is corpus AG with anti-parietal cell and anti-intrinsic factor antibodies, which are associated with pernicious anemia. The process of AG occurs in approximately half of the H. pylori-colonized population, and it develops initially in those people and in those parts of the stomach where inflammation is the most severe [49,50]. In particular, H. pylori Cag+-pathogenicity-island-containing strains induce inflammation and have a higher risk of development of AG and GC [51,52,53,54]. Areas of gland loss and IM extend with time multi-focally, and although these do not give rise to any specific symptoms, these increase the risk for GC by 5- to 90-fold depending on the extent and severity of AG [55]. Paradoxically, patients with advanced AG and acholiorhydria revealed that even after eradication of H. pylori, there may be considerable risk of GC [56].
The link between H. pylori-triggered inflammation and IM is well established and is considered a major determinant of gastric carcinogenesis, but the bacterium is frequently undetected in these lesions [57,58]. Pre-neoplastic metaplasia has also evolved in H. pylori-infected mice models, notably spasmolytic polypeptide-expressing metaplasia (SPEM) [59]. IM occurs when the mucosa of the stomach mimics intestinal epithelium with goblet cells [60]. IM is classified into three types by Jass and Filipe, particularly type I (complete form), type II (incomplete form), and type III (intermediate form) [61]. The presence of type II or III IM is found to be related to a 20-fold greater risk of GC [62]. Some studies also proved the link between H. pylori-induced gastritis and IM [63,64]. Three mechanisms could generate this risk: (i) the metaplastic tissue is an early step in a multi-step induction process of gastric carcinogenesis; (ii) IM exhibits epigenetic changes, raises the pH of gastric juice by replacing oxyntic mucosa, and favors the growth of H. pylori capable of producing endogenous mutagens; (iii) IM is only a marker for chronic gastritis due to H. pylori infection or pernicious anemia [55,60]. Individuals with a high risk for GC can be identified using IM.
4. Risk Modulation of GC by Co-Existence of Gut Microbiota and Infestation with Intestinal Helminths
4.1. Gut Microbiota Other Than H. pylori and Their Role in Cascade of Gastric Cancer
Metagenomics and advanced nucleotide sequencing techniques unhooked the mucosal and luminal composition of the gut microbiota and revealed their significance in natural habitats other than H. pylori [65,66,67,68]. In transgenic mice, gastrointestinal over-expression of human IL-1β was sufficient for the development of gastric dysplasia and carcinoma in a stepwise manner [69,70]. Some experimental animal models, particularly transgenic insulin-gastrin (INS-GAS) mice model, showed the importance of non-H. pylori gastric microbiome in enhancing the effects of H. pylori in the development of GC [71,72]. Patients with GC also showed dysbiotic microbial population with genotoxic potential that differs from the patients with chronic gastritis [73]. The H. pylori infection generates an inflammatory reaction in the stomach, resulting in the loss of parietal cells and an elevation in gastric pH. H. pylori may contribute to microbial dysbiosis, and effective eradication can restore the gut microbiota to a state comparable to that of uninfected people [74]. The colonization of gut microbiota grows over time as AG develops, and in spite of H. pylori becoming diminished, the precancerous lesionscontinue to develop (Figure 2). Higher metabolites produced by dysbiosis of the gut microbiome, such as N-nitroso compounds and lactate, are thought to affect the immunological response as well as DNA damage, resulting in gastric carcinogenesis [75,76]. Recent studies proved that gut microbiota plays a significant role in the progression of gastric inflammation, AG, and IM after H. pylori eradication [77,78]. However, retrospective and only association-based findings are the limitations of these studies. It is unclear that the microbial changes seen in GC cause disease or are a result of the histologic progression through the precancerous cascade. Therefore, better understanding of the role of the gut microbiota in the development and progression of GC should lead to better diagnostic and preventive options.
4.2. Intestinal Helminth Infection Down-Regulates the Effect of H. pylori in Gastric Carcinogenesis
Co-infection with intestinal helminths may affect the outcome of H. pylori infection. A study on two Colombian communities infected with virulence-associated genotypes of H. pylori found a high versus low incidence of GC (endemic area of helminth infection), supporting the notion that intestinal helminth infection reduces the influence of H. pylori on gastric carcinogenesis [79]. Concurrent helminth infection has been shown in animal experiments to reduce the severity of H. pylori-induced gastritis [80]. Recently, a study on gastric mucosal samples also showed decreased expression of proinflammatory cytokines and predominant Th2 response (higher level IL-4) among H. pylori-infected humans co-infected with intestinal helminthes [81]. The higher load of H. pylori and intestinal parasites in the India and Venezuela populations is associated with a low risk of GC, which might explain these enigmas [9,81,82]. One study in a Chinese population found that concurrent helminth infections altered serological IgG responses as well as the pepsinogen I/II ratio, indicating a lower chance of developing H. pylori-induced atrophy [83]. A study on children from regions of low versus high risk of GC but similar H. pylori seropositivity showed that subjects from the low-risk area were more commonly infected with helminths and showed higher Th2-associated IgG1 responses to H. pylori infection [84]. These findings suggest that early childhood exposure to intestinal helminths induces immunoregulatory lymphocytes and anti-inflammatory cytokines such as IL-4, IL-10, and TGF-β and lowers the expression of pro-inflammatory IFN-γ, TNF-α, and IL-1β, including Th1-associated IP-10, RANTES, and MIP-1β. Cysteinyl leukotrienes are produced by tuft cells during helminth infection [85]. On the other hand, chemosensing by tuft cells activates group 2 innate lymphoid cells (ILC2), leading to an increase in tuft cell frequency, and exhibiting significant physiologic alterations in the tissue, including hyperplasia of mucus-secreting goblet cells [86]. Intestinal remodeling and helminth removal require this feedback control pathway. This mechanism provides a new direction for future research towards co-infection of helminths and H. pylori. Another study showed that H. pylori infection causes diseases by hypermethylation of key cellular promoters at CpG dinucleotides (promoter silencing), especially in gastric mucosa [87]. However, helminths may induce pathological changes by epigenetic reprogramming of host cells [88]. Poor hygienic and environmental conditions favor the endemic prerequisite for transmission and existence of both H. pylori and intestinal helminths acquired at an early age. In particular, helminths promote a Th2-polarizing response that may decrease H. pylori-induced cancer risk in individuals later in life (Figure 3). Studies addressing this issue are warranted and represent an intriguing area of research with the potential for future studies focused on H. pylori pathogenesis.
5. Relationship between Dietary Behavior of the People and Risk of GC
Diet may interact with bacterial and host genetic factors; dietary variations in different populations may explain difference in prevalence of GC in different areas of the world [89].Though gastric inflammation is caused primarily by H. pylori infection; exposure to other factors such as high salt, preserved food, and bile salts also influences the degree of gastritis [90,91,92,93]. Red and processed meat increase the oxidative stress to the inflamed stomach of individuals infected with H. pylori, putting them at higher risk of GC [94]. Many studies proved that smoking and eating a high-salt diet are both substantial risk factors for GC [95,96,97]. Obesity and alcohol (some say it is controversial depending on the amount) are also well-known risk factors for GC [98,99]. A diet rich in fresh fruits and vegetables, especially lycopene and lycopene-containing foods, and possibly vitamin C and selenium, can lower the risk of GC [100]. Many epidemiological studies found that diets high in antioxidant-rich fresh fruits and vegetables can minimize oxidative stress and lower the incidence of GC [101,102]. Turmeric has also been found to inhibit the growth of H. pylori and its anti-carcinogenic compound curcumin showed increased bioavailability in the presence of piperine (a compound from black pepper), and quercetine (a compound found in onion), which may decrease chronic inflammation [9,103]. In India, the eastern and southern parts of the country have a higher GC frequency than northern regions. Rice, non-vegetarian foods, mainly fish, are prevalent in the eastern Indian diet, which is spicy and with more salt [104]. In contrast, the northern Indian diet is wheat-based, and a greater proportion of people are vegetarian [104]. This observation suggests that geographical variations of dietary behavior are a significant component in the development and prevention of GC.
6. Inflammation-Related Genetic Polymorphisms Together with Other Factors as Initiators of Precancerous Lesions and GC
6.1. Cytokine Genes Influence Individual Response to Carcinogenic Exposures
Chronic inflammation can occur in genetically susceptible hosts with defective mucosal host defense systems or dysregulated immune responses, leading to excessively aggressive responses to ubiquitous antigens, which is the root cause of inflammation-related carcinogenesis [20,23,105]. Inflammation-related genetic polymorphisms act as initiators of H. pylori-induced chronic atrophic gastritis, and together with other factors they become the precursor of precancerous lesions and carcinoma [106,107]. Cytokines induced by specific stimuli, such as toxins produced by pathogens, are involved in immunity, inflammation, and cell proliferation. By secreting cytokines and recruiting specific inflammatory cells, H. pylori influence both the mucosal and systemic immune responses. In addition, H. pylori cause cellular changes as well as changes in genes that are important for epigenetic integrity and mucosal homeostasis. These genetic changes during the development of chronic inflammation are the subject of extensive investigation. A primary strategy for screening and early detection of GC in individuals at risk may include finding persons who have H. pylori infection with a pro-inflammatory makeup [108]. Genetic variations in pro-inflammatory and anti-inflammatory cytokine genes influence individual response to carcinogenic exposures. The degree of inflammation in the host tissue is determined not only by external factors such as infection with bacteria but also on the host’s genetic makeup, whether they have a high- or a low-producing genotype.
Pro- and anti-inflammatory cytokines modulate the inflammatory response of the stomach mucosa [109,110]. Pro-inflammatory cytokines activate inflammatory cells by the migration of neutrophils, mononuclear phagocytes, eosinophils, and mast cells (e.g., IL-8, MCP-1, RANTES, TNF-α,and IL-1), and also play a significant role in acquired immune responses that can regulate the growth, differentiation, and activation of lymphocytes, mast cells, eosinophils, and other hemopoietic cells (e.g., IFN- γ, IL-12 etc.) [111]. In another way, anti-inflammatory cytokine (e.g., IL-10, a product of Th2 cells) is a potent factor for suppressing the inflammatory and neoplastic environment. It inhibits IFN- γ production and antigen-specific T-cell activation by down-regulating antigen presentation as well as IL-1, TNF- α, and IL-6 production by monocytes or macrophages [112]. Cytokines, particularly SNPs of IL-1B, IL-4, IL-6, IL-8, IL-10, IL17A, and IL-17F, may influence cancer prognosis and prevention [113]. A balance between pro- (IL-8) and anti-inflammatory (IL-10) cytokines may influence the degree of chronic inflammation, which is a potential factor in development of gastritis and GC [16,114]. Furthermore, genetic diversity of cytokine genes revealed differences in the severity of H. pylori-induced inflammation and the risk of GC among various populations. More data will be necessary to assess the above hypothesis.
6.2. Inflammatory Gene Polymorphism Alters Acid Production
Acid production, oxidative stress, and DNA damage are all affected by polymorphisms in both bacterial and host genes, which are thought to be a major factor in the pathogenesis of GC [115]. When H. pylori infect the gastric mucosa, it stimulates neutrophils and mononuclear cells, causing them to release a variety of inflammatory cytokines [116]. In particular, IL-1β and TNF-α are potent inhibitors of gastric acid secretion [109,117,118].They promote the development of non-H. pylori gut microbiota capable of sustaining inflammation and continually producing oxidative stress, thereby hastening the process of gastric carcinogenesis. Moreover, long-term H. pylori infection in the context of vulnerable host gene polymorphisms may result in hypochlorhydria or hyperchlorhydria, depending on the degree and location of gastritis, which may proceed to gastric ulceration or GC. However, we believe the integration of all the information will be helpful for future study into novel molecular pathways and processes implicated in H. pylori-induced inflammation.
7. Role of Exosomes in Gastric Cancer
Exosomes are nanovesicles (30–150 nm) of endocytic origin and are largely recognized in the intercellular communications via transfer of vital contents, which may induce tumor cell proliferation, metastasis, inflammation, immune suppression, remodeling of tumor microenvironment, drug resistance, etc [119,120]. Chronic inflammation is one of the most common features associated with peptic ulceration and GC development and progression. Cells of tumor microenvironment are known to play a major role in chronic inflammation. It is reported that H. pylori-infected, GC-cells-derived exosomes were enriched with the phosphorylated (active) form of mesenchymal epithelial transition factor (MET) (Figure 4).Exosome-mediated transfer of MET modulates the macrophages towards a pro-tumorigenesis phenotype, stimulating the secretion of pro-inflammatory cytokine IL-1β that promotes GC cell growth and proliferation [121]. In another important finding, CagA, encoded by cytotoxin-associated gene A, a major virulence factor of H. pylori, was shown to be present in the exosomes isolated from the blood of H. pylori-infected patients [122]. These CagA positive exosomes modulate the gastric epithelial cell behavior. Circulating CagA positive exosomes secreted from gastric epithelial cells may be involved in disease spread and may help in the development of extra-gastric disorders associated with CagA-positive H. pylori infection [122]. SPEM is an important risk factor driven by macrophage-mediated chronic inflammation. It is demonstrated that exosomes derived from the macrophage induced by deoxycholic acid (a type of harmful secondary bile acid) increased the expression of SPEM markers (TFF2 and GSII lectin) in gastric organoids, indicating the role of macrophage-derived exosomes in the onsets of GC [123]. Zheng et al. (2017) showed that tumor-associated macrophage (TAM)-derived exosome enriched with miRNA-21 down-regulates PTEN, suppresses cell apoptosis, and enhances the activation of PI3K/AKT signaling pathway [124]. In another study, TAM-derived exosomes were found to be enriched with apolipoprotein E (ApoE) and were shown to activate PI3K-Akt signaling pathway in recipient GC cells and affect cytoskeleton remodeling and promote GC cells migration [124]. Similarly, GC cells-derived exosomes were shown to activate PI3K/Akt and mitogen-activated protein kinase/extracellular-regulated protein kinase pathways leading to tumor cell proliferation [125]. In another study, exosome derived from GC cells was shown to activate macrophages by inducing NF-κB activity and thus enhanced the release of pro-inflammatory cytokines, including IL-6 and TNF-α, which in turn promotes proliferation and migration of GC cells [126]. The expression of ZFAS1, a long noncoding RNA (lncRNA), is known to be elevated in the GC cells, tumor tissues, serum, and serum exosomes of GC patients. Mechanistically, ZFAS1 promotes proliferation and migration and suppresses apoptosis in GC cells [127]. Wei et al. (2020) reported that miR-15b-3p transferred via exosomes promotes GC development and GES-1 cell malignant transformation by inhibiting the expression of DYNLT1, Cleaved Caspase-9, and Caspase-3 [128].
Cancer-associated fibroblasts (CAFs) play a multifaceted role in cancer progression. It is shown that exosomes derived from the GC cells promote differentiation of mesenchymal stem cells (MSCs) into CAFs. The PKM2-enriched exosomes derived from the GC cells are associated with nuclear factor kappa B (NF-κB) activity and elevated levels of IL-6, IL-8, and other inflammatory cytokines [129]. CAFs are also known to play an important role in drug resistance. It is shown that CAF-derived exosomes transfer miR-522 to GC cells, suppressing the expression of ALOX15 and thus decreasing the lipid-ROS accumulation in GC cells, leading to resistance to paclitaxel and cisplatinm [130]. In a study by Zhang et al. (2018), GC-cell-derived exosomes, by transferring high mobility group box-1 (HMGB1), are shown to activate NF-κB signaling pathway. HMGB1 were shown to induce autophagy and activation of neutrophils via HMGB1/TLR4/NF-κB signaling axis and help in the progression of GC [131]. Altogether, these findings affirm the role of exosomes in chronic inflammation and growth and development of GC.
8. Conclusions and Future Directions
H. pylori: though it is known to be a carcinogen, other factors also influence its carcinogenic potential. The variation in the other factors may partly explain differences in the prevalence of GC in different populations in the world despite high frequency of H. pylori infection in some countries. These factors include the genetic factors of the host, diet, gut microbiota, and other infective agents such as helminths. The knowledge of the modifiable factors in the development of GC such as diet and gut microbiota, which are known to be modified with dietary change, may help in prevention of gastric carcinogenesis. It is critical to learn more about the pathogenesis of H. pylori-induced inflammation, as well as potential variables that influence the risk of GC, not just to find more effective therapies for this frequent malignancy, but also because it might serve as a model for how persistent inflammation plays a role in the development of other gastrointestinal malignancies. Moreover, knowledge about multifactorial pathogenesis of H. pylori-induced gastric carcinogenesis may help to predict the risk of development of GC in an individual with H. pylori infection by complex mathematical modeling; this may help in deciding on surveillance strategy, H. pylori eradication, testing for re-infection, and screening endoscopy, as mass eradication of H. pylori in an unselected population, even in high-risk areas, was not found to be rewarding.
Author Contributions
Conceptualization, S.K. and U.C.G.; writing—original draft preparation, S.K., G.K.P. and U.C.G.; writing—review and editing S.K., G.K.P. and U.C.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work did not receive any funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not Applicable.
Conflicts of Interest
None of the authors have any conflict of interest to declare in relation to this paper.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Berardi, R.; Scartozzi, M.; Romagnoli, E.; Antognoli, S.; Cascinu, S. Gastric cancer treatment: A systematic review. Oncol. Rep. 2004, 11, 911–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plummer, M.; Franceschi, S.; Vignat, J. Global burden of gastric cancer attributable to Helicobacter pylori. Int. J. Cancer 2015, 136, 487–490. [Google Scholar] [CrossRef]
- Uemura, N.; Okamoto, S.; Yamamoto, S.; Matsumura, N.; Yamaguchi, S.; Yamakido, M.; Taniyama, K.; Sasaki, N.; Schlemper, R.J. Helicobacter pylori Infection and the Development of Gastric Cancer. N. Engl. J. Med. 2001, 345, 784–789. [Google Scholar] [CrossRef]
- Hooi, J.K.; Lai, W.Y.; Ng, W.K.; Suen, M.M.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.; Wu, J.C.; et al. Global Prevalence of Helicobacter pylori Infection: Systematic Review and Meta-Analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidd, M.; Lastovica, A.J.; Atherton, J.C. Heterogeneity in the Helicobacter pylori vacA and cagA genes: Association with gastroduodenal disease in South Africa? Gut 1999, 45, 499–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, P. Human gastric carcinogenesis: A multistep and multifactorial process—First American Cancer Society Award Lecture on Cancer Epidemiology and Prevention. Cancer Res. 1992, 52, 6735–6740. [Google Scholar] [PubMed]
- Sugano, K. Premalignant conditions of gastric cancer. J. Gastroenterol. Hepatol. 2013, 28, 906–911. [Google Scholar] [CrossRef]
- Ghoshal, U.C.; Chaturvedi, R.; Correa, P. The enigma of Helicobacter pylori infection and gastric cancer. Indian J. Gastroenterol. 2010, 29, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Correa, P.; Piazuelo, B.M.; Wilson, K. Pathology of Gastric Intestinal Metaplasia: Clinical Implications. Am. J. Gastroenterol. 2010, 105, 493–498. [Google Scholar] [CrossRef] [Green Version]
- Wroblewski, L.E.; Peek, R.M.; Wilson, K. Helicobacter pylori and Gastric Cancer: Factors That Modulate Disease Risk. Clin. Microbiol. Rev. 2010, 23, 713–739. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Ghoshal, U.C. Causal role of Helicobacter pylori infection in gastric cancer: An Asian enigma. World J. Gastroenterol. 2006, 12, 1346–1351. [Google Scholar] [CrossRef]
- Arnold, M.; Park, J.Y.; Camargo, M.C.; Lunet, N.; Forman, D.; Soerjomataram, I. Is gastric cancer becoming a rare disease? A global assessment of predicted incidence trends to 2035. Gut 2020, 69, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Heer, E.V.; Harper, A.S.; Sung, H.; Jemal, A.; Fidler-Benaoudia, M.M. Emerging cancer incidence trends in Canada: The growing burden of young adult cancers. Cancer 2020, 126, 4553–4562. [Google Scholar] [CrossRef]
- Terao, S.; Suzuki, S.; Yaita, H.; Kurahara, K.; Shunto, J.; Furuta, T.; Maruyama, Y.; Ito, M.; Kamada, T.; Aoki, R.; et al. Multicenter study of autoimmune gastritis in Japan: Clinical and endoscopic characteristics. Dig. Endosc. 2020, 32, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kumari, N.; Mittal, R.D.; Mohindra, S.; Ghoshal, U.C. Association between pro-(IL-8) and anti-inflammatory (IL-10) cytokine variants and their serum levels and H. pylori-related gastric carcinogenesis in northern India. Meta Gene 2015, 6, 9–16. [Google Scholar] [CrossRef]
- Shukla, S.K.; Prasad, K.N.; Tripathi, A.; Singh, A.; Saxena, A.; Ghoshal, U.C.; Krishnani, N.; Husain, N. Epstein-Barr virus DNA load and its association with Helicobacter pylori infection in gastroduodenal diseases. Braz. J. Infect. Dis. 2012, 15, 583–590. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela, M.A.; Canales, J.; Corvalan, A.; Quest, A.F.G. Helicobacter pylori-induced inflammation and epigenetic changes during gastric carcinogenesis. World J. Gastroenterol. 2015, 21, 12742–12756. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Meng, W.; Wang, B.; Qiao, L. Helicobacter pylori-induced gastric inflammation and gastric cancer. Cancer Lett. 2014, 345, 196–202. [Google Scholar] [CrossRef]
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Correa, P. Gastric cancer: Overview. Gastroenterol. Clin. N. Am. 2013, 42, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Mishra, K.K.; Srivastava, S.; Aayyagari, A.; Ghosh, K. Development of an animal model of Helicobacter pylori (Indian strain) infection. Indian J. Gastroenterol. 2019, 38, 167–172. [Google Scholar] [CrossRef]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef]
- Correa, P.; Schneider, B.G. Etiology of gastric cancer: What is new? Cancer Epidemiol. Biomark. Prev. 2005, 14, 1865–1868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, P.; Haenszel, W.; Cuello, C.; Zavala, D.; Fontham, E.; Zarama, G.; Tannenbaum, S.; Collazos, T.; Ruiz, B. Gastric precancerous process in a high risk population: Cohort follow-up. Cancer Res. 1990, 50, 4737–4740. [Google Scholar]
- Correa, P.; Piazuelo, M. Natural history of Helicobacter pylori infection. Dig. Liver Dis. 2008, 40, 490–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz, P.; Valderrama, M.V.; Bravo, J.; Quest, A.F.G. Helicobacter pylori and Gastric Cancer: Adaptive Cellular Mechanisms Involved in Disease Progression. Front. Microbiol. 2018, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. International Agency for Research on Cancer: Schistosomes LFaHp. IARC Monogr. Eval. Carcinog. Risks Hum. 1994, 61, 9–243. [Google Scholar]
- Nomura, A.; Stemmermann, G.N.; Chyou, P.-H.; Perez, G.P.; Blaser, M.J. Helicobacter pylori Infection and the Risk for Duodenal and Gastric Ulceration. Ann. Intern. Med. 1994, 120, 977–981. [Google Scholar] [CrossRef] [PubMed]
- Houghton, J.; Wang, T.C. Helicobacter pylori and Gastric Cancer: A New Paradigm for Inflammation-Associated Epithelial Cancers. Gastroenterology 2005, 128, 1567–1578. [Google Scholar] [CrossRef]
- Correa, P. Helicobacter pylori and gastric cancer: State of the art. Cancer Epidemiol. Biomark. Prev. 1996, 5, 477–481. [Google Scholar]
- Atherton, J.C.H. Pylori virulence factors. Br Med. Bull. 1998, 54, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.Y.; Yamaoka, Y. Disease-specific Helicobacter pylori Virulence Factors: The Unfulfilled Promise. Helicobacter 2000, 5, 3–9. [Google Scholar] [CrossRef]
- Bodger, K.; Crabtree, J.E. Helicobacter pylori and gastric inflammation. Br Med. Bull. 1998, 54, 139–150. [Google Scholar] [CrossRef]
- Crabtree, J.E. Role of cytokines in pathogenesis of Helicobacter pylori-induced mucosal damage. Dig. Dis. Sci. 1998, 43, 46S–55S. [Google Scholar]
- Harris, P.R.; Smythies, L.E.; Smith, P.D.; Dubois, A. Inflammatory Cytokine mRNA Expression during Early and Persistent Helicobacter pylori Infection in Nonhuman Primates. J. Infect. Dis. 2000, 181, 783–786. [Google Scholar] [CrossRef] [Green Version]
- Tomita, T.; Jackson, A.M.; Hida, N. Expression of Interleukin-18, a Th1 cytokine, in human gastric mucosa is increased in Helicobacter pylori infection. J. Infect. Dis. 2001, 183, 620–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Brooks, E.G.; Bamford, K.B. Negative selection of T cells by Helicobacter pylori as a model for bacterial strain selection by immune evasion. J. Immunol. 2001, 167, 926–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smythies, L.E.; Waites, K.B.; Lindsey, J.R.; Harris, P.R.; Ghiara, P.; Smith, P.D. Helicobacter pylori-Induced Mucosal Inflammation Is Th1 Mediated and Exacerbated in IL-4, But Not IFN-γ, Gene-Deficient Mice. J. Immunol. 2000, 165, 1022–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amedei, A.; Munari, F.; DELLA Bella, C.; Niccolai, E.; Benagiano, M.; Bencini, L.; Cianchi, F.; Farsi, M.; Emmi, G.; Zanotti, G.; et al. Helicobacter pylori secreted peptidyl prolyl cis, trans-isomerase drives Th17 inflammation in gastric adenocarcinoma. Intern. Emerg. Med. 2012, 9, 303–309. [Google Scholar] [CrossRef]
- Dixon, M.F.; Genta, R.M.; Yardley, J.H. Classification and grading of gastritis. The updated Sydney System. International Workshop on the Histopathology of Gastritis, Houston 1994. Am. J. Surg. Pathol. 1996, 20, 1161–1181. [Google Scholar] [CrossRef] [PubMed]
- Genta, R.M.; Lew, G.M.; Graham, D.Y. Changes in the gastric mucosa following eradication of Helicobacter pylori. Mod. Pathol. 1993, 6, 271–289. [Google Scholar]
- Fallone, C.A.; Chiba, N.; van Zanten, S.V.; Fischbach, L.; Gisbert, J.P.; Hunt, R.H.; Jones, N.L.; Render, C.; Leontiadis, G.I.; Moayyedi, P. The Toronto Consensus for the Treatment of Helicobacter pylori Infection in Adults. Gastroenterology 2016, 151, 51–69.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jha, S.K.; Mishra, M.K.; Saharawat, K.; Jha, P.; Purkayastha, S.; Ranjan, R. Comparison of concomitant therapy versus standard triple-drug therapy for eradication of Helicobacter pylori infection: A prospective open-label randomized controlled trial. Indian J. Gastroenterol. 2019, 38, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Sipponen, P.; Maaroos, H.I. Chronic gastritis. Scand. J. Gastroenterol. 2015, 50, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Sipponen, P.; Riihela, M.; Hyvarinen, H. Chronic nonatropic (’superficial’) gastritis increases the risk of gastric carcinoma. A case-control study. Scand. J. Gastroenterol. 1994, 29, 336–340. [Google Scholar] [CrossRef]
- Correa, P.; Haenszel, W.; Cuello, C. A model for gastric cancer epidemiology. Lancet 1975, 2, 58–60. [Google Scholar] [CrossRef]
- Rugge, M.; Capelle, L.G.; Cappellesso, R.; Nitti, D.; Kuipers, E.J. Precancerous lesions in the stomach: From biology to clinical patient management. Best Pr. Res. Clin. Gastroenterol. 2013, 27, 205–223. [Google Scholar] [CrossRef]
- Kuipers, E.J.; Lee, A.; Klinkenberg-Knol, E.C. Review article: The development of atrophic gastritis--Helicobacter pylori and the effects of acid suppressive therapy. Aliment Pharmacol. Ther. 1995, 9, 331–340. [Google Scholar] [CrossRef]
- Kuipers, E.J.; Uyterlinde, A.M.; Peña, A.S.; Hazenberg, H.J.; Bloemena, E.; Lindeman, J.; Klinkenberg-Knol, E.C.; Meuwissen, S.G. Increase of Helicobacter pylori-associated corpus gastritis during acid suppressive therapy: Implications for long-term safety. Am. J. Gastroenterol. 1995, 90, 1401–1406. [Google Scholar]
- Crabtree, J.; Taylor, J.; Heatley, R.; Shallcross, T.; Rathbone, B.; Wyatt, J.; Tompkins, D. Mucosal IgA recognition of Helicobacter pylori 120 kDa protein, peptic ulceration, and gastric pathology. Lancet 1991, 338, 332–335. [Google Scholar] [CrossRef]
- Kuipers, E.J.; Pérez-Pérez, G.I.; Meuwissen, S.G.M.; Blaser, M.J. Helicobacter pylori and Atrophic Gastritis: Importance of the cagA Status. J. Natl. Cancer Inst. 1995, 87, 1777–1780. [Google Scholar] [CrossRef] [PubMed]
- Parsonnet, J.; Friedman, G.D.; Orentreich, N.; Vogelman, H. Risk for gastric cancer in people with CagA positive or CagA negative Helicobacter pylori infection. Gut 1997, 40, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, U.C.; Tiwari, S.; Dhingra, S.; Pandey, R.; Ghoshal, U.; Tripathi, S.; Singh, H.; Gupta, V.K.; Nagpal, A.K.; Naik, S. Frequency of Helicobacter pylori and CagA Antibody in Patients with Gastric Neoplasms and Controls: The Indian Enigma. Dig. Dis. Sci. 2008, 53, 1215–1222. [Google Scholar] [CrossRef]
- Sipponen, P.; Kimura, K. Intestinal metaplasia, atrophic gastritis and stomach cancer: Trends over time. Eur. J. Gastroenterol. Hepatol. 1994, 6 (Suppl. 1), S79–S83. [Google Scholar] [PubMed]
- Sipponen, P.; Kekki, M.; Haapakoski, J.; Ihamäki, T.; Siurala, M. Gastric cancer risk in chronic atrophic gastritis: Statistical calculations of cross-sectional data. Int. J. Cancer 1985, 35, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Semino-Mora, C.; Doi, S.Q.; Marty, A.; Simko, V.; Carlstedt, I.; Dubois, A. Intracellular and Interstitial Expression ofHelicobacterpyloriVirulence Genes in Gastric Precancerous Intestinal Metaplasia and Adenocarcinoma. J. Infect. Dis. 2003, 187, 1165–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghoshal, U.C.; Kumar, S.; Krishnani, N.; Kumari, N.; Chourasia, D.; Tripathi, S. Serological assessment of gastric intestinal metaplasia and atrophy using pepsinogen-I, pepsinogen-II and gastrin-17 levels in a low incidence area of gastric cancer endemic for H. pylori infection. Trop. Gastroenterol. 2012, 32, 292–298. [Google Scholar]
- Weis, V.; Goldenring, J.R. Current understanding of SPEM and its standing in the preneoplastic process. Gastric Cancer 2009, 12, 189–197. [Google Scholar] [CrossRef]
- Stemmermann, G.N. Intestinal metaplasia of the stomach. A status report. Cancer 1994, 74, 556–564. [Google Scholar] [CrossRef]
- Jass, J.R.; Filipe, M.I. Sulphomucins and precancerous lesions of the human stomach. Histopathology 1980, 4, 271–279. [Google Scholar] [CrossRef]
- De Vries, A.C.; van Grieken, N.C.; Looman, C.W. Gastric cancer risk in patients with premalignant gastric lesions: A nationwide cohort study in the Netherlands. Gastroenterology 2008, 134, 945–952. [Google Scholar] [CrossRef]
- Filipe, M.I.; Muñoz, N.; Matko, I.; Kato, I.; Pompe-Kirn, V.; Jutersek, A.; Teuchmann, S.; Benz, M.; Prijon, T. Intestinal metaplasia types and the risk of gastric cancer: A cohort study in Slovenia. Int. J. Cancer 1994, 57, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Safatle-Ribeiro, A.V.; Ribeiro, U.; Clarke, M.R.; Sakai, P.; Ishioka, S.; Garrido, A.B.; Gama-Rodrigues, J.; Safatle, N.F.; Reynolds, J.C. Relationship between persistence of Helicobacter pylori and dysplasia, intestinal metaplasia, atrophy, inflammation, and cell proliferation following partial gastrectomy. Dig. Dis. Sci. 1999, 44, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Pulipati, P.; Sarkar, P.; Jakkampudi, A. The Indian gut microbiota-Is it unique? Indian. J. Gastroenterol. 2020, 39, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, U.C.; Goel, A.; Quigley, E.M.M. Gut microbiota abnormalities, small intestinal bacterial overgrowth, and non-alcoholic fatty liver disease: An emerging paradigm. Indian J. Gastroenterol. 2020, 39, 9–21. [Google Scholar] [CrossRef]
- Schulz, C.; Schütte, K.; Malfertheiner, P. Helicobacter pylori and Other Gastric Microbiota in Gastroduodenal Pathologies. Dig. Dis. 2016, 34, 210–216. [Google Scholar] [CrossRef]
- Schulz, C.; Schütte, K.; Mayerle, J.; Malfertheiner, P. The role of the gastric bacterial microbiome in gastric cancer: Helicobacter pylori and beyond. Ther. Adv. Gastroenterol. 2019, 12, 1756284819894062. [Google Scholar] [CrossRef] [Green Version]
- Tu, S.; Bhagat, G.; Cui, G. Overexpression of interleukin-1beta induces gastric inflammation and cancer and mobilizes myeloid-derived suppressor cells in mice. Cancer Cell. 2008, 14, 408–419. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.Y.; Chan, A.O.; Rashid, A. Interleukin-1beta increases the risk of gastric cancer through induction of aberrant DNA methylation in a mouse model. Oncol. Lett. 2016, 11, 2919–2924. [Google Scholar] [CrossRef] [Green Version]
- Lofgren, J.L.; Whary, M.T.; Ge, Z.; Muthupalani, S.; Taylor, N.S.; Mobley, M.; Potter, A.; Varro, A.; Eibach, D.; Suerbaum, S.; et al. Lack of Commensal Flora in Helicobacter pylori–Infected INS-GAS Mice Reduces Gastritis and Delays Intraepithelial Neoplasia. Gastroenterology 2011, 140, 210–220.e4. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.C.; Dangler, C.A.; Chen, D.; Goldenring, J.R.; Koh, T.; Raychowdhury, R.; Coffey, R.J.; Ito, S.; Varro, A.; Dockray, G.J.; et al. Synergistic interaction between hypergastrinemia and Helicobacter infection in a mouse model of gastric cancer. Gastroenterology 2000, 118, 36–47. [Google Scholar] [CrossRef]
- Ferreira, R.M.; Pereira-Marques, J.; Pinto-Ribeiro, I.; Costa, J.L.; Carneiro, F.; Machado, J.C.; Figueiredo, C. Gastric microbial community profiling reveals a dysbiotic cancer-associated microbiota. Gut 2017, 67, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Zhang, Y.; Gerhard, M.; Gao, J.-J.; Mejias-Luque, R.; Zhang, L.; Vieth, M.; Ma, J.-L.; Bajbouj, M.; Suchanek, S.; et al. Effect of Helicobacter pylori on gastrointestinal microbiota: A population-based study in Linqu, a high-risk area of gastric cancer. Gut 2019, 69, 1598–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Zhou, X.; Liu, X.; Ling, Z.; Ji, F. Role of the Gastric Microbiome in Gastric Cancer: From Carcinogenesis to Treatment. Front. Microbiol. 2021, 12, 641322. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shao, L.; Liu, X. Alterations of gastric mucosal microbiota across different stomach microhabitats in a cohort of 276 patients with gastric cancer. E Bio. Med. 2019, 40, 336–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, J.J.Y.; Coker, O.O.; Chu, E.; Szeto, C.H.; Luk, S.T.Y.; Lau, H.; Yu, J. Gastric microbes associated with gastric inflammation, atrophy and intestinal metaplasia 1 year after Helicobacter pylori eradication. Gut 2020, 69, 1572–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barra, W.F.; Sarquis, D.P.; Khayat, A.S. Gastric Cancer Microbiome. Pathobiology 2021, 88, 156–169. [Google Scholar] [CrossRef]
- Bravo, L.E.; van Doom, L.J.; Realpe, J.L. Virulence-associated genotypes of Helicobacter pylori: Do they explain the African enigma? Am. J. Gastroenterol. 2002, 97, 2839–2842. [Google Scholar] [CrossRef]
- Fox, J.G.; Beck, P.; Dangler, C.A.; Whary, M.T.; Wang, T.C.; Shi, H.N.; Nagler-Anderson, C. Concurrent enteric helminth infection modulates inflammation and gastric immune responses and reduces helicobacter-induced gastric atrophy. Nat. Med. 2000, 6, 536–542. [Google Scholar] [CrossRef]
- Fuenmayor-Boscán, A.; Hernández-Rincón, I.; Arismendi-Morillo, G.; Mengual, E.; Rivero, Z.; Romero, G.; Lizarzábal, M.; Álvarez-Mon, M. Changes in the severity of gastric mucosal inflammation associated with Helicobacter pylori in humans coinfected with intestinal helminths. Indian J. Gastroenterol. 2020, 39, 186–195. [Google Scholar] [CrossRef]
- Hussain, Z.; El-Omar, E.; Lee, Y.Y. Dual infective burden of Helicobacter pylori and intestinal parasites: Good or bad news for the host? Indian J. Gastroenterol. 2020, 39, 111–116. [Google Scholar] [CrossRef]
- Du, Y.; Agnew, A.; Ye, X.-P.; Robinson, P.A.; Forman, D.; Crabtree, J.E. Helicobacter pylori and Schistosoma japonicum co-infection in a Chinese population: Helminth infection alters humoral responses to H. pylori and serum pepsinogen I/II ratio. Microbes Infect. 2006, 8, 52–60. [Google Scholar] [CrossRef]
- Whary, M.T.; Sundina, N.; Bravo, L.E.; Correa, P.; Quiñones, F.; Caro, F.; Fox, J.G. Intestinal Helminthiasis in Colombian Children Promotes a Th2 Response to Helicobacter pylori: Possible Implications for Gastric Carcinogenesis. Cancer Epidemiol. Biomark. Prev. 2005, 14, 1464–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGinty, J.W.; Ting, H.-A.; Billipp, T.E.; Nadjsombati, M.S.; Khan, D.M.; Barrett, N.A.; Liang, H.-E.; Matsumoto, I.; von Moltke, J. Tuft-Cell-Derived Leukotrienes Drive Rapid Anti-helminth Immunity in the Small Intestine but Are Dispensable for Anti-protist Immunity. Immunity 2020, 52, 528–541.e7. [Google Scholar] [CrossRef]
- Von Moltke, J.; Ji, M.; Liang, H.E. Tuft-cell-derived IL-25 regulates an intestinal ILC2-epithelial response circuit. Nature 2016, 529, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Yousefi, B.; Mohammadlou, M.; Abdollahi, M.; Farrokhi, A.S.; Karbalaei, M.; Keikha, M.; Kokhaei, P.; Valizadeh, S.; Rezaiemanesh, A.; Arabkari, V.; et al. Epigenetic changes in gastric cancer induction by Helicobacter pylori. J. Cell. Physiol. 2019, 234, 21770–21784. [Google Scholar] [CrossRef] [PubMed]
- Minarovits, J. Microbe-induced epigenetic alterations in host cells: The coming era of patho-epigenetics of microbial infections. Acta Microbiol. Immunol. Hung. 2009, 56, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Ghoshal, U.C.; Tripathi, S.; Ghoshal, U. The Indian enigma of frequent H. pylori infection but infrequent gastric cancer: Is the magic key in Indian diet, host’s genetic make up, or friendly bug? Am. J. Gastroenterol. 2007, 102, 2113–2114. [Google Scholar] [CrossRef] [PubMed]
- Parsonnet, J.; Friedman, G.D.; Vandersteen, D.P.; Chang, Y.; Vogelman, J.H.; Orentreich, N.; Sibley, R.K. Helicobacter pyloriInfection and the Risk of Gastric Carcinoma. N. Engl. J. Med. 1991, 325, 1127–1131. [Google Scholar] [CrossRef] [PubMed]
- Jakszyn, P.; Gonzalez, C.A. Nitrosamine and related food intake and gastric and oesophageal cancer risk: A systematic review of the epidemiological evidence. World J. Gastroenterol. 2006, 12, 4296–4303. [Google Scholar] [CrossRef]
- Butt, J.; Varga, M.G.; Wang, T.; Tsugane, S.; Shimazu, T.; Zheng, W.; Abnet, C.C.; Yoo, K.-Y.; Park, S.K.; Kim, J.; et al. Smoking, Helicobacter Pylori Serology, and Gastric Cancer Risk in Prospective Studies from China, Japan, and Korea. Cancer Prev. Res. 2019, 12, 667–674. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Metz, D.C.; Ellenberg, S.; Kaplan, D.E.; Goldberg, D.S. Risk Factors and Incidence of Gastric Cancer After Detection of Helicobacter pylori Infection: A Large Cohort Study. Gastroenterology 2020, 158, 527–536.e7. [Google Scholar] [CrossRef] [Green Version]
- Van Hecke, T.; van Camp, J.; De Smet, S. Oxidation During Digestion of Meat: Interactions with the Diet and Helicobacter pylori Gastritis, and Implications on Human Health. Compr. Rev. Food Sci. Food Saf. 2017, 16, 214–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Torre, G.; Chiaradia, G.; Gianfagna, F. Smoking status and gastric cancer risk: An updated meta-analysis of case-control studies published in the past ten years. Tumori 2009, 95, 13–22. [Google Scholar] [CrossRef]
- Ladeiras-Lopes, R.; Pereira, A.K.; Nogueira, A.; Pinheiro-Torres, T.; Pinto, I.; Santos-Pereira, R.; Lunet, N. Smoking and gastric cancer: Systematic review and meta-analysis of cohort studies. Cancer Causes Control. 2008, 19, 689–701. [Google Scholar] [CrossRef] [PubMed]
- D’Elia, L.; Rossi, G.; Ippolito, R.; Cappuccio, F.P.; Strazzullo, P. Habitual salt intake and risk of gastric cancer: A meta-analysis of prospective studies. Clin. Nutr. 2012, 31, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, L.; Wang, X.; Wang, J.; Yan, Z.; Cheng, J.; Gong, G.; Li, G. Body Mass Index and Risk of Gastric Cancer: A Meta-analysis of a Population with More Than Ten Million from 24 Prospective Studies. Cancer Epidemiol. Biomark. Prev. 2013, 22, 1395–1408. [Google Scholar] [CrossRef] [Green Version]
- López-Carrillo, L.; López-Cervantes, M.; Ramírez-Espitia, A.; Rueda, C.; Fernández-Ortega, C.; Orozco-Rivadeneyra, S. Alcohol consumption and gastric cancer in Mexico. Cad. Saude Publica 1998, 14, S25–S32. [Google Scholar] [CrossRef] [Green Version]
- Clinton, S.K.; Giovannucci, E.L.; Hursting, S.D. The World Cancer Research Fund/American Institute for Cancer Research Third Expert Report on Diet, Nutrition, Physical Activity, and Cancer: Impact and Future Directions. J. Nutr. 2019, 150, 663–671. [Google Scholar] [CrossRef]
- González, C.A.; Pera, G.; Agudo, A.; Bueno-De-Mesquita, H.B.; Ceroti, M.; Boeing, H.; Schulz, M.; Del Giudice, G.; Plebani, M.; Carneiro, F.; et al. Fruit and vegetable intake and the risk of stomach and oesophagus adenocarcinoma in the European Prospective Investigation into Cancer and Nutrition (EPIC–EURGAST). Int. J. Cancer 2006, 118, 2559–2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serafini, M.; Bellocco, R.; Wolk, A.; Ekström, A.M. Total antioxidant potential of fruit and vegetables and risk of gastric cancer. Gastroenterology 2002, 123, 985–991. [Google Scholar] [CrossRef]
- Foryst-Ludwig, A.; Neumann, M.; Schneider-Brachert, W. Curcumin blocks NF-kappaB and the motogenic response in Helicobacter pylori-infected epithelial cells. Biochem. Biophys. Res. Commun. 2004, 316, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Ahuja, V.; Ghoshal, U.C.; Makharia, G.; Dutta, U.; Zargar, S.A.; Venkataraman, J.; Dutta, A.K.; Mukhopadhyay, A.K.; Singh, A.; et al. Management of Helicobacter pylori infection: The Bhubaneswar Consensus Report of the Indian Society of Gastroenterology. Indian J. Gastroenterol. 2021, 1–25. [Google Scholar] [CrossRef]
- Okada, F.; Izutsu, R.; Goto, K.; Osaki, M. Inflammation-Related Carcinogenesis: Lessons from Animal Models to Clinical Aspects. Cancers 2021, 13, 921. [Google Scholar] [CrossRef] [PubMed]
- Machado, J.C.; Figueiredo, C.; Canedo, P.; Pharoah, P.; Carvalho, R.; Nabais, S.; Alves, C.C.; Campos, M.L.; Van Doorn, L.-J.; Caldas, C.; et al. A proinflammatory genetic profile increases the risk for chronic atrophic gastritis and gastric carcinoma. Gastroenterology 2003, 125, 364–371. [Google Scholar] [CrossRef]
- Graham, D.Y. Helicobacter pylori Update: Gastric Cancer, Reliable Therapy, and Possible Benefits. Gastroenterology 2015, 148, 719–731.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forman, D.; Newell, D.G.; Fullerton, F.; Yarnell, J.W.; Stacey, A.R.; Wald, N.; Sitas, F. Association between infection with Helicobacter pylori and risk of gastric cancer: Evidence from a prospective investigation. BMJ 1991, 302, 1302–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, M.; Furuta, T.; Yamaoka, Y. Influence of inflammatory cytokine polymorphisms on eradication rates ofHelicobacter pylori. J. Gastroenterol. Hepatol. 2009, 24, 1725–1732. [Google Scholar] [CrossRef] [Green Version]
- White, J.R.; Winter, J.A.; Robinson, K. Differential inflammatory response to Helicobacter pylori infection: Etiology and clinical outcomes. J. Inflamm. Res. 2015, 8, 137–147. [Google Scholar]
- Morse, H.; Olomolaiye, O.; Wood, N.; Keen, L.; Bidwell, J. Induced Heteroduplex Genotyping of Tnf-A, Il-1β, Il-6 And Il-10 Polymorphisms Associated With Transcriptional Regulation. Cytokine 1999, 11, 789–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidwell, J.; Keen, L.; Gallagher, G.; Kimberly, R.; Huizinga, T.; McDermott, M.; Oksenberg, J.; McNicholl, J.; Pociot, F.; Hardt, C.; et al. Cytokine gene polymorphism in human disease: Online databases. Genes Immun. 1999, 1, 3–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuzhalin, A. The role of interleukin DNA polymorphisms in gastric cancer. Hum. Immunol. 2011, 72, 1128–1136. [Google Scholar] [CrossRef]
- Sugimoto, M.; Yamaoka, Y.; Furuta, T. Influence of interleukin polymorphisms on development of gastric cancer and peptic ulcer. World J. Gastroenterol. 2010, 16, 1188–1200. [Google Scholar] [CrossRef]
- Smoot, D.T.; Elliott, T.B.; Verspaget, H.W.; Jones, D.; Allen, C.R.; Vernon, K.G.; Bremner, T.; Kidd, L.C.R.; Kim, K.S.; Groupman, J.D.; et al. Influence of Helicobacter pylori on reactive oxygen-induced gastric epithelial cell injury. Carcinogenesis 2000, 21, 2091–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, I.R.; Kodama, T.; Kikuchi, S. Effect of interleukin 1 polymorphisms on gastric mucosal interleukin 1beta production in Helicobacter pylori infection. Gastroenterology 2002, 123, 1793–1803. [Google Scholar] [CrossRef]
- Wolfe, M.; Nompleggi, D. Cytokine inhibition of gastric acid secretion—A little goes a long way. Gastroenterology 1992, 102, 2177–2178. [Google Scholar] [CrossRef]
- El-Omar, E.; Carrington, M.; Chow, W.-H.; McColl, K.E.L.; Bream, J.; Young, H.A.; Herrera, J.; Lissowska, J.; Yuan, C.-C.; Rothman, N.; et al. Interleukin-1 polymorphisms associated with increased risk of gastric cancer. Nature 2000, 404, 398–402. [Google Scholar] [CrossRef] [Green Version]
- Kahroba, H.; Hejazi, M.S.; Samadi, N. Exosomes: From carcinogenesis and metastasis to diagnosis and treatment of gastric cancer. Cell. Mol. Life Sci. 2019, 76, 1747–1758. [Google Scholar] [CrossRef]
- Patel, G.K.; Khan, M.A.; Bhardwaj, A.; Srivastava, S.K.; Zubair, H.; Patton, M.C.; Singh, S.; Khushman, M.; Singh, A. Exosomes confer chemoresistance to pancreatic cancer cells by promoting ROS detoxification and miR-155-mediated suppression of key gemcitabine-metabolising enzyme, DCK. Br. J. Cancer 2017, 116, 609–619. [Google Scholar] [CrossRef] [Green Version]
- Che, Y.; Geng, B.; Xu, Y.; Miao, X.; Chen, L.; Mu, X.; Pan, J.; Zhang, C.; Zhao, T.; Wang, C.; et al. Helicobacter pylori -induced exosomal MET educates tumour-associated macrophages to promote gastric cancer progression. J. Cell. Mol. Med. 2018, 22, 5708–5719. [Google Scholar] [CrossRef]
- Shimoda, A.; Ueda, K.; Nishiumi, S.; Murata-Kamiya, N.; Mukai, S.-A.; Sawada, S.-I.; Azuma, T.; Hatakeyama, M.; Akiyoshi, K. Exosomes as nanocarriers for systemic delivery of the Helicobacter pylori virulence factor CagA. Sci. Rep. 2016, 6, 18346. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Cheng, J.; Luo, S.; Gong, X.; Huang, D.; Xu, J.; Qian, Y.; Wan, X.; Zhou, H. Deoxycholic acid-stimulated macrophage-derived exosomes promote spasmolytic polypeptide-expressing metaplasia in the stomach. Biochem. Biophys. Res. Commun. 2020, 524, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Chen, L.; Yuan, X.; Luo, Q.; Liu, Y.; Xie, G.; Ma, Y.; Shen, L. Exosomal transfer of tumor-associated macrophage-derived miR-21 confers cisplatin resistance in gastric cancer cells. J. Exp. Clin. Cancer Res. 2017, 36, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, J.-L.; Qu, X.-J.; Zhao, M.-F.; Teng, Y.-E.; Zhang, Y.; Hou, K.-Z.; Jiang, Y.-H.; Yang, X.-H.; Liu, Y.-P. Gastric cancer exosomes promote tumour cell proliferation through PI3K/Akt and MAPK/ERK activation. Dig. Liver Dis. 2009, 41, 875–880. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, X.; Zhang, B. Exosomes derived from gastric cancer cells activate NF-kappa B pathway in macrophages to promote cancer progression. Tumour Biol. 2016, 37, 12169–12180. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Liang, W.; Fu, M.; Huang, Z.-H.; Li, X.; Zhang, W.; Zhang, P.; Qian, H.; Jiang, P.-C.; Xu, W.-R.; et al. Exosomes-mediated transfer of long noncoding RNA ZFAS1 promotes gastric cancer progression. J. Cancer Res. Clin. Oncol. 2017, 143, 991–1004. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Peng, L.; Yang, J. Exosomal transfer of miR-15b-3p enhances tumorigenesis and malignant transformation through the DYNLT1/Caspase-3/Caspase-9 signaling pathway in gastric cancer. J. Exp. Clin. Cancer Res. 2020, 39, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, J.; Li, X.; Zhao, L. The role of PKM2 nuclear translocation in the constant activation of the NF-kappa B signaling pathway in cancer-associated fibroblasts. Cell Death Dis. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, T.; Liu, R.; Ning, T.; Yang, H.; Liu, D.; Zhang, Q.; Lin, D.; Ge, S.; Bai, M.; et al. CAF secreted miR-522 suppresses ferroptosis and promotes acquired chemo-resistance in gastric cancer. Mol. Cancer 2020, 19, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Shi, H.; Yuan, X.; Jiang, P.; Qian, H.; Xu, W. Tumor-derived exosomes induce N2 polarization of neutrophils to promote gastric cancer cell migration. Mol. Cancer 2018, 17, 1–16. [Google Scholar] [CrossRef] [PubMed]
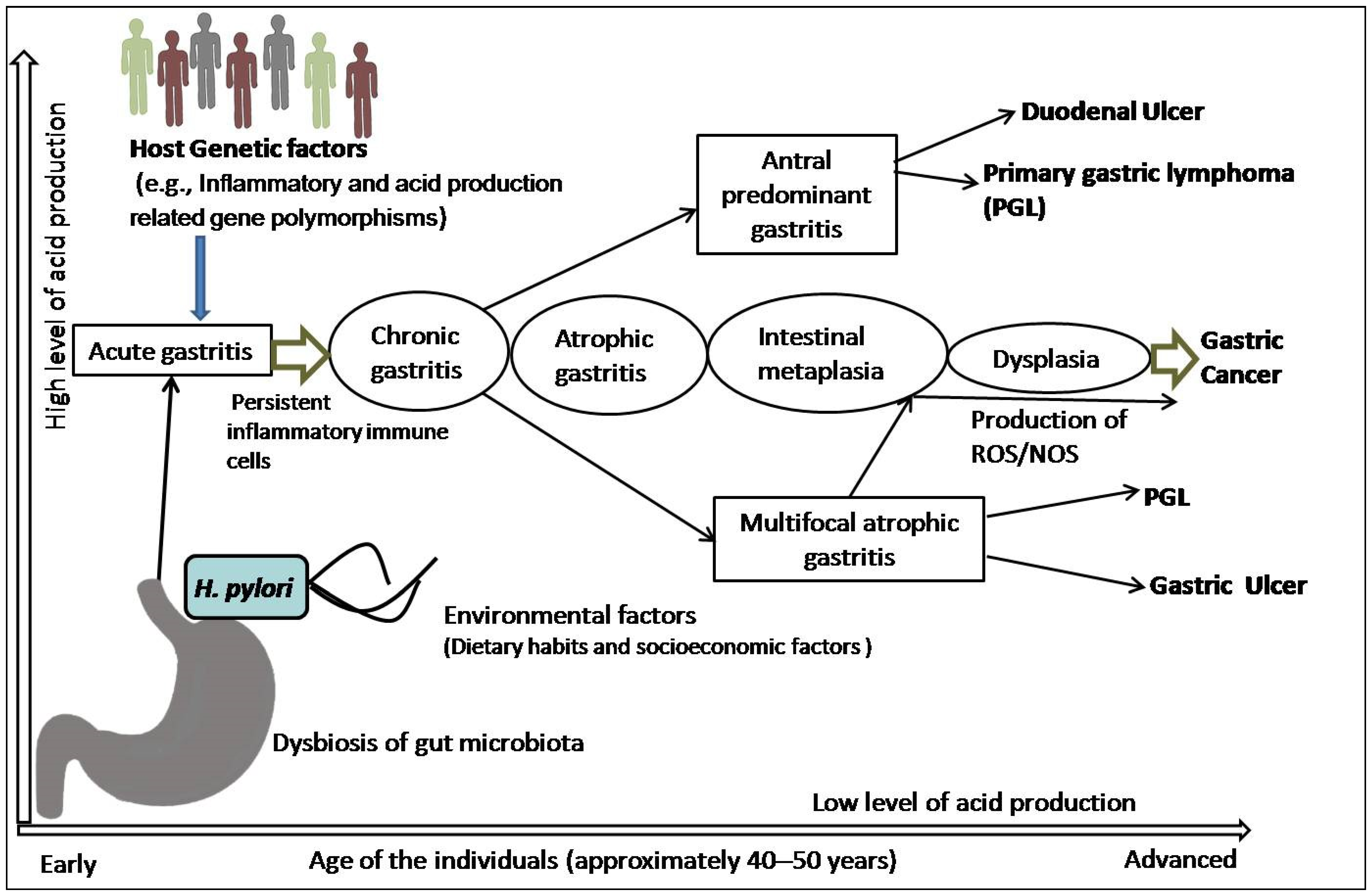
Figure 1.
Schematic representation of pathophysiology of gastric cancer: H. pylori infection and the combination of inflammatory and acid-production-related genetic polymorphisms, including dietary and socioeconomic variables, lead to development of acute gastritis that subsequently becomes chronic gastritis in the presence of persistent inflammatory immune cells. In a subset of patients, this chronic inflammation with antral and corpus localization develops peptic (duodenal and gastric) ulcer or primary gastric lymphoma (PGL). However, dysbiosis, interactions with other variables, and the generation of reactive oxygen species (ROS)/nitrogen oxygen species (NOS) cause atrophic gastritis, which is followed by intestinal metaplasia, dysplasia, and carcinoma.
Figure 1.
Schematic representation of pathophysiology of gastric cancer: H. pylori infection and the combination of inflammatory and acid-production-related genetic polymorphisms, including dietary and socioeconomic variables, lead to development of acute gastritis that subsequently becomes chronic gastritis in the presence of persistent inflammatory immune cells. In a subset of patients, this chronic inflammation with antral and corpus localization develops peptic (duodenal and gastric) ulcer or primary gastric lymphoma (PGL). However, dysbiosis, interactions with other variables, and the generation of reactive oxygen species (ROS)/nitrogen oxygen species (NOS) cause atrophic gastritis, which is followed by intestinal metaplasia, dysplasia, and carcinoma.

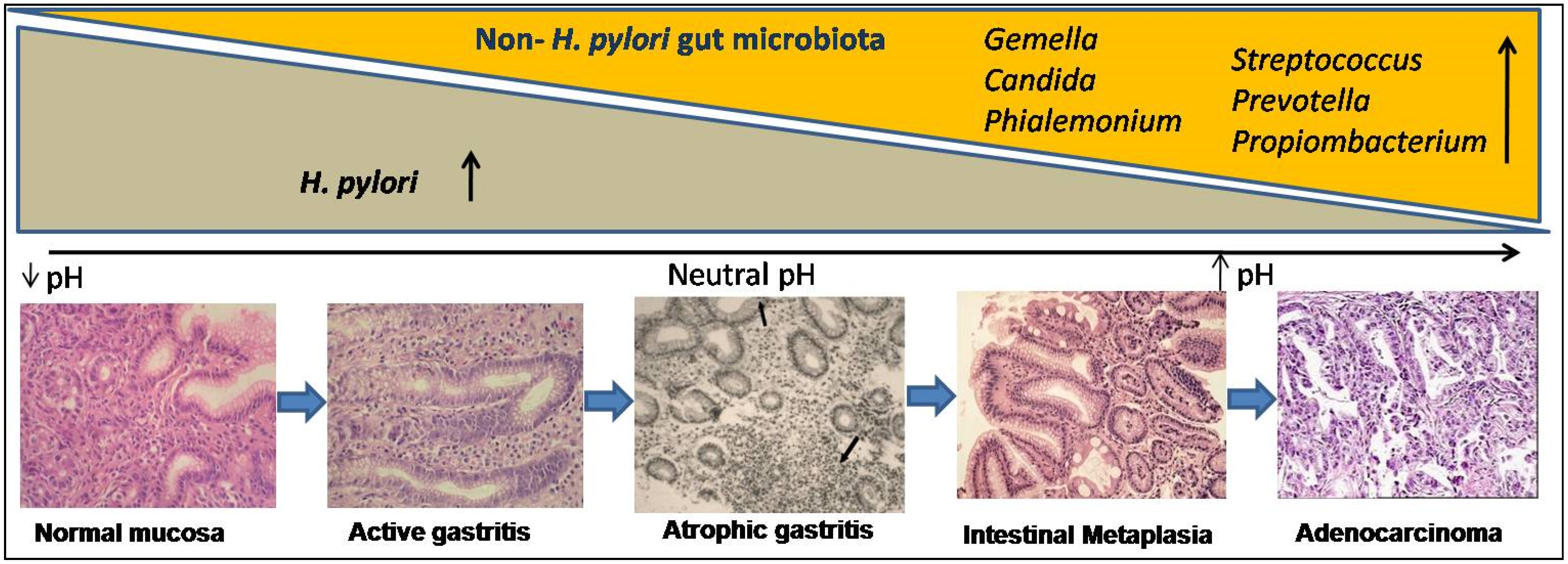
Figure 2.
Gastric microbial dysbiosis process:in the histological course of gastric carcinogenesis (at the bottom), H. pylori-dependent and independent phases showed exponential changes in gut microbiota (above). H. pylori-infected gastric mucosa shows low pH and after years to decades of colonization, H. pylori disappear and progressive atrophic gastritis, which reduces gastric acid secretion, raising the intra-gastric pH, favors overgrowing of non-H. pylori microbiota (i.e., Gemella, Candida, Streptococcus, Prevotella, Propiombacterium, and Phialemonium), which have carcinogenic potential.
Figure 2.
Gastric microbial dysbiosis process:in the histological course of gastric carcinogenesis (at the bottom), H. pylori-dependent and independent phases showed exponential changes in gut microbiota (above). H. pylori-infected gastric mucosa shows low pH and after years to decades of colonization, H. pylori disappear and progressive atrophic gastritis, which reduces gastric acid secretion, raising the intra-gastric pH, favors overgrowing of non-H. pylori microbiota (i.e., Gemella, Candida, Streptococcus, Prevotella, Propiombacterium, and Phialemonium), which have carcinogenic potential.

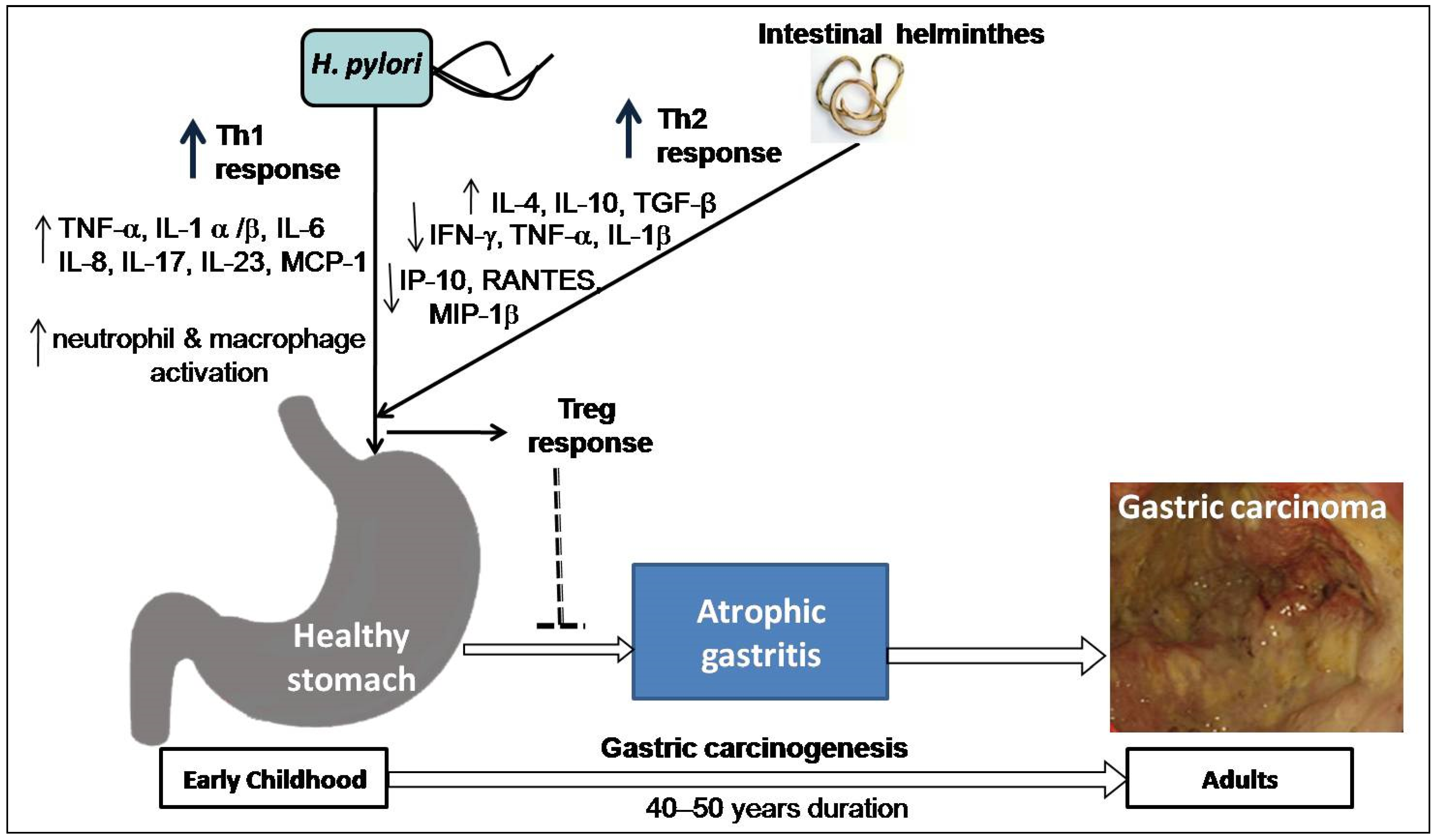
Figure 3.
Hypothesis on the impact of co-existence of H. pylori and intestinal helminths: both H. pylori and intestinal helminth infection interact in early childhood, resulting in change in Th1 response caused by H. pylori primarily upregulation of tumor necrosis factor (TNF)- α, interleukin (IL)-1 α/β, IL-6, IL-8, IL-17, IL-23 and monocyte chemoattractant protein (MCP-1) as well as activation of neutrophil and macrophage and Th2-polarizing response by helminthes mainly upregulation of IL-4, IL-10, Transforming growth factor (TGF)-β and downregulation of interferon (IFN)-γ, TNF-α, and IL-1β, including Th1-associated interferon gamma-induced protein 10 (IP-10), Regulated on Activation, Normal T Expressed and Secreted (RANTES) protein, and Macrophage inflammatory protein (MIP)-1β. Thus, T regulatory (Treg) response inhibits the development of atrophic gastritis and possibly lowers cancer risk in later life (after 40–50 years).
Figure 3.
Hypothesis on the impact of co-existence of H. pylori and intestinal helminths: both H. pylori and intestinal helminth infection interact in early childhood, resulting in change in Th1 response caused by H. pylori primarily upregulation of tumor necrosis factor (TNF)- α, interleukin (IL)-1 α/β, IL-6, IL-8, IL-17, IL-23 and monocyte chemoattractant protein (MCP-1) as well as activation of neutrophil and macrophage and Th2-polarizing response by helminthes mainly upregulation of IL-4, IL-10, Transforming growth factor (TGF)-β and downregulation of interferon (IFN)-γ, TNF-α, and IL-1β, including Th1-associated interferon gamma-induced protein 10 (IP-10), Regulated on Activation, Normal T Expressed and Secreted (RANTES) protein, and Macrophage inflammatory protein (MIP)-1β. Thus, T regulatory (Treg) response inhibits the development of atrophic gastritis and possibly lowers cancer risk in later life (after 40–50 years).

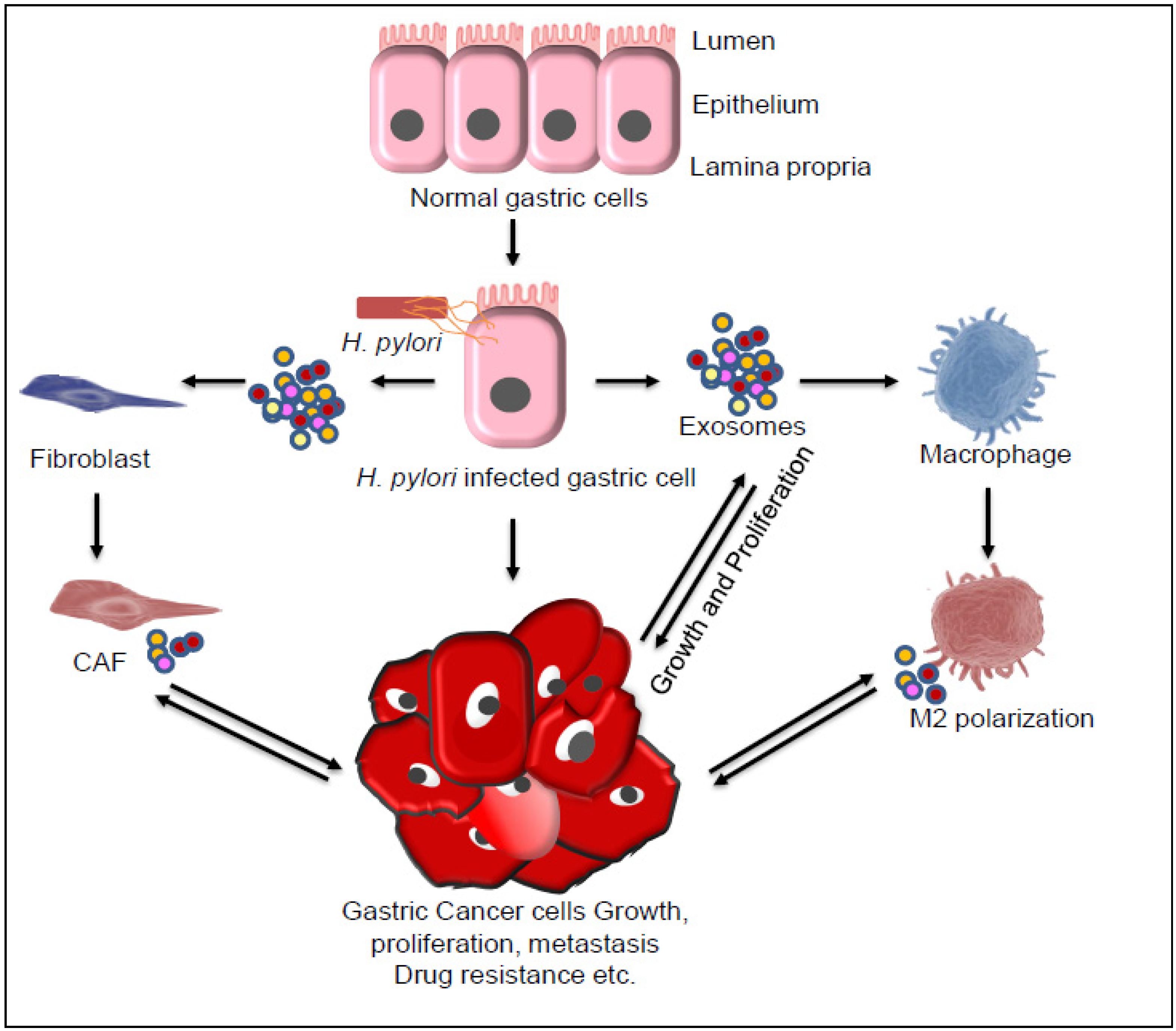
Figure 4.
Exosome-mediated inter-cellular communications lead to gastric cancer (GC) progression. Different genetic and epigenetic factors and H. pylori infection alone or in combination lead to development of GC. Exosome derived from H.pylori or from infected gastric cells, cancer-associated fibroblasts (CAF), and macrophage marker (M2 macrophage) cells plays an important role in GC progression through different pathways by transfer of vital molecules, including Cytotoxin-associated antigen A (CagA), mesenchymal epithelial transition factor (MET), TFF2 and GSII ApoE (biomarkers of spasmolytic polypeptide-expressing metaplasia [SPEM]), high mobility group box-1 (HMGB1), pyruvate kinase M2 (PKM2), long noncoding RNA, and microRNA. Besides exosomes, the secreted inflammatory cytokines, such as interleukin (IL)-6, IL-8, and IL-1β also play a major role in cancer cell growth proliferation, metastasis, and drug resistance.
Figure 4.
Exosome-mediated inter-cellular communications lead to gastric cancer (GC) progression. Different genetic and epigenetic factors and H. pylori infection alone or in combination lead to development of GC. Exosome derived from H.pylori or from infected gastric cells, cancer-associated fibroblasts (CAF), and macrophage marker (M2 macrophage) cells plays an important role in GC progression through different pathways by transfer of vital molecules, including Cytotoxin-associated antigen A (CagA), mesenchymal epithelial transition factor (MET), TFF2 and GSII ApoE (biomarkers of spasmolytic polypeptide-expressing metaplasia [SPEM]), high mobility group box-1 (HMGB1), pyruvate kinase M2 (PKM2), long noncoding RNA, and microRNA. Besides exosomes, the secreted inflammatory cytokines, such as interleukin (IL)-6, IL-8, and IL-1β also play a major role in cancer cell growth proliferation, metastasis, and drug resistance.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kumar, S.; Patel, G.K.; Ghoshal, U.C. Helicobacter pylori-Induced Inflammation: Possible Factors Modulating the Risk of Gastric Cancer. Pathogens 2021, 10, 1099. https://doi.org/10.3390/pathogens10091099
AMA Style
Kumar S, Patel GK, Ghoshal UC. Helicobacter pylori-Induced Inflammation: Possible Factors Modulating the Risk of Gastric Cancer. Pathogens. 2021; 10(9):1099. https://doi.org/10.3390/pathogens10091099
Chicago/Turabian StyleKumar, Sushil, Girijesh Kumar Patel, and Uday C. Ghoshal. 2021. "Helicobacter pylori-Induced Inflammation: Possible Factors Modulating the Risk of Gastric Cancer" Pathogens 10, no. 9: 1099. https://doi.org/10.3390/pathogens10091099
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.