
Advax-CpG Adjuvant Provides Antigen Dose-Sparing and Enhanced Immunogenicity for Inactivated Poliomyelitis Virus Vaccines


Abstract
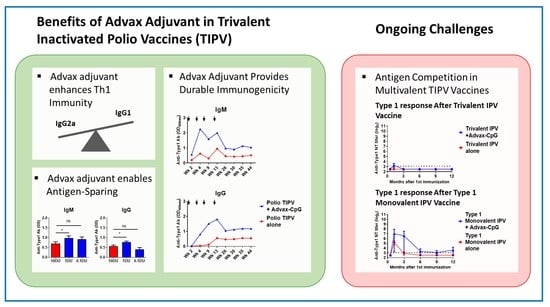
:
1. Introduction
2. Methods
2.1. Animals
2.2. Vaccines and Adjuvants
2.3. Immunization Schedule for Mouse and Rat Studies
2.4. ELISA for Detection of Anti-IPV Antibodies
2.5. Neutralization Assays
2.6. Statistical Analysis
3. Results
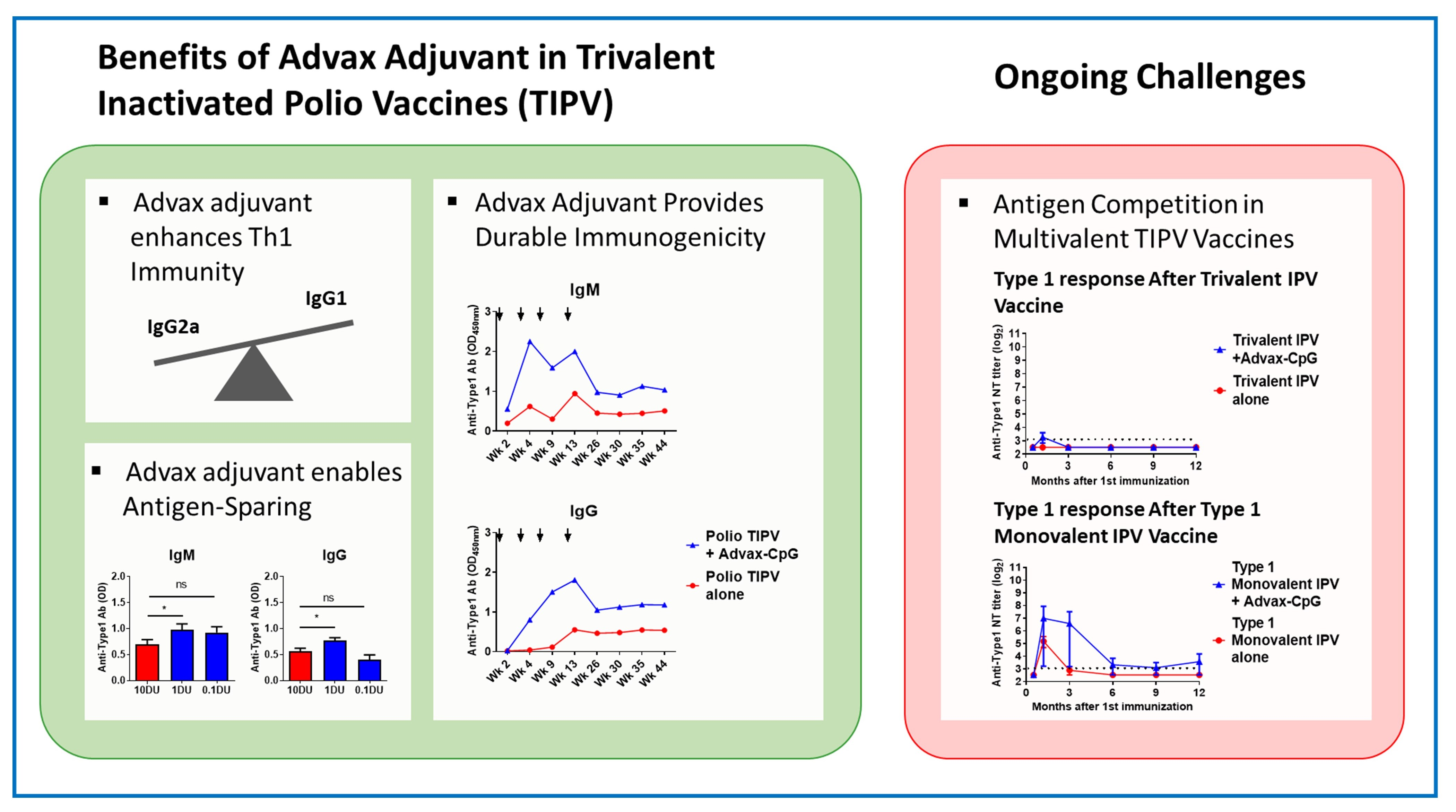
3.1. Advax-CpG Improves IPOL® TIPV Vaccine Long-Term
3.2. Advax-CpG Induces a Strong Th1 Immune Response to IPOL® TIPV Vaccine
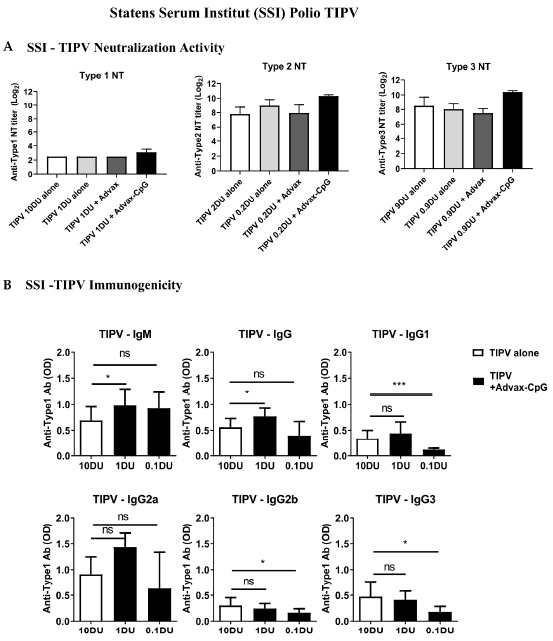
3.3. Dose-Sparing Effects of Advax-CpG Adjuvant on SSI TIPV Immunogenicity
3.4. Effects of Advax Adjuvants on Antibody Kinetics and Longevity
3.5. TIPV Vaccine Formulated with Advax-CpG Adjuvant Induces Protective Antibody Levels in Rats
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baicus, A. History of polio vaccination. World J. Virol. 2012, 1, 108–114. [Google Scholar] [CrossRef]
- Bandyopadhyay, A.S.; Garon, J.; Seib, K.; Orenstein, W.A. Polio vaccination: Past, present and future. Future Microbiol. 2015, 10, 791–808. [Google Scholar] [CrossRef] [Green Version]
- Jiang, B.; Patel, M.; Glass, R.I. Polio endgame: Lessons for the global rotavirus vaccination program. Vaccine 2019, 37, 3040–3049. [Google Scholar] [CrossRef]
- Alfaro-Murillo, J.A.; Ávila-Agüero, M.L.; Fitzpatrick, M.C.; Crystal, C.J.; Falleiros-Arlant, L.-H.; Galvani, A.P. The case for replacing live oral polio vaccine with inactivated vaccine in the Americas. Lancet 2020, 395, 1163–1166. [Google Scholar] [CrossRef]
- Shahzad, A.; Köhler, G. Inactivated Polio Vaccine (IPV): A strong candidate vaccine for achieving global polio eradication program. Vaccine 2009, 27, 5293–5294. [Google Scholar] [CrossRef]
- Vidor, E.; Meschievitz, C.; Plotkin, S. Fifteen years of experience with Vero-produced enhanced potency inactivated poliovirus vaccine. Pediatr. Infect. Dis. J. 1997, 16, 312–322. [Google Scholar] [CrossRef]
- Hamer, D.H.; Griffiths, J.; Maguire, J.H.; Heggenhougen, K.; Quah, S.R. Public Health and Infectious Diseases; Elsevier Science: Oxford, UK, 2010. [Google Scholar]
- Bonnet, M.-C.; Dutta, A. World wide experience with inactivated poliovirus vaccine. Vaccine 2008, 26, 4978–4983. [Google Scholar] [CrossRef]
- Baldwin, S.L.; Fox, C.B.; Pallansch, M.A.; Coler, R.N.; Reed, S.G.; Friede, M. Increased potency of an inactivated trivalent polio vaccine with oil-in-water emulsions. Vaccine 2011, 29, 644–649. [Google Scholar] [CrossRef]
- Sureau, P.; Fabre, P.S.; N’Garo, S.B.; Butor, S.C. Simultaneous Testanus and Poliomyelitis Vaccination of Infants in a Tropical Environment. Bull. World Health Organ. 1977, 55, 739–746. [Google Scholar]
- Drescher, J.; Grutzner, L.; Godgluck, G. Further Investigations on the Immunogenic Activity of Aqueous and Aluminium Oxide Adsorbed Inactivated Poliovirus Vaccines in Macaca Mulatto. Am. J. Epidemiol. 1967, 85, 413–423. [Google Scholar] [CrossRef]
- Ivanov, A.P.; Dragunsky, E.M.; Chumakov, K.M. 1,25-Dihydroxyvitamin D3 Enhances Systemic and Mucosal Immune Responses to Inactivated Poliovirus Vaccine in Mice. J. Infect. Dis. 2006, 193, 598–600. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Shi, H.; Zhou, J.; Liang, Y.; Xu, H. CpG oligodeoxynucleotides are a potent adjuvant for an inactivated polio vaccine produced from Sabin strains of poliovirus. Vaccine 2009, 27, 6558–6563. [Google Scholar] [CrossRef]
- Petrovsky, N.; Cooper, P.D. Advax™, a novel microcrystalline polysaccharide particle engineered from delta inulin, provides robust adjuvant potency together with tolerability and safety. Vaccine 2015, 33, 5920–5926. [Google Scholar] [CrossRef]
- Honda-Okubo, Y.; Saade, F.; Petrovsky, N. Advax™, a polysaccharide adjuvant derived from delta inulin, provides improved influenza vaccine protection through broad-based enhancement of adaptive immune responses. Vaccine 2012, 30, 5373–5381. [Google Scholar] [CrossRef] [Green Version]
- Gordon, D.L.; Sajkov, D.; Honda-Okubo, Y.; Wilks, S.H.; Aban, M.; Barr, I.G.; Petrovsky, N. Human Phase 1 trial of low-dose inactivated seasonal influenza vaccine formulated with Advax™ delta inulin adjuvant. Vaccine 2016, 34, 3780–3786. [Google Scholar] [CrossRef]
- Gordon, D.L.; Sajkov, D.; Woodman, R.J.; Honda-Okubo, Y.; Cox, M.M.; Heinzel, S.; Petrovsky, N. Randomized Clinical Trial of Immunogenicity and Safety of a Recombinant H1n1/2009 Pandemic Influenza Vaccine Containing Advax™ Polysaccharide Adjuvant. Vaccine 2012, 30, 5407–5416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobigs, M.; Pavy, M.; Hall, R.A.; Lobigs, P.; Cooper, P.; Komiya, T.; Toriniwa, H.; Petrovsky, N. An inactivated Vero cell-grown Japanese encephalitis vaccine formulated with Advax, a novel inulin-based adjuvant, induces protective neutralizing antibody against homologous and heterologous flaviviruses. J. Gen. Virol. 2010, 91, 1407–1417. [Google Scholar] [CrossRef]
- Saade, F.; Honda-Okubo, Y.; Trec, S.; Petrovsky, N. A novel hepatitis B vaccine containing Advax™, a polysaccharide adjuvant derived from delta inulin, induces robust humoral and cellular immunity with minimal reactogenicity in preclinical testing. Vaccine 2013, 31, 1999–2007. [Google Scholar] [CrossRef] [Green Version]
- Cosseddu, G.M.; Polci, A.; Pinoni, C.; Dondona, A.C.; Iapaolo, F.; Orsini, G.; Izzo, F.; Bortone, G.; Ronchi, F.G.; Di Ventura, M.; et al. Evaluation of Humoral Response and Protective Efficacy of an Inactivated Vaccine Against Peste des Petits Ruminants Virus in Goats. Transbound. Emerg. Dis. 2015, 63, e447–e452. [Google Scholar] [CrossRef]
- Honda-Okubo, Y.; Kolpe, A.; Li, L.; Petrovsky, N. A single immunization with inactivated H1N1 influenza vaccine formulated with delta inulin adjuvant (Advax™) overcomes pregnancy-associated immune suppression and enhances passive neonatal protection. Vaccine 2014, 32, 4651–4659. [Google Scholar] [CrossRef] [Green Version]
- Bielefeldt-Ohmann, H.; Prow, N.A.; Wang, W.; Tan, C.S.; Coyle, M.; Douma, A.; Hobson-Peters, J.; Kidd, L.; Hall, R.A.; Petrovsky, N. Safety and immunogenicity of a delta inulin-adjuvanted inactivated Japanese encephalitis virus vaccine in pregnant mares and foals. Vet. Res. 2014, 45, 130. [Google Scholar] [CrossRef] [Green Version]
- Honda-Okubo, Y.; Ong, C.H.; Petrovsky, N. Advax delta inulin adjuvant overcomes immune immaturity in neonatal mice thereby allowing single–dose influenza vaccine protection. Vaccine 2015, 33, 4892–4900. [Google Scholar] [CrossRef]
- Chu, R.S.; Targoni, O.S.; Krieg, A.M.; Lehmann, P.V.; Harding, C.V. CpG Oligodeoxynucleotides Act as Adjuvants that Switch on T Helper 1 (Th1) Immunity. J. Exp. Med. 1997, 186, 1623–1631. [Google Scholar] [CrossRef] [Green Version]
- Counoupas, C.; Pinto, R.; Nagalingam, G.; Britton, W.J.; Petrovsky, N.; Triccas, J.A. Delta inulin-based adjuvants promote the generation of polyfunctional CD4+ T cell responses and protection against Mycobacterium tuberculosis infection. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Stronsky, S.M.; Cooper, C.L.; Steffens, J.; Van Tongeren, S.; Bavari, S.; Martins, K.A.; Petrovsky, N. Adjuvant selection impacts the correlates of vaccine protection against Ebola infection. Vaccine 2020, 38, 4601–4608. [Google Scholar] [CrossRef]
- Honda-Okubo, Y.; Barnard, D.L.; Ong, C.H.; Peng, B.-H.; Tseng, C.-T.K.; Petrovsky, N. Severe Acute Respiratory Syndrome-Associated Coronavirus Vaccines Formulated with Delta Inulin Adjuvants Provide Enhanced Protection while Ameliorating Lung Eosinophilic Immunopathology. J. Virol. 2014, 89, 2995–3007. [Google Scholar] [CrossRef] [Green Version]
- Davtyan, H.; Zagorski, K.; Rajapaksha, H.; Hovakimyan, A.; Davtyan, A.; Petrushina, I.; Kazarian, K.; Cribbs, D.H.; Petrovsky, N.; Agadjanyan, M.G.; et al. Alzheimer’s Disease Advax Cpg-Adjuvanted Multitep-Based Dual and Single Vaccines Induce High-Titer Antibodies against Various Forms of Tau and Aβ Pathological Molecules. Sci. Rep. 2016, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Simizu, B.; Abe, S.; Yamamoto, H.; Tano, Y.; Ota, Y.; Miyazawa, M.; Horie, H.; Satoh, K.; Wakabayashi, K. Development of inactivated poliovirus vaccine derived from Sabin strains. Biologicals 2006, 34, 151–154. [Google Scholar] [CrossRef]
- Kärber, G. Beitrag zur kollektiven Behandlung pharmakologischer Reihenversuche. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1931, 162, 480–483. [Google Scholar] [CrossRef]
- Romagnani, S. T-cell subsets (Th1 versus Th2). Ann. Allergy Asthma Immunol. 2000, 85, 9–21. [Google Scholar] [CrossRef]
- Singh, V.K.; Mehrotra, S.; Agarwal, S.S. The paradigm of Th1 and Th2 cytokines. Immunol. Res. 1999, 20, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Bode, C.; Zhao, G.; Steinhagen, F.; Kinjo, T.; Klinman, D.M. CpG DNA as a vaccine adjuvant. Expert Rev. Vaccines 2011, 10, 499–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietrich, J.; Andreasen, L.V.; Andersen, P.; Agger, E.M. Inducing Dose Sparing with Inactivated Polio Virus Formulated in Adjuvant CAF01. PLoS ONE 2014, 9, e100879. [Google Scholar] [CrossRef] [PubMed]
- Rockman, S.; Becher, D.; Dyson, A.; Koernig, S.; Morelli, A.B.; Barnden, M.; Camuglia, S.; Soupourmas, P.; Pearse, M.; Maraskovsky, E. Role of viral RNA and lipid in the adverse events associated with the 2010 Southern Hemisphere trivalent influenza vaccine. Vaccine 2014, 32, 3869–3876. [Google Scholar] [CrossRef] [Green Version]
- Lambert, S.B.; Chuk, L.R.; Nissen, M.D.; Nolan, T.M.; McVernon, J.; Booy, R.; Heron, L.; Richmond, P.C.; Walls, T.; Marshall, H.S.; et al. Safety and tolerability of a 2009 trivalent inactivated split-virion influenza vaccine in infants, children and adolescents. Influenza Other Respir. Viruses 2013, 7, 676–685. [Google Scholar] [CrossRef]
- Li, L.; Honda-Okubo, Y.; Li, C.; Sajkov, D.; Petrovsky, N. Delta Inulin Adjuvant Enhances Plasmablast Generation, Expression of Activation-Induced Cytidine Deaminase and B-Cell Affinity Maturation in Human Subjects Receiving Seasonal Influenza Vaccine. PLoS ONE 2015, 10, e0132003. [Google Scholar] [CrossRef]
- Gordon, D.; Kelley, P.; Heinzel, S.; Cooper, P.; Petrovsky, N. Immunogenicity and safety of Advax™, a novel polysaccharide adjuvant based on delta inulin, when formulated with hepatitis B surface antigen: A randomized controlled Phase 1 study. Vaccine 2014, 32, 6469–6477. [Google Scholar] [CrossRef] [Green Version]
- Heddle, R.; Smith, A.; Woodman, R.; Hissaria, P.; Petrovsky, N. Randomized controlled trial demonstrating the benefits of delta inulin adjuvanted immunotherapy in patients with bee venom allergy. J. Allergy Clin. Immunol. 2019, 144, 504–513.e16. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sanofi-Pasteur (IPOL) Polio Vaccine | |||||
| Study No. | Animals | Vaccine Antigen | Adjuvant | Dose Schedule | Purpose of Study |
| 1 | Female BALB/c | Trivalent inactivated polio (8 DU type 1, 1.6 DU type 2 and 6.4 DU type 3) | Alone or with Advax or Advax-CpG | 4 doses (weeks 0, 3, 7 and 12) | Long term immunogenicity/Neutralization Activity |
| Statens Serum Institut (SSI) Polio Vaccine | |||||
| Study No. | Animals | Vaccine Formulation | Adjuvant | Dose Schedule | Purpose of Study |
| 2 | Female BALB/c | Trivalent inactivated polio (10 DU type 1, 1 DU type 2 and 9 DU type 3) at 1, 1:10 and 1:100 dose | Alone or with Advax or Advax-CpG | 2-doses (3 weeks apart) | Immunogenicity/Neutralization Activity/Antigen dose sparing capacity |
| 3 | Female BALB/c | Trivalent inactivated polio and Monovalent at three different doses at 1, 1:10 and 1:100 dose | Alone or with Advax-CpG or CpG | 2-doses (3 weeks apart) | Antigen competition/Long-term Immunogencity |
| 4 | Female Wistar rats | Trivalent inactivated polio (10 DU type 1, 1 DU type 2 and 9 DU type 3) at 1, 1:10 and 1:100 dose | Alone or with Advax or Advax-CpG | 2-doses (3 weeks apart) | Species-specific differences Immunogenicity/Neutralization Activity/Weight loss |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Honda-Okubo, Y.; Baldwin, J.; Petrovsky, N. Advax-CpG Adjuvant Provides Antigen Dose-Sparing and Enhanced Immunogenicity for Inactivated Poliomyelitis Virus Vaccines. Pathogens 2021, 10, 500. https://doi.org/10.3390/pathogens10050500
Honda-Okubo Y, Baldwin J, Petrovsky N. Advax-CpG Adjuvant Provides Antigen Dose-Sparing and Enhanced Immunogenicity for Inactivated Poliomyelitis Virus Vaccines. Pathogens. 2021; 10(5):500. https://doi.org/10.3390/pathogens10050500
Chicago/Turabian StyleHonda-Okubo, Yoshikazu, Jeremy Baldwin, and Nikolai Petrovsky. 2021. "Advax-CpG Adjuvant Provides Antigen Dose-Sparing and Enhanced Immunogenicity for Inactivated Poliomyelitis Virus Vaccines" Pathogens 10, no. 5: 500. https://doi.org/10.3390/pathogens10050500