
Circulation of Babesia Species and Their Exposure to Humans through Ixodes ricinus


 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Babesia in Wildlife
2.1.1. Babesia Sensu Stricto (Clade X)
2.1.2. Babesia Clade I
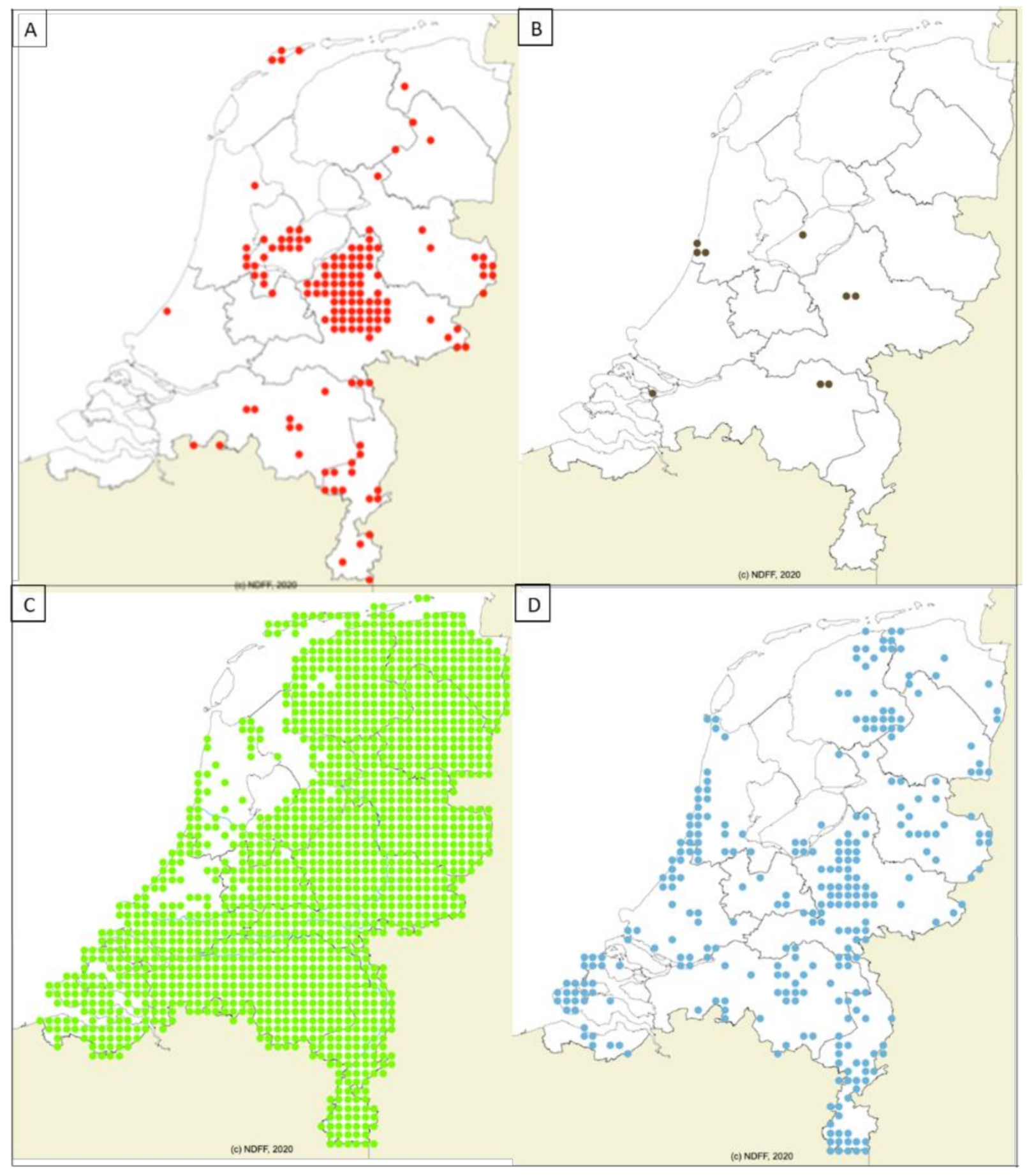
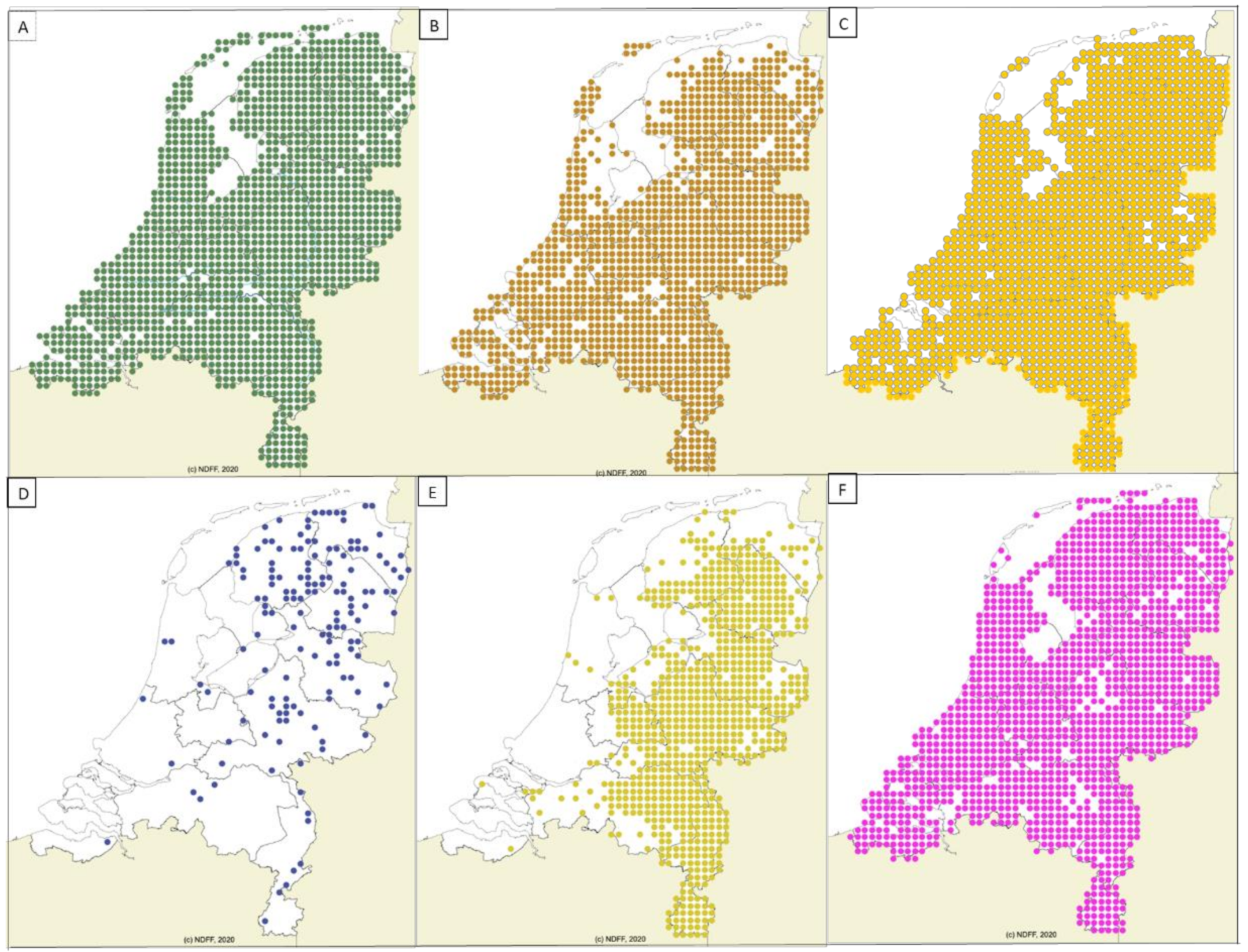
2.2. Geographic Distribution of Wildlife
2.3. Babesia Species in Ixodes ricinus
3. Discussion
4. Materials and Methods
4.1. Collection of Field Samples
4.2. DNA Extraction, qPCR, and RLB Protocols
4.3. DNA Sequencing
4.4. Species Identification Based on DNA Sequencing
4.5. Species Specific PCR
4.6. Data Collection on Geographic Spread of Vertebrates in The Netherlands
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Homer, M.J.; Aguilar-Delfin, I.; Telford, S.R., 3rd; Krause, P.J.; Persing, D.H. Babesiosis. Clin. Microbiol Rev. 2000, 13, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef] [PubMed]
- Jalovecka, M.; Sojka, D.; Ascencio, M.; Schnittger, L. Babesia Life Cycle—When Phylogeny Meets Biology. Trends Parasitol. 2019, 35, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Schreeg, M.E.; Marr, H.S.; Tarigo, J.L.; Cohn, L.A.; Bird, D.M.; Scholl, E.H.; Levy, M.G.; Wiegmann, B.M.; Birkenheuer, A.J. Mitochondrial Genome Sequences and Structures Aid in the Resolution of Piroplasmida phylogeny. PLoS ONE 2016, 11, e0165702. [Google Scholar] [CrossRef]
- Baneth, G.; Florin-Christensen, M.; Cardoso, L.; Schnittger, L. Reclassification of Theileria annae as Babesia vulpes sp. nov. Parasit. Vectors 2015, 8, 207. [Google Scholar] [CrossRef] [Green Version]
- Hornok, S.; Horvath, G.; Takacs, N.; Kontschan, J.; Szoke, K.; Farkas, R. Molecular identification of badger-associated Babesia sp. DNA in dogs: Updated phylogeny of piroplasms infecting Caniformia. Parasit. Vectors 2018, 11, 235. [Google Scholar] [CrossRef]
- Hrazdilova, K.; Rybarova, M.; Siroky, P.; Votypka, J.; Zintl, A.; Burgess, H.; Steinbauer, V.; Zakovcik, V.; Modry, D. Diversity of Babesia spp. in cervid ungulates based on the 18S rDNA and cytochrome c oxidase subunit I phylogenies. Infect. Genet. Evol. 2020, 77, 104060. [Google Scholar] [CrossRef]
- Nathaly Wieser, S.; Schnittger, L.; Florin-Christensen, M.; Delbecq, S.; Schetters, T. Vaccination against babesiosis using recombinant GPI-anchored proteins. Int. J. Parasitol. 2019, 49, 175–181. [Google Scholar] [CrossRef]
- Solano-Gallego, L.; Sainz, A.; Roura, X.; Estrada-Pena, A.; Miro, G. A review of canine babesiosis: The European perspective. Parasit. Vectors 2016, 9, 336. [Google Scholar] [CrossRef] [Green Version]
- Vannier, E.; Krause, P.J. Human babesiosis. N. Engl. J. Med. 2012, 366, 2397–2407. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Christie, J.; Koster, L.; Du, A.; Yao, C. Emerging Human Babesiosis with “Ground Zero” in North America. Microorganisms 2021, 9, 440. [Google Scholar] [CrossRef]
- Gray, J.S. Identity of the causal agents of human babesiosis in Europe. Int. J. Med. Microbiol 2006, 296 (Suppl. 40), 131–136. [Google Scholar] [CrossRef]
- Gray, J.; Zintl, A.; Hildebrandt, A.; Hunfeld, K.P.; Weiss, L. Zoonotic babesiosis: Overview of the disease and novel aspects of pathogen identity. Ticks Tick Borne Dis. 2010, 1, 3–10. [Google Scholar] [CrossRef]
- Hamsikova, Z.; Kazimirova, M.; Harustiakova, D.; Mahrikova, L.; Slovak, M.; Berthova, L.; Kocianova, E.; Schnittger, L. Babesia spp. in ticks and wildlife in different habitat types of Slovakia. Parasit. Vectors 2016, 9, 292. [Google Scholar] [CrossRef] [Green Version]
- Goethert, H.K.; Telford, S.R., 3rd. What is Babesia microti? Parasitology 2003, 127, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Wielinga, P.R.; Fonville, M.; Sprong, H.; Gaasenbeek, C.; Borgsteede, F.; van der Giessen, J.W. Persistent detection of Babesia EU1 and Babesia microti in Ixodes ricinus in the Netherlands during a 5-year surveillance: 2003–2007. Vector Borne Zoonotic Dis. 2009, 9, 119–122. [Google Scholar] [CrossRef]
- Bos, J.H.; Klip, F.C.; Sprong, H.; Broens, E.M.; Kik, M.J.L. Clinical outbreak of babesiosis caused by Babesia capreoli in captive reindeer (Rangifer tarandus tarandus) in the Netherlands. Ticks Tick Borne Dis. 2017, 8, 799–801. [Google Scholar] [CrossRef]
- Uilenberg, G.; Rombach, M.C.; Perie, N.M.; Zwart, D. Blood parasites of sheep in the Netherlands. II. Babesia motasi (Sporozoa, Babesiidae). Vet. Q 1980, 2, 3–14. [Google Scholar] [CrossRef]
- Uilenberg, G.; Top, P.D.; Arends, P.J.; Kool, P.J.; van Dijk, J.E.; van Schieveen, P.B.; Zwart, D. [Autochthonous babesiosis in dogs in the Netherlands?]. Tijdschr Diergeneeskd 1985, 110, 93–98. [Google Scholar]
- Matjila, T.P.; Nijhof, A.M.; Taoufik, A.; Houwers, D.; Teske, E.; Penzhorn, B.L.; de Lange, T.; Jongejan, F. Autochthonous canine babesiosis in the Netherlands. Vet. Parasitol. 2005, 131, 23–29. [Google Scholar] [CrossRef]
- Jongejan, F.; Ringenier, M.; Putting, M.; Berger, L.; Burgers, S.; Kortekaas, R.; Lenssen, J.; van Roessel, M.; Wijnveld, M.; Madder, M. Novel foci of Dermacentor reticulatus ticks infected with Babesia canis and Babesia caballi in the Netherlands and in Belgium. Parasit. Vectors 2015, 8, 232. [Google Scholar] [CrossRef] [Green Version]
- Carpi, G.; Walter, K.S.; Mamoun, C.B.; Krause, P.J.; Kitchen, A.; Lepore, T.J.; Dwivedi, A.; Cornillot, E.; Caccone, A.; Diuk-Wasser, M.A. Babesia microti from humans and ticks hold a genomic signature of strong population structure in the United States. BMC Genom. 2016, 17, 888. [Google Scholar] [CrossRef] [Green Version]
- Krause, P.J. Human babesiosis. Int. J. Parasitol. 2019, 49, 165–174. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Gray, J.S.; Hunfeld, K.P. Human babesiosis in Europe: What clinicians need to know. Infection 2013, 41, 1057–1072. [Google Scholar] [CrossRef]
- Gonzalez, L.M.; Castro, E.; Lobo, C.A.; Richart, A.; Ramiro, R.; Gonzalez-Camacho, F.; Luque, D.; Velasco, A.C.; Montero, E. First report of Babesia divergens infection in an HIV patient. Int. J. Infect. Dis. 2015, 33, 202–204. [Google Scholar] [CrossRef] [Green Version]
- Morch, K.; Holmaas, G.; Frolander, P.S.; Kristoffersen, E.K. Severe human Babesia divergens infection in Norway. Int. J. Infect. Dis. 2015, 33, 37–38. [Google Scholar] [CrossRef] [Green Version]
- Paleau, A.; Candolfi, E.; Souply, L.; De Briel, D.; Delarbre, J.M.; Lipsker, D.; Jouglin, M.; Malandrin, L.; Hansmann, Y.; Martinot, M. Human babesiosis in Alsace. Med. Mal. Infect. 2020, 50, 486–491. [Google Scholar] [CrossRef]
- Hildebrandt, A.; Hunfeld, K.P.; Baier, M.; Krumbholz, A.; Sachse, S.; Lorenzen, T.; Kiehntopf, M.; Fricke, H.J.; Straube, E. First confirmed autochthonous case of human Babesia microti infection in Europe. Eur. J. Clin. Microbiol. Infect. Dis 2007, 26, 595–601. [Google Scholar] [CrossRef]
- Arsuaga, M.; Gonzalez, L.M.; Lobo, C.A.; de la Calle, F.; Bautista, J.M.; Azcarate, I.G.; Puente, S.; Montero, E. First Report of Babesia microti-Caused Babesiosis in Spain. Vector Borne Zoonotic Dis. 2016, 16, 677–679. [Google Scholar] [CrossRef] [Green Version]
- Haselbarth, K.; Tenter, A.M.; Brade, V.; Krieger, G.; Hunfeld, K.P. First case of human babesiosis in Germany—Clinical presentation and molecular characterisation of the pathogen. Int. J. Med. Microbiol. 2007, 297, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Gorenflot, A.; Moubri, K.; Precigout, E.; Carcy, B.; Schetters, T.P. Human babesiosis. Ann. Trop. Med. Parasitol. 1998, 92, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Foppa, I.M.; Krause, P.J.; Spielman, A.; Goethert, H.; Gern, L.; Brand, B.; Telford, S.R., 3rd. Entomologic and serologic evidence of zoonotic transmission of Babesia microti, eastern Switzerland. Emerg. Infect. Dis. 2002, 8, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Hunfeld, K.P.; Lambert, A.; Kampen, H.; Albert, S.; Epe, C.; Brade, V.; Tenter, A.M. Seroprevalence of Babesia infections in humans exposed to ticks in midwestern Germany. J. Clin. Microbiol. 2002, 40, 2431–2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrielli, S.; Calderini, P.; Cassini, R.; Galuppi, R.; Tampieri, M.P.; Pietrobelli, M.; Cancrini, G. Human exposure to piroplasms in Central and Northern Italy. Vet. Ital. 2014, 50, 41–47. [Google Scholar] [CrossRef]
- Lempereur, L.; Shiels, B.; Heyman, P.; Moreau, E.; Saegerman, C.; Losson, B.; Malandrin, L. A retrospective serological survey on human babesiosis in Belgium. Clin. Microbiol. Infect. 2015, 21, 96.e1–96.e7. [Google Scholar] [CrossRef] [Green Version]
- Zukiewicz-Sobczak, W.; Zwolinski, J.; Chmielewska-Badora, J.; Galinska, E.M.; Cholewa, G.; Krasowska, E.; Zagorski, J.; Wojtyla, A.; Tomasiewicz, K.; Klapec, T. Prevalence of antibodies against selected zoonotic agents in forestry workers from eastern and southern Poland. Ann. Agric. Environ. Med. 2014, 21, 767–770. [Google Scholar] [CrossRef] [Green Version]
- Zintl, A.; Mulcahy, G.; Skerrett, H.E.; Taylor, S.M.; Gray, J.S. Babesia divergens, a bovine blood parasite of veterinary and zoonotic importance. Clin. Microbiol. Rev. 2003, 16, 622–636. [Google Scholar] [CrossRef] [Green Version]
- Tijsse-Klasen, E.; Koopmans, M.P.; Sprong, H. Tick-borne pathogen—reversed and conventional discovery of disease. Front. Public Health 2014, 2, 73. [Google Scholar] [CrossRef] [Green Version]
- Jahfari, S.; Hofhuis, A.; Fonville, M.; van der Giessen, J.; van Pelt, W.; Sprong, H. Molecular Detection of Tick-Borne Pathogens in Humans with Tick Bites and Erythema Migrans, in the Netherlands. PLoS Negl. Trop. Dis. 2016, 10, e0005042. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.S.; Estrada-Pena, A.; Zintl, A. Vectors of Babesiosis. Annu. Rev. Entomol. 2019, 64, 149–165. [Google Scholar] [CrossRef]
- Friedhoff, K.T. Transmission of Babesia. In Babesiosis of Domestic Animals and Man; CRC Press: Boca Raton, FL, USA, 1988; pp. 23–52. [Google Scholar]
- Sprong, H.; Azagi, T.; Hoornstra, D.; Nijhof, A.M.; Knorr, S.; Baarsma, M.E.; Hovius, J.W. Control of Lyme borreliosis and other Ixodes ricinus-borne diseases. Parasit. Vectors 2018, 11, 145. [Google Scholar] [CrossRef] [Green Version]
- Mehlhorn, H.; Shein, E. The piroplasms: Life cycle and sexual stages. Adv. Parasitol. 1984, 23, 37–103. [Google Scholar] [CrossRef]
- Coipan, E.C.; Jahfari, S.; Fonville, M.; Maassen, C.B.; van der Giessen, J.; Takken, W.; Takumi, K.; Sprong, H. Spatiotemporal dynamics of emerging pathogens in questing Ixodes ricinus. Front. Cell Infect. Microbiol. 2013, 3, 36. [Google Scholar] [CrossRef] [Green Version]
- Takumi, K.; Sprong, H.; Hofmeester, T.R. Impact of vertebrate communities on Ixodes ricinus-borne disease risk in forest areas. Parasit. Vectors 2019, 12, 434. [Google Scholar] [CrossRef] [Green Version]
- Hofmeester, T.; Coipan, E.; Van Wieren, S.; Prins, H.; Takken, W.; Sprong, H. Few vertebrate species dominate the Borrelia burgdorferi sl life cycle. Environ. Res. Lett. 2016, 11, 043001. [Google Scholar] [CrossRef] [Green Version]
- Mihalca, A.D.; Sandor, A.D. The role of rodents in the ecology of Ixodes ricinus and associated pathogens in Central and Eastern Europe. Front. Cell Infect. Microbiol. 2013, 3, 56. [Google Scholar] [CrossRef] [Green Version]
- Martin, L.B.; Burgan, S.C.; Adelman, J.S.; Gervasi, S.S. Host Competence: An Organismal Trait to Integrate Immunology and Epidemiology. Integr. Comp. Biol. 2016, 56, 1225–1237. [Google Scholar] [CrossRef] [Green Version]
- Keesing, F.; Brunner, J.; Duerr, S.; Killilea, M.; Logiudice, K.; Schmidt, K.; Vuong, H.; Ostfeld, R.S. Hosts as ecological traps for the vector of Lyme disease. Proc. Biol. Sci. 2009, 276, 3911–3919. [Google Scholar] [CrossRef] [Green Version]
- LoGiudice, K.; Ostfeld, R.S.; Schmidt, K.A.; Keesing, F. The ecology of infectious disease: Effects of host diversity and community composition on Lyme disease risk. Proc. Natl. Acad. Sci. USA 2003, 100, 567–571. [Google Scholar] [CrossRef] [Green Version]
- Malandrin, L.; Jouglin, M.; Sun, Y.; Brisseau, N.; Chauvin, A. Redescription of Babesia capreoli (Enigk and Friedhoff, 1962) from roe deer (Capreolus capreolus): Isolation, cultivation, host specificity, molecular characterisation and differentiation from Babesia divergens. Int. J. Parasitol. 2010, 40, 277–284. [Google Scholar] [CrossRef]
- Overzier, E.; Pfister, K.; Herb, I.; Mahling, M.; Bock, G., Jr.; Silaghi, C. Detection of tick-borne pathogens in roe deer (Capreolus capreolus), in questing ticks (Ixodes ricinus), and in ticks infesting roe deer in southern Germany. Ticks Tick Borne Dis. 2013, 4, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Michel, A.O.; Mathis, A.; Ryser-Degiorgis, M.P. Babesia spp. in European wild ruminant species: Parasite diversity and risk factors for infection. Vet. Res. 2014, 45, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bown, K.J.; Lambin, X.; Telford, G.R.; Ogden, N.H.; Telfer, S.; Woldehiwet, Z.; Birtles, R.J. Relative importance of Ixodes ricinus and Ixodes trianguliceps as vectors for Anaplasma phagocytophilum and Babesia microti in field vole (Microtus agrestis) populations. Appl. Environ. Microbiol. 2008, 74, 7118–7125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cayol, C.; Jaaskelainen, A.; Koskela, E.; Kyrolainen, S.; Mappes, T.; Siukkola, A.; Kallio, E.R. Sympatric Ixodes-tick species: Pattern of distribution and pathogen transmission within wild rodent populations. Sci. Rep. 2018, 8, 16660. [Google Scholar] [CrossRef]
- Najm, N.A.; Meyer-Kayser, E.; Hoffmann, L.; Herb, I.; Fensterer, V.; Pfister, K.; Silaghi, C. A molecular survey of Babesia spp. and Theileria spp. in red foxes (Vulpes vulpes) and their ticks from Thuringia, Germany. Ticks Tick Borne Dis. 2014, 5, 386–391. [Google Scholar] [CrossRef]
- Dudek, K. Impact of biodiversity on tick-borne diseases. Przegl Epidemiol. 2014, 68, 681–684. [Google Scholar]
- Tomassone, L.; Berriatua, E.; De Sousa, R.; Duscher, G.G.; Mihalca, A.D.; Silaghi, C.; Sprong, H.; Zintl, A. Neglected vector-borne zoonoses in Europe: Into the wild. Vet. Parasitol. 2018, 251, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Lambin, E.F.; Tran, A.; Vanwambeke, S.O.; Linard, C.; Soti, V. Pathogenic landscapes: Interactions between land, people, disease vectors, and their animal hosts. Int. J. Health Geogr. 2010, 9, 54. [Google Scholar] [CrossRef] [Green Version]
- Tijsse-Klasen, E.; Fonville, M.; Reimerink, J.H.; Spitzen-van der Sluijs, A.; Sprong, H. Role of sand lizards in the ecology of Lyme and other tick-borne diseases in the Netherlands. Parasit. Vectors 2010, 3, 42. [Google Scholar] [CrossRef] [Green Version]
- Krawczyk, A.I.; van Duijvendijk, G.L.A.; Swart, A.; Heylen, D.; Jaarsma, R.I.; Jacobs, F.H.H.; Fonville, M.; Sprong, H.; Takken, W. Effect of rodent density on tick and tick-borne pathogen populations: Consequences for infectious disease risk. Parasit. Vectors 2020, 13, 34. [Google Scholar] [CrossRef] [Green Version]
- Heylen, D.; Lasters, R.; Adriaensen, F.; Fonville, M.; Sprong, H.; Matthysen, E. Ticks and tick-borne diseases in the city: Role of landscape connectivity and green space characteristics in a metropolitan area. Sci. Total Environ. 2019, 670, 941–949. [Google Scholar] [CrossRef]
- Sprong, H.; Moonen, S.; van Wieren, S.E.; Hofmeester, T.R. Effects of cattle grazing on Ixodes ricinus-borne disease risk in forest areas of the Netherlands. Ticks Tick Borne Dis. 2020, 11, 101355. [Google Scholar] [CrossRef]
- Bonnet, S.; Jouglin, M.; L’Hostis, M.; Chauvin, A. Babesia sp. EU1 from roe deer and transmission within Ixodes ricinus. Emerg. Infect. Dis. 2007, 13, 1208–1210. [Google Scholar] [CrossRef]
- Kauffmann, M.; Rehbein, S.; Hamel, D.; Lutz, W.; Heddergott, M.; Pfister, K.; Silaghi, C. Anaplasma phagocytophilum and Babesia spp. in roe deer (Capreolus capreolus), fallow deer (Dama dama) and mouflon (Ovis musimon) in Germany. Mol. Cell Probes 2017, 31, 46–54. [Google Scholar] [CrossRef]
- Zanet, S.; Trisciuoglio, A.; Bottero, E.; de Mera, I.G.; Gortazar, C.; Carpignano, M.G.; Ferroglio, E. Piroplasmosis in wildlife: Babesia and Theileria affecting free-ranging ungulates and carnivores in the Italian Alps. Parasit. Vectors 2014, 7, 70. [Google Scholar] [CrossRef] [Green Version]
- Welc-Faleciak, R.; Werszko, J.; Cydzik, K.; Bajer, A.; Michalik, J.; Behnke, J.M. Co-infection and genetic diversity of tick-borne pathogens in roe deer from Poland. Vector Borne Zoonotic Dis. 2013, 13, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Duh, D.; Petrovec, M.; Bidovec, A.; Avsic-Zupanc, T. Cervids as Babesiae hosts, Slovenia. Emerg. Infect. Dis. 2005, 11, 1121–1123. [Google Scholar] [CrossRef]
- Garcia-Sanmartin, J.; Aurtenetxe, O.; Barral, M.; Marco, I.; Lavin, S.; Garcia-Perez, A.L.; Hurtado, A. Molecular detection and characterization of piroplasms infecting cervids and chamois in Northern Spain. Parasitology 2007, 134, 391–398. [Google Scholar] [CrossRef]
- Cezanne, R.; Mrowietz, N.; Eigner, B.; Duscher, G.G.; Glawischnig, W.; Fuehrer, H.P. Molecular analysis of Anaplasma phagocytophilum and Babesia divergens in red deer (Cervus elaphus) in Western Austria. Mol. Cell Probes 2017, 31, 55–58. [Google Scholar] [CrossRef]
- Sawczuk, M.; Maciejewska, A.; Adamska, M.; Skotarczak, B. [Roe deer (Capreolus capreolus) and red deer (Cervus elaphus) as a reservoir of protozoans from Babesia and Theileria genus in north-western Poland]. Wiad Parazytol. 2005, 51, 243–247. [Google Scholar]
- Karbowiak, G.; Demiaszkiewicz, A.W.; Pyziel, A.M.; Wita, I.; Moskwa, B.; Werszko, J.; Bien, J.; Gozdzik, K.; Lachowicz, J.; Cabaj, W. The parasitic fauna of the European bison (Bison bonasus) (Linnaeus, 1758) and their impact on the conservation. Part 1. The summarising list of parasites noted. Acta Parasitol. 2014, 59, 363–371. [Google Scholar] [CrossRef]
- Razanske, I.; Rosef, O.; Radzijevskaja, J.; Bratchikov, M.; Griciuviene, L.; Paulauskas, A. Prevalence and co-infection with tick-borne Anaplasma phagocytophilum and Babesia spp. in red deer (Cervus elaphus) and roe deer (Capreolus capreolus) in Southern Norway. Int. J. Parasitol. Parasites Wildl. 2019, 8, 127–134. [Google Scholar] [CrossRef]
- Silaghi, C.; Hamel, D.; Pfister, K.; Rehbein, S. Babesia species and co-infection with Anaplasma phagocytophilum in free-ranging ungulates from Tyrol (Austria). Tierärztliche Mschr. Vet. Med. Austria 2011, 98, 268–274. [Google Scholar]
- Michelet, L.; Delannoy, S.; Devillers, E.; Umhang, G.; Aspan, A.; Juremalm, M.; Chirico, J.; van der Wal, F.J.; Sprong, H.; Boye Pihl, T.P.; et al. High-throughput screening of tick-borne pathogens in Europe. Front. Cell Infect. Microbiol. 2014, 4, 103. [Google Scholar] [CrossRef] [PubMed]
- Lempereur, L.; Wirtgen, M.; Nahayo, A.; Caron, Y.; Shiels, B.; Saegerman, C.; Losson, B.; Linden, A. Wild cervids are host for tick vectors of babesia species with zoonotic capability in Belgium. Vector Borne Zoonotic Dis. 2012, 12, 275–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schorn, S.; Pfister, K.; Reulen, H.; Mahling, M.; Silaghi, C. Occurrence of Babesia spp., Rickettsia spp. and Bartonella spp. in Ixodes ricinus in Bavarian public parks, Germany. Parasit. Vectors 2011, 4, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamska, M.; Skotarczak, B. Molecular detecting of piroplasms in feeding and questing Ixodes ricinus ticks. Ann. Parasitol. 2017, 63, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, M.E.; Andersson, M.O. Babesia species in questing Ixodes ricinus, Sweden. Ticks Tick Borne Dis. 2016, 7, 10–12. [Google Scholar] [CrossRef]
- Schotta, A.M.; Wijnveld, M.; Stockinger, H.; Stanek, G. Approaches for Reverse Line Blot-Based Detection of Microbial Pathogens in Ixodes ricinus Ticks Collected in Austria and Impact of the Chosen Method. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Egyed, L.; Elo, P.; Sreter-Lancz, Z.; Szell, Z.; Balogh, Z.; Sreter, T. Seasonal activity and tick-borne pathogen infection rates of Ixodes ricinus ticks in Hungary. Ticks Tick Borne Dis. 2012, 3, 90–94. [Google Scholar] [CrossRef]
- Joyner, L.P.; Davies, S.F.; Kendall, S.B. The experimental transmission of Babesia divergens by Ixodes Ricinus. Exp. Parasitol. 1963, 14, 367–373. [Google Scholar] [CrossRef]
- Bonnet, S.; Jouglin, M.; Malandrin, L.; Becker, C.; Agoulon, A.; L’Hostis, M.; Chauvin, A. Transstadial and transovarial persistence of Babesia divergens DNA in Ixodes ricinus ticks fed on infected blood in a new skin-feeding technique. Parasitology 2007, 134, 197–207. [Google Scholar] [CrossRef]
- Radzijevskaja, J.; Paulauskas, A.; Rosef, O. Prevalence of Anaplasma phagocytophilum and Babesia divergens in Ixodes ricinus ticks from Lithuania and Norway. Int. J. Med Microbiol. 2008, 298, 218–221. [Google Scholar] [CrossRef] [Green Version]
- Donnelly, J.; Peirce, M.A. Experiments on the transmission of Babesia divergens to cattle by the tick Ixodes ricinus. Int. J. Parasitol. 1975, 5, 363–367. [Google Scholar] [CrossRef]
- Mackenstedt, U.; Gauer, M.; Mehlhorn, H.; Schein, E.; Hauschild, S. Sexual cycle of Babesia divergens confirmed by DNA measurements. Parasitol. Res. 1990, 76, 199–206. [Google Scholar] [CrossRef]
- Andersson, M.O.; Vichova, B.; Tolf, C.; Krzyzanowska, S.; Waldenstrom, J.; Karlsson, M.E. Co-infection with Babesia divergens and Anaplasma phagocytophilum in cattle (Bos taurus), Sweden. Ticks Tick Borne Dis. 2017, 8, 933–935. [Google Scholar] [CrossRef]
- Lommano, E.; Bertaiola, L.; Dupasquier, C.; Gern, L. Infections and coinfections of questing Ixodes ricinus ticks by emerging zoonotic pathogens in Western Switzerland. Appl. Environ. Microbiol. 2012, 78, 4606–4612. [Google Scholar] [CrossRef] [Green Version]
- Meer-Scherrer, L.; Adelson, M.; Mordechai, E.; Lottaz, B.; Tilton, R. Babesia microti infection in Europe. Curr. Microbiol. 2004, 48, 435–437. [Google Scholar] [CrossRef]
- Hvidsten, D.; Frafjord, K.; Gray, J.S.; Henningsson, A.J.; Jenkins, A.; Kristiansen, B.E.; Lager, M.; Rognerud, B.; Slatsve, A.M.; Stordal, F.; et al. The distribution limit of the common tick, Ixodes ricinus, and some associated pathogens in north-western Europe. Ticks Tick Borne Dis. 2020, 11, 101388. [Google Scholar] [CrossRef]
- Gray, J.S.; Dautel, H.; Estrada-Pena, A.; Kahl, O.; Lindgren, E. Effects of climate change on ticks and tick-borne diseases in europe. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 593232. [Google Scholar] [CrossRef]
- Burbaitė, L.; Csányi, S. Red deer population and harvest changes in Europe. Acta Zool. Litu. 2010, 20, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Checa, R.; Lopez-Beceiro, A.M.; Montoya, A.; Barrera, J.P.; Ortega, N.; Galvez, R.; Marino, V.; Gonzalez, J.; Olmeda, A.S.; Fidalgo, L.E.; et al. Babesia microti-like piroplasm (syn. Babesia vulpes) infection in red foxes (Vulpes vulpes) in NW Spain (Galicia) and its relationship with Ixodes hexagonus. Vet. Parasitol. 2018, 252, 22–28. [Google Scholar] [CrossRef]
- Falkeno, U.; Tasker, S.; Osterman-Lind, E.; Tvedten, H.W. Theileria annae in a young Swedish dog. Acta Vet. Scand. 2013, 55, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rene-Martellet, M.; Moro, C.V.; Chene, J.; Bourdoiseau, G.; Chabanne, L.; Mavingui, P. Update on epidemiology of canine babesiosis in Southern France. BMC Vet. Res. 2015, 11, 223. [Google Scholar] [CrossRef] [PubMed]
- Bartley, P.M.; Wilson, C.; Innes, E.A.; Katzer, F. Detection of Babesia DNA in blood and spleen samples from Eurasian badgers (Meles meles) in Scotland. Parasitology 2017, 144, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Trauttwein, K.; Takacs, N.; Hodzic, A.; Duscher, G.G.; Kontschan, J. Molecular analysis of Ixodes rugicollis, Candidatus Neoehrlichia sp. (FU98) and a novel Babesia genotype from a European badger (Meles meles). Ticks Tick Borne Dis. 2017, 8, 41–44. [Google Scholar] [CrossRef]
- Beck, R.; Vojta, L.; Curkovic, S.; Mrljak, V.; Margaletic, J.; Habrun, B. Molecular survey of Babesia microti in wild rodents in central Croatia. Vector Borne Zoonotic Dis. 2011, 11, 81–83. [Google Scholar] [CrossRef]
- Obiegala, A.; Pfeffer, M.; Pfister, K.; Karnath, C.; Silaghi, C. Molecular examinations of Babesia microti in rodents and rodent-attached ticks from urban and sylvatic habitats in Germany. Ticks Tick Borne Dis. 2015, 6, 445–449. [Google Scholar] [CrossRef]
- Turner, C.M. Seasonal and age distributions of Babesia, Hepatozoon, Trypanosoma and Grahamella species in Clethrionomys glareolus and Apodemus sylvaticus populations. Parasitology 1986, 93 (Pt. 2), 279–289. [Google Scholar] [CrossRef]
- Duh, D.; Petrovec, M.; Trilar, T.; Avsic-Zupanc, T. The molecular evidence of Babesia microti infection in small mammals collected in Slovenia. Parasitology 2003, 126, 113–117. [Google Scholar] [CrossRef]
- Kallio, E.R.; Begon, M.; Birtles, R.J.; Bown, K.J.; Koskela, E.; Mappes, T.; Watts, P.C. First report of Anaplasma phagocytophilum and Babesia microti in rodents in Finland. Vector Borne Zoonotic Dis. 2014, 14, 389–393. [Google Scholar] [CrossRef] [Green Version]
- Hartelt, K.; Oehme, R.; Frank, H.; Brockmann, S.O.; Hassler, D.; Kimmig, P. Pathogens and symbionts in ticks: Prevalence of Anaplasma phagocytophilum (Ehrlichia sp.), Wolbachia sp., Rickettsia sp., and Babesia sp. in Southern Germany. Int. J. Med. Microbiol. 2004, 293 (Suppl. 37), 86–92. [Google Scholar] [CrossRef]
- Asman, M.; Solarz, K.; Cuber, P.; Gasior, T.; Szilman, P.; Szilman, E.; Tondas, E.; Matzullok, A.; Kusion, N.; Florek, K. Detection of protozoans Babesia microti and Toxoplasma gondii and their co-existence in ticks (Acari: Ixodida) collected in Tarnogorski district (Upper Silesia, Poland). Ann. Agric. Environ. Med. 2015, 22, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Lempereur, L.; De Cat, A.; Caron, Y.; Madder, M.; Claerebout, E.; Saegerman, C.; Losson, B. First molecular evidence of potentially zoonotic Babesia microti and Babesia sp. EU1 in Ixodes ricinus ticks in Belgium. Vector Borne Zoonotic Dis. 2011, 11, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Karbowiak, G.; Biernat, B.; Stanczak, J.; Werszko, J.; Szewczyk, T.; Sytykiewicz, H. The role of particular ticks developmental stages in the circulation of tick-borne pathogens in Central Europe. 6. Babesia. Ann. Parasitol. 2018, 64, 265–284. [Google Scholar] [CrossRef]
- Topolovec, J.; Puntaric, D.; Antolovic-Pozgain, A.; Vukovic, D.; Topolovec, Z.; Milas, J.; Drusko-Barisic, V.; Venus, M. Serologically detected "new" tick-borne zoonoses in eastern Croatia. Croat. Med. J. 2003, 44, 626–629. [Google Scholar]
- Hunfeld, K.P.; Allwinn, R.; Peters, S.; Kraiczy, P.; Brade, V. Serologic evidence for tick-borne pathogens other than Borrelia burgdorferi (TOBB) in Lyme borreliosis patients from midwestern Germany. Wien. Klin Wochenschr. 1998, 110, 901–908. [Google Scholar]
- Belongia, E.A. Epidemiology and impact of coinfections acquired from Ixodes ticks. Vector Borne Zoonotic Dis. 2002, 2, 265–273. [Google Scholar] [CrossRef]
- Raileanu, C.; Moutailler, S.; Pavel, I.; Porea, D.; Mihalca, A.D.; Savuta, G.; Vayssier-Taussat, M. Borrelia Diversity and Co-infection with Other Tick Borne Pathogens in Ticks. Front. Cell Infect. Microbiol. 2017, 7, 36. [Google Scholar] [CrossRef] [Green Version]
- Moutailler, S.; Valiente Moro, C.; Vaumourin, E.; Michelet, L.; Tran, F.H.; Devillers, E.; Cosson, J.F.; Gasqui, P.; Van, V.T.; Mavingui, P.; et al. Co-infection of Ticks: The Rule Rather Than the Exception. PLoS Negl. Trop. Dis. 2016, 10, e0004539. [Google Scholar] [CrossRef] [Green Version]
- Wormser, G.P.; Dattwyler, R.J.; Shapiro, E.D.; Halperin, J.J.; Steere, A.C.; Klempner, M.S.; Krause, P.J.; Bakken, J.S.; Strle, F.; Stanek, G.; et al. The clinical assessment, treatment, and prevention of lyme disease, human granulocytic anaplasmosis, and babesiosis: Clinical practice guidelines by the Infectious Diseases Society of America. Clin. Infect. Dis. 2006, 43, 1089–1134. [Google Scholar] [CrossRef]
- Krause, P.J.; Telford, S.R., 3rd; Spielman, A.; Sikand, V.; Ryan, R.; Christianson, D.; Burke, G.; Brassard, P.; Pollack, R.; Peck, J.; et al. Concurrent Lyme disease and babesiosis. Evidence for increased severity and duration of illness. JAMA 1996, 275, 1657–1660. [Google Scholar] [CrossRef]
- Morner, T.; Obendorf, D.L.; Artois, M.; Woodford, M.H. Surveillance and monitoring of wildlife diseases. Rev. Sci. Tech. 2002, 21, 67–76. [Google Scholar] [CrossRef]
- Coipan, C.E.; van Duijvendijk, G.L.A.; Hofmeester, T.R.; Takumi, K.; Sprong, H. The genetic diversity of Borrelia afzelii is not maintained by the diversity of the rodent hosts. Parasit. Vectors 2018, 11, 454. [Google Scholar] [CrossRef] [Green Version]
- Hofmeester, T.R.; Sprong, H.; Jansen, P.A.; Prins, H.H.T.; van Wieren, S.E. Deer presence rather than abundance determines the population density of the sheep tick, Ixodes ricinus, in Dutch forests. Parasit. Vectors 2017, 10, 433. [Google Scholar] [CrossRef]
- Hofmeester, T.R.; Krawczyk, A.I.; van Leeuwen, A.D.; Fonville, M.; Montizaan, M.G.E.; van den Berge, K.; Gouwy, J.; Ruyts, S.C.; Verheyen, K.; Sprong, H. Role of mustelids in the life-cycle of ixodid ticks and transmission cycles of four tick-borne pathogens. Parasit. Vectors 2018, 11, 600. [Google Scholar] [CrossRef]
- Ruyts, S.C.; Frazer-Mendelewska, E.; Van Den Berge, K.; Verheyen, K.; Sprong, H. Molecular detection of tick-borne pathogens Borrelia afzelii, Borrelia miyamotoi and Anaplasma phagocytophilum in Eurasian red squirrels (Sciurus vulgaris). Eur. J. Wildl. Res. 2017, 63, 43. [Google Scholar] [CrossRef]
- Maas, M.; van Roon, A.; Dam-Deisz, W.; van der Giessen, J. Geringe Verspreiding Van Vossenlintworm in Groningen; RIVM Briefrapport: De Bilt, The Netherlands, 2018. [Google Scholar]
- Maas, M.; Mulder, J.; Montizaan, M.; Dam-Deisz, W.; Jaarsma, R.; Takumi, K.; van Roon, A.; Franssen, F.; van der Giessen, J. Zoönotische Pathogenen bij de Wasbeerhond en Wasbeer in Nederland; RIVM Briefrapport: De Bilt, The Netherlands, 2018; pp. 1–28. [Google Scholar]
- Guldemond, A.; Dijkman, W.; Keuper, D. Wilde Zwijnen op Weg in Nederland; CLM Onderzoek en Advies: Culemborg, The Netherlands, 2015; pp. 1–48. [Google Scholar]
- Rijks, J.M.; Montizaan, M.G.E.; Bakker, N.; de Vries, A.; Van Gucht, S.; Swaan, C.; van den Broek, J.; Grone, A.; Sprong, H. Tick-Borne Encephalitis Virus Antibodies in Roe Deer, the Netherlands. Emerg. Infect. Dis. 2019, 25, 342–345. [Google Scholar] [CrossRef]
- Estrada-Pena, A.; D’Amico, G.; Palomar, A.M.; Dupraz, M.; Fonville, M.; Heylen, D.; Habela, M.A.; Hornok, S.; Lempereur, L.; Madder, M.; et al. A comparative test of ixodid tick identification by a network of European researchers. Ticks Tick Borne Dis. 2017, 8, 540–546. [Google Scholar] [CrossRef] [Green Version]
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Ticks of Europe and North Africa: A Guide to Species Identification; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Nagore, D.; Garcia-Sanmartin, J.; Garcia-Perez, A.L.; Juste, R.A.; Hurtado, A. Detection and identification of equine Theileria and Babesia species by reverse line blotting: Epidemiological survey and phylogenetic analysis. Vet. Parasitol. 2004, 123, 41–54. [Google Scholar] [CrossRef]
- Schnittger, L.; Yin, H.; Qi, B.; Gubbels, M.J.; Beyer, D.; Niemann, S.; Jongejan, F.; Ahmed, J.S. Simultaneous detection and differentiation of Theileria and Babesia parasites infecting small ruminants by reverse line blotting. Parasitol. Res. 2004, 92, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Altay, K.; Dumanli, N.; Aktas, M. Molecular identification, genetic diversity and distribution of Theileria and Babesia species infecting small ruminants. Vet. Parasitol. 2007, 147, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Schouls, L.M.; Van De Pol, I.; Rijpkema, S.G.; Schot, C.S. Detection and identification of Ehrlichia, Borrelia burgdorferi sensu lato, and Bartonella species in Dutch Ixodes ricinus ticks. J. Clin. Microbiol. 1999, 37, 2215–2222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oines, O.; Radzijevskaja, J.; Paulauskas, A.; Rosef, O. Prevalence and diversity of Babesia spp. in questing Ixodes ricinus ticks from Norway. Parasit. Vectors 2012, 5, 156. [Google Scholar] [CrossRef] [Green Version]
- Kazimirova, M.; Hamsikova, Z.; Spitalska, E.; Minichova, L.; Mahrikova, L.; Caban, R.; Sprong, H.; Fonville, M.; Schnittger, L.; Kocianova, E. Diverse tick-borne microorganisms identified in free-living ungulates in Slovakia. Parasit. Vectors 2018, 11, 495. [Google Scholar] [CrossRef]
- Hrazdilova, K.; Mysliwy, I.; Hildebrand, J.; Bunkowska-Gawlik, K.; Janaczyk, B.; Perec-Matysiak, A.; Modry, D. Paralogs vs. genotypes? Variability of Babesia canis assessed by 18S rDNA and two mitochondrial markers. Vet. Parasitol. 2019, 266, 103–110. [Google Scholar] [CrossRef]
- Gou, H.; Guan, G.; Liu, A.; Ma, M.; Xu, Z.; Liu, Z.; Ren, Q.; Li, Y.; Yang, J.; Chen, Z.; et al. A DNA barcode for Piroplasmea. Acta Trop. 2012, 124, 92–97. [Google Scholar] [CrossRef]
- Pan, W.; Byrne-Steele, M.; Wang, C.; Lu, S.; Clemmons, S.; Zahorchak, R.J.; Han, J. DNA polymerase preference determines PCR priming efficiency. BMC Biotechnol. 2014, 14, 10. [Google Scholar] [CrossRef] [Green Version]
- Tuvshintulga, B.; Sivakumar, T.; Battsetseg, B.; Narantsatsaral, S.O.; Enkhtaivan, B.; Battur, B.; Hayashida, K.; Okubo, K.; Ishizaki, T.; Inoue, N.; et al. The PCR detection and phylogenetic characterization of Babesia microti in questing ticks in Mongolia. Parasitol. Int. 2015, 64, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Coimbra-Dores, M.J.; Jaarsma, R.I.; Carmo, A.O.; Maia-Silva, M.; Fonville, M.; da Costa, D.F.F.; Brandao, R.M.L.; Azevedo, F.; Casero, M.; Oliveira, A.C.; et al. Mitochondrial sequences of Rhipicephalus and Coxiella endosymbiont reveal evidence of lineages co-cladogenesis. FEMS Microbiol. Ecol. 2020, 96. [Google Scholar] [CrossRef]
- NDFF. NDFF Dissemination Atlas. Available online: http://verspreidingsatlas.nl (accessed on 4 September 2020).
- Team, R. Core. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012; Available online: https://www.R-project.org (accessed on 4 September 2019).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Wickham, H.; François, R.; Henry, L.; Müller, K. Dplyr: A Grammar of Data Manipulation. R Package Version 1.0.2. 2020. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 4 September 2020).



{kind=link}
{kind=link}
| Vertebrate (Order) | Vertebrate (Species) | Tested (n) | Clade I Positive (%) | Clade X Positive (%) | Samples Successfully Typed (18 S rRNA Typing) | >99% Identity (GenBank) |
|---|---|---|---|---|---|---|
| Artiodactyla | Bos taurus | 116 | 0 | 0 | - | |
| Bison bonasus | 19 | 0 | 4 (21%) | B. divergens (2) | AY046576 | |
| Capra aegagrus | 5 | 0 | 0 | - | ||
| Capreolus capreolus | 608 | 0 | 518 (85%) | B. capreoli (291) B. venatorum (3) Ambiguous (23) ** | AY726009 FJ215873 | |
| Cervus elaphus | 147 | 0 | 94 (64%) | B. divergens (26) Babesia sp. deer clade (32) | AY046576 HQ638138 | |
| Dama dama | 100 | 0 | 9 (9%) | B. capreoli (4) Babesia sp. deer clade-EU (1) | AY726009 HQ638138 | |
| Ovis aries | 634 | 0 | 10 (2%) | B. divergens (2) B. venatorum (3) | AY046576 FJ215873 | |
| Sus scrofa | 111 | 0 | 0 | - | ||
| Aves | 13 species | 99 | 0 | 0 | - | |
| Carnivora | Martes foina | 134 | 0 | 0 | - | |
| Martes martes | 128 | 0 | 0 | - | ||
| Meles meles | 128 | 108 (84%) | 0 | B. badger-type A (48) B. badger-type B (4) B. badger-type C (1) | KT223484 KT223485 MG799847 | |
| Mustela putorius | 242 | 0 | 0 | - | ||
| Nyctereutes procyonoides | 7 | 2 (29%) | 0 | B. vulpes (2) | AF188001 | |
| Vulpes vulpes | 173 | 124 (72%) | 0 | B. vulpes (58) | AF188001 | |
| Eulipotyphla | Talpa europaea | 125 | 0 | 0 | - | |
| Erinaceidae | Erinaceus europaeus | 32 | 0 | 0 | - | |
| Lagomorpha | Lepus europaeus | 150 | 0 | 0 | - | |
| Oryctolagus cuniculus | 88 | 0 | 0 | - | ||
| Perissodactyla | Equus ferus caballus | 15 | 0 | 0 | - | |
| Rodentia | Apodemus flavicollis | 29 | 0 | 0 | - | |
| Apodemus sylvaticus | 634 | 5 (0.8%) | 0 | B. microti (2) | KX161765 | |
| Castor fiber | 8 | 0 | 0 | - | ||
| Microtus arvalis | 100 | 3 (3%) | 0 | - | ||
| Myodes glareolus | 405 | 25 (6%) | 0 | B. microti (12) | KX161765 | |
| Ondatra zibethicus | 210 | 0 | 0 | - | ||
| Rattus rattus | 49 | 0 | 0 | - | ||
| Sciurus vulgaris | 115 | 0 | 0 | - |
| Study Acronym | I. ricinus | Method | Tested | Positive | Samples Successfully Typed/Sequenced | Period | Province | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| (Reference) | stage | n | n | % | B. capreoli | B. divergens | Babesia sp. deer clade | B. venatorum | years | sites (n) | |
| National Survey [44] | N + A | RLB | 857 | 13 | 0.015 | 13 | 2000–2010 | Gelderland (7) | |||
| N + A | RLB | 232 | 7 | 0.03 | 7 | 2006–2010 | Limburg (1) | ||||
| N + A | RLB | 1995 | 11 | 0.006 | 1 | 10 | 2000–2010 | Noord-Holland (4) | |||
| N + A | RLB | 242 | 1 | 0.004 | 1 | 2006–2010 | Brabant (1) | ||||
| N + A | RLB | 393 | 4 | 0.01 | 4 | 2006–2010 | Drenthe (2) | ||||
| N + A | RLB | 232 | 0 | 0 | 2006–2010 | Overijssel (2) | |||||
| N + A | RLB | 162 | 2 | 0.012 | 2 | 2006–2010 | Zuid-Holland (1) | ||||
| N | RLB | 79 | 3 | 0.038 | 3 | 2007–2010 | Friesland (1) | ||||
| N | RLB | 6 | 0 | 0 | 2007 | Groningen (1) | |||||
| N | RLB | 40 | 2 | 0.05 | 2 | 2007–2010 | Utrecht (1) | ||||
| Duin-Kruidberg [16] | L + N + A | RLB | 1.488 | 14 | 0.009 | 1 | 13 | 2003–2007 | Noord-Holland (1) | ||
| Lizard study [60] | N + A | RLB | 491 | 2 | 0.004 | 2 | 2007–2009 | Gelderland (8) | |||
| Drenthe * | N + A | RLB | 1727 | 3 | 0.002 | 3 | 2010–2012 | Drenthe (32) | |||
| Rodent study [61] * | N | qPCR | 7637 | 73 | 0.01 | 5 | 2 | 1 | 55 | 2012–2014 | Gelderland (2) |
| Urban ticks [62] | N | qPCR | 1780 | 19 | 0.011 | 2 | 16 | 2014–2016 | Antwerpen, Belgium (13) | ||
| A | qPCR | 268 | 3 | 0.011 | 3 | 2014–2016 | Antwerpen, Belgium (11) | ||||
| Cattle study [63] | N | qPCR | 112 | 0 | 0 | 2015 | Drenthe (1) | ||||
| N | qPCR | 181 | 2 | 0.011 | 1 | 2015 | Gelderland (1) | ||||
| N | qPCR | 191 | 1 | 0.005 | 1 | 2015 | Noord-Brabant (1) | ||||
| N | qPCR | 735 | 1 | 0.001 | 2015 | Noord-Holland (4) | |||||
| N | qPCR | 165 | 2 | 0.012 | 1 | 2015 | Overijssel (1) | ||||
| N | qPCR | 783 | 12 | 0.015 | 8 | 2015 | Utrecht (4) | ||||
| N | qPCR | 185 | 1 | 0.005 | 1 | 2015 | Zuid-Holland (1) | ||||
| De Groote Peel * | N | qPCR | 79 | 4 | 0.051 | 3 | 2017 | Brabant (1) | |||
| Sheep study * | N + A | qPCR | 1555 | 24 | 0.015 | 1 | 16 | 2017 | Drenthe (3) | ||
| Microbiome * | N | qPCR | 560 | 13 | 0.023 | 11 | 2018 | Gelderland (1) | |||
| N | qPCR | 520 | 2 | 0.004 | 1 | 2018 | Noord-Holland (1) | ||||
| N | qPCR | 1960 | 47 | 0.024 | 3 | 33 | 2018 | Utrecht (1) | |||
| Nijverdal * | N | qPCR | 1194 | 223 | 0.187 | 2019 | Overijssel (1) | ||||
| Total | 25849 | 489 | 1.9% | 11 | 4 | 1 | 210 | 2000–2019 | |||
| Study Acronym | I. ricinus | Method | Tested | Positive | Typing | Period | Province | |
|---|---|---|---|---|---|---|---|---|
| (Reference) | stage | n | n | % | Sequencing | years | sites (n) | |
| National Survey [44] | N + A | RLB | 857 | 6 | 0.7 | - | 2000–2010 | Gelderland (7) |
| N + A | RLB | 1995 | 2 | 0.1 | - | 2000–2010 | Noord-Holland (4) | |
| N + A | RLB | 242 | 0 | 0 | - | 2006–2010 | Brabant (1) | |
| N + A | RLB | 393 | 0 | 0 | - | 2006–2010 | Drenthe (2) | |
| N + A | RLB | 232 | 0 | 0 | - | 2006–2010 | Limburg (1) | |
| N + A | RLB | 232 | 8 | 3.4 | - | 2006–2010 | Overijssel (2) | |
| N + A | RLB | 162 | 0 | 0 | - | 2006–2010 | Zuid-Holland (1) | |
| N | RLB | 79 | 1 | 1.3 | - | 2007–2010 | Friesland (1) | |
| N | RLB | 6 | 0 | 0 | - | 2007 | Groningen (1) | |
| N | RLB | 40 | 0 | 0 | - | 2007–2010 | Utrecht (1) | |
| Duin-Kruidberg [16] | N + A | RLB | 908 | 2 | 0.20 | B. microti (2) | 2003–2007 | Noord-Holland (1) |
| Lizard study [60] | N + A | RLB | 491 | 6 | 1.20 | - | 2007–2009 | Gelderland (8) |
| Drenthe (this study) | N + A | RLB | 1727 | 0 | 0 | - | 2010–2012 | Drenthe (32) |
| Rodent study [61] | N | qPCR | 7637 | 393 | 5.1 | B. microti (39) | 2012–2014 | Gelderland (2) |
| Cattle study [63] * | N | qPCR | 112 | 0 | 0 | - | 2015 | Drenthe (1) |
| N | qPCR | 181 | 14 | 7.7 | B. microti (4) | 2015 | Gelderland (1) | |
| N | qPCR | 191 | 0 | 0 | - | 2015 | Noord-Brabant (1) | |
| N | qPCR | 735 | 0 | 0 | - | 2015 | Noord-Holland (4) | |
| N | qPCR | 165 | 0 | 0 | - | 2015 | Overijssel (1) | |
| N | qPCR | 783 | 0 | 0 | - | 2015 | Utrecht (4) | |
| N | qPCR | 185 | 0 | 0 | - | 2015 | Zuid-Holland (1) | |
| De Groote Peel * | N | qPCR | 79 | 0 | 0 | - | 2017 | Brabant (1) |
| Nijverdal * | N | qPCR | 1194 | 50 | 4.20 | - | 2019 | Overijssel (1) |
| Total | 18626 | 482 | 2.6 | B. microti (45) | 2000–2019 | |||
| Babesia Clade | Pathogen | Positive | Observed | Expected | P-Value | Odds Ratio |
|---|---|---|---|---|---|---|
| Babesia Clade I | B. burgdorferi sl | 1292 | 273 | 65 | 1.10 × 10−15 | 11.72 |
| B. miyamotoi | 261 | 14 | 13 | 1 | 1.08 | |
| N. mikurensis | 939 | 129 | 47 | 1.10 × 10−15 | 3.86 | |
| A. phagocytophilum | 642 | 21 | 32 | 1.80 × 10−1 | 0.62 | |
| Babesia Clade X | 123 | 7 | 6 | 1 | 1.15 | |
| Babesia Clade X | B. burgdorferi sl | 1292 | 17 | 18 | 1 | 0.93 |
| B. miyamotoi | 261 | 6 | 4 | 0.90 | 1.69 | |
| N. mikurensis | 939 | 6 | 13 | 0.18 | 0.42 | |
| A. phagocytophilum | 642 | 2 | 9 | 0.04 | 0.2 | |
| Babesia Clade I | 442 | 7 | 6 | 1 | 1.15 |
| Babesia | Target | Primer (Name) | Primer (Sequence) | Purpose | Size (bp) | Reference |
|---|---|---|---|---|---|---|
| Clade I | ITS | Bmicr_ITS_F | 5’-CTCACACAACGATGAAGGACGCA-3’ | qPCR | 103 bp | [136] |
| Bmicr_ITS_R | 5’-AACAGAGGCAGTGTGTACAATACATTCAGA-3’ | |||||
| Bmicr_ITS_Px1 | 5′- Atto520-GCA+GAATTTAG+CAAAT+CAACAGG- BHQ1-3′ | |||||
| Clade X | 18SrRNA | Bab_18SrRNA-F | 5’-CAGCTTGACGGTAGGGTATTGG-3’ | qPCR | 62 bp | [129] |
| Bab_18SrRNA-R | 5’-TCGAACCCTAATTCCCCGTTA-3’ | |||||
| Bab_18SrRNA-P | 5’-Atto647N-CGAGGCAGCAACGG-MGB-BHQ2-3’ | |||||
| Babesia spp. | 18SrRNA | Bath-Fn | 5’-TAAGAATTTCACCTCTGACAGTTA-3’ | PCR/SEQ | ±420 bp | [16] |
| Bath-Rn | 5’-ACACAGGGAGGTAGTGACAAG-3’ | |||||
| Clade I | COI | Cox1F133 | GGAGAGCTAGGTAGTAGTGGAGATAGG | PCR/SEQ | 1023 bp | [134] |
| Cox1R1130 | GTGGAAGTGAGCTACCACATACGCTG | |||||
| Clade X | COI | Cox1_Bab_For1 | 5′-ATWGGATTYTATATGAGTAT-3′ | PCR/SEQ | ±1250 bp | [7] |
| Cox1_Bab_Rev1 | 5′-ATAATCWGGWATYCTCCTTGG-3′ | |||||
| B. venatorum | COI | Bven-F159 | 5’-ATTGGAAGTGGTACTGGTTGGACTT-3’ | PCR | 538 bp | This study |
| Bven-R696 | 5’-GACATCATTACGATTCCTATGC-3’ | |||||
| B. divergens | COI | Bdiv-F165 | 5’-AGTGGAACTGGGTGGACATTGTAC-3’ | PCR | 234 bp | This study |
| Bdiv-R398 | 5’-TACCGGCAATGACAAAAGTAG-3’ | |||||
| B. capreoli | COI | Bcap-F165 | 5’-AGTGGAACAGGATGGACGCTATAT-3’ | PCR | 443 bp | This study |
| Bcap-R607 | 5’-GTCTGATTACCGAACACTTCC-3’ | |||||
| Babesia sp. deer clade | COI | Bodo-F360 | 5’-CTTTGACTGCTTTCTTGTTG-3’ | PCR | 434 bp | This study |
| Bodo-R793 | 5’-ATCATAACAATTCCTATGCTC-3’ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azagi, T.; Jaarsma, R.I.; Docters van Leeuwen, A.; Fonville, M.; Maas, M.; Franssen, F.F.J.; Kik, M.; Rijks, J.M.; Montizaan, M.G.; Groenevelt, M.; et al. Circulation of Babesia Species and Their Exposure to Humans through Ixodes ricinus. Pathogens 2021, 10, 386. https://doi.org/10.3390/pathogens10040386
Azagi T, Jaarsma RI, Docters van Leeuwen A, Fonville M, Maas M, Franssen FFJ, Kik M, Rijks JM, Montizaan MG, Groenevelt M, et al. Circulation of Babesia Species and Their Exposure to Humans through Ixodes ricinus. Pathogens. 2021; 10(4):386. https://doi.org/10.3390/pathogens10040386
Chicago/Turabian StyleAzagi, Tal, Ryanne I. Jaarsma, Arieke Docters van Leeuwen, Manoj Fonville, Miriam Maas, Frits F. J. Franssen, Marja Kik, Jolianne M. Rijks, Margriet G. Montizaan, Margit Groenevelt, and et al. 2021. "Circulation of Babesia Species and Their Exposure to Humans through Ixodes ricinus" Pathogens 10, no. 4: 386. https://doi.org/10.3390/pathogens10040386