
Differentiation of Gastric Helicobacter Species Using MALDI-TOF Mass Spectrometry

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Helicobacter Isolates and Growth Conditions
4.2. Sample Preparation for MALDI-TOF MS Analysis
4.3. Generation of (Reference) Spectra
4.4. Testing of Reproducibility
4.5. Testing of Culture Conditions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Solnick, J.V. Clinical significance of Helicobacter species other than Helicobacter pylori. Clin. Inf. Dis. 2003, 36, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Haesebrouck, F.; Pasmans, F.; Flahou, B.; Chiers, K.; Baele, M.; Meyns, T.; Decostere, A.; Ducatelle, R. Gastric helicobacters in domestic animals and nonhuman primates and their significance for human health. Clin. Microbiol. Rev. 2009, 22, 202–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labigne, A.; Cussac, V.; Courcoux, P. Shuttle cloning and nucleotide sequences of Helicobacter pylori genes responsible for urease activity. J. Bacteriol. 1991, 173, 1920–1931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weeks, D.L.; Eskandari, S.; Scott, D.R.; Sachs, G. A H+-gated urea channel: The link between Helicobacter pylori urease and gastric colonization. Science 2000, 287, 482–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solnick, J.V.; Schauer, D.B. Emergence of diverse Helicobacter species in the pathogenesis of gastric and enterohepatic diseases. Clin. Microbiol. Rev. 2001, 14, 59–97. [Google Scholar] [CrossRef] [Green Version]
- Sterzenbach, T.; Lee, S.K.; Brenneke, B.; von Goetz, F.; Schauer, D.B.; Fox, J.G.; Suerbaum, S.; Josenhans, C. Inhibitory effect of enterohepatic Helicobacter hepaticus on innate immune responses of mouse intestinal epithelial cells. Infect. Immun. 2007, 75, 2717–2728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamani, M.; Ebrahimtabar, F.; Zamani, V.; Miller, W.H.; Alizadeh-Navaei, R.; Shokri-Shirvani, J.; Derakhshan, M.H. Systemic review with meta-analysis: The worldwide prevalence of Helicobacter pylori infection. Aliment. Pharm. Ther. 2018, 47, 868–876. [Google Scholar] [CrossRef] [Green Version]
- Kusters, J.G.; van Vliet, A.H.M.; Kuipers, E.J. Pathogenesis of Helicobacter pylori infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Øverby, A.; Murayama, S.Y.; Michimae, H.; Suzuki, H.; Suzuki, M.; Serizawa, H.; Tamura, R.; Nakamura, S.; Takahashi, S.; Nakamura, M. Prevalence of gastric non-Helicobacter pylori helicobacters in Japanese patients with gastric disease. Digestion 2017, 95, 61–66. [Google Scholar] [CrossRef]
- De Groote, D.; van Doorn, L.J.; Ducatelle, R.; Verschuuren, A.; Tilmant, K.; Quint, W.H.; Haesebrouck, F.; Vandamme, P. Phylogenetic characterization of ‘Candidatus Helicobacter bovis’, a new gastric helicobacter in cattle. Int. J. Syst. Bacteriol. 1999, 49, 1707–1715. [Google Scholar] [CrossRef]
- De Groote, D.; van Doorn, L.J.; Van den Bulck, K.; Vandamme, P.; Vieth, M.; Stolte, M.; Debongnie, J.C.; Burette, A.; Haesebrouck, F.; Ducatelle, R. Detection of non-pylori Helicobacter species in "Helicobacter heilmannii"-infected humans. Helicobacter 2005, 10, 398–406. [Google Scholar] [CrossRef]
- García-Amado, M.A.; Al-Soud, W.A.; Borges-Landaéz, P.; Contreras, M.; Cedeño, S.; Baéz-Ramírez, E.; Domínguez-Bello, M.G.; Wadström, T.; Gueneau, P. Non-pylori helicobacteraceae in the upper digestive tract of asymptomatic Venezuelan subjects: Detection of Helicobacter cetorum-like and Candidatus Wolinella africanus-like DNA. Helicobacter 2007, 12, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Flahou, B.; Haesebrouck, F.; Smet, A. Non-Helicobacter pylori helicobacter infections in humans and animals. In Helicobacter Pylori Research, from Bench to bedside; Backert, S., Yamaoka, Y., Eds.; Springer: Tokyo, Japan, 2016; pp. 233–269. [Google Scholar]
- Goto, K.; Ohashi, H.; Ebukuro, S.; Itoh, K.; Tohma, Y.; Takakura, A.; Wakana, S.; Ito, M.; Itoh, T. Isolation and characterization of Helicobacter species from the stomach of the house musk shrew (Suncus murinus) with chronic gastritis. Curr. Microbiol. 1998, 37, 44–51. [Google Scholar] [CrossRef]
- Flahou, B.; Modrý, D.; Pomajbíková, K.; Petrželková, K.J.; Smet, A.; Ducatelle, R.; Pasmans, F.; Sá, R.M.; Todd, A.; Hashimoto, C.; et al. Diversity of zoonotic enterohepatic Helicobacter species and detection of a putative novel gastric Helicobacter species in wild and wild-born captive chimpanzees and western lowland gorillas. Vet. Microbiol. 2014, 174, 186–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruntar, I.; Papić, B.; Pate, M.; Zajc, U.; Ocepek, M.; Kušar, D. Helicobacter labacensis sp. nov., Helicobacter mehlei sp. nov., and Helicobacter vulpis sp. nov., isolated from gastric mucosa of red foxes (Vulpes Vulpes). Int. J. Syst. Evol. Microbiol. 2020, 70, 2395–2404. [Google Scholar] [CrossRef]
- Berlamont, H.; Smet, A.; De Bruyckere, S.; Boyen, F.; Ducatelle, R.; Haesebrouck, F.; De Witte, C. Antimicrobial susceptibility pattern of Helicobacter suis isolates from pigs and macaques. Vet. Microbiol. 2019, 239, 108459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matos, R.; De Witte, C.; Smet, A.; Berlamont, H.; De Bruyckere, S.; Amorim, I.; Gärtner, F.; Haesebrouck, F. Antimicrobial susceptibility pattern of Helicobacter heilmannii and Helicobacter ailurogastricus isolates. Microorganisms 2020, 8, 957. [Google Scholar] [CrossRef] [PubMed]
- Owen, R.J. Helicobacter-species classification and identification. Br. Med. Bull. 1998, 54, 17–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bento-Miranda, M.; Figueiredo, C. Helicobacter heilmannii sensu lato: An overview of the infection in humans. World J. Gastroenterol. 2014, 20, 17779–17787. [Google Scholar] [CrossRef] [Green Version]
- Blaecher, C.; Bauwens, E.; Tay, A.; Peters, F.; Dobbs, S.; Dobbs, J.; Charlett, A.; Ducatelle, R.; Haesebrouck, F.; Smet, A. A novel isolation protocol and probe-based RT-PCR for diagnosis of gastric infections with the zoonotic pathogen Helicobacter suis. Helicobacter 2016, 22, e12369. [Google Scholar] [CrossRef] [PubMed]
- Anhalt, J.P.; Fenselau, C. Identification of bacteria using mass spectrometry. Anal. Chem. 1975, 47, 219–225. [Google Scholar] [CrossRef]
- Cain, T.; Lubman, D.M.; Weber, W.J. Differentiation of bacteria using protein profiles from matrix-assisted laser-desorption ionization time-of-flight mass-spectrometry. Rapid Commun. Mass Spectrom. 1994, 8, 1026–1030. [Google Scholar] [CrossRef]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieser, A.; Schneider, L.; Jung, J.; Schubert, S. MALDI-TOF MS in microbiological diagnostics-identification of microorganisms and beyond (mini review). Appl. Microbiol. Biotechnol. 2012, 93, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Dash, H.R.; Mangwani, N.; Chakraborty, J.; Kumari, S. Understanding molecular identification and polyphasic taxonomic approaches for genetic relatedness and phylogenetic relationships of microorganisms. J. Microbiol. Methods 2014, 103, 80–100. [Google Scholar] [CrossRef]
- Murray, P.R. What is new in clinical microbiology-microbial identification by MALDI-TOF mass spectrometry: A paper from the 2011 William Beaumont Hospital Symposium on molecular pathology. J. Mol. Diagn. 2012, 14, 419–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seng, P.; Drancourt, M.; Gouriet, F.; La Scola, B.; Fournier, P.E.; Rolain, J.M.; Raoult, D. Ongoing revolution in bacteriology: Routine identification of bacteria by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Clin. Infect. Dis. 2009, 49, 543–551. [Google Scholar] [CrossRef]
- Smet, A.; Yahara, K.; Rossi, M.; Tay, A.; Backert, S.; Armin, E.; Fox, J.G.; Flahou, B.; Ducatelle, R.; Haesebrouck, F.; et al. Macroevolution of gastric Helicobacter species unveils interspecies admixture and time of divergence. ISME J. 2018, 12, 2518–2531. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.G.; Correa, P.; Taylor, N.S.; Lee, A.; Otto, G.; Murphy, J.C.; Rose, R. Helicobacter mustelae-associated gastritis in ferrets: An animal model of Helicobacter pylori gastritis in humans. Gastroenterology 1990, 99, 352–361. [Google Scholar] [CrossRef]
- Fox, J.G.; Marin, R.P. Helicobacter mustelae infection in ferrets: Pathogenesis, epizootiology, diagnosis and treatment. Sem. Avian Exot. Pet Med. 2001, 10, 36–44. [Google Scholar] [CrossRef]
- Croxatto, A.; Prod’hom, G.; Greub, G. Applications of MALDI-TOF mass spectrometry in clinical diagnostic microbiology. FEMS Microbiol. Rev. 2012, 36, 380–407. [Google Scholar] [CrossRef]
- Mörtelmaier, C.; Panda, S.; Robertson, I.; Krell, M.; Christodoulou, M.; Reichardt, N.; Mulder, I. Identification performance of MALDI-ToF-MS upon mono- and bi-microbial cultures is cell number and culture proportion dependent. Anal. Bioanal. Chem. 2019, 411, 7027–7038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, K.S.; Kim, Y.H. Rapid and robust MALDI-TOF MS techniques for microbial identification: A brief overview of their diverse applications. J. Micriobiol. 2018, 56, 209–216. [Google Scholar] [CrossRef]
- Chen, J.H.; Ho, P.L.; Kwan, G.S.W.; She, K.K.K.; Siu, G.K.H.; Cheng, V.C.C.; Yuen, K.Y.; Yam, W.C. Direct bacterial identification in positive blood cultures by use of two commercial matrix-assisted laser desorption ionization-time of flight mass spectrometry systems. J. Clin. Microbiol. 2013, 51, 1733–1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitagawa, K.; Shigemura, K.; Onuma, K.I.; Nishida, M.; Fujiwara, M.; Kobayashi, S.; Yamasaki, M.; Nakamura, T.; Yamamichi, F.; Shirakawa, T.; et al. Improved bacterial identification directly from urine samples with matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. J. Clin. Lab. Anal. 2018, 32, e22301. [Google Scholar] [CrossRef] [Green Version]
- Oviaño, M.; Rodríguez-Sánchez, B.; Gómara, M.; Alcalá, L.; Zvezdanova, E.; Ruíz, A.; Velasco, D.; Gude, M.J.; Bouza, E.; Bou, G. Direct identification of clinical pathogens from liquid culture media by MALDI-TOF MS analysis. Clin. Microbiol. Infect. 2018, 24, 624–629. [Google Scholar] [CrossRef] [Green Version]
- Bokma, J.; Van Driessche, L.; Deprez, P.; Haesebrouck, F.; Vahl, M.; Weesendorp, E.; Deurenberg, R.H.; Pardon, B.; Boyen, F. Rapid identification of Mycoplasma bovis strains from bovine bronchoalveolar lavage fluid with Matrix-Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry after enrichment procedure. J. Clin. Microbiol. 2020, 58, e00004-20. [Google Scholar] [CrossRef] [PubMed]
- Spergser, J.; Hess, C.; Loncaric, I.; Ramírez, A.S. Matrix-assisted laser desorption ionization-time of flight mass spectrometry is a superior diagnostic tool for the identification and differentiation of mycoplasmas isolated from animals. J. Clin. Microbiol. 2019, 57, e00316-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Driessche, L.; Bokma, J.; Deprez, P.; Haesebrouck, F.; Boyen, F.; Pardon, B. Rapid identification of respiratory bacterial pathogens from bronchoalveolar lavage fluid in cattle by MALDI-TOF MS. Sci. Rep. 2019, 9, 18381. [Google Scholar] [CrossRef]
- Welker, M. Proteomics for routine identification of microorganisms. Proteomics 2011, 11, 3143–3153. [Google Scholar] [CrossRef] [PubMed]
- Valentine, N.; Wunschel, S.; Wunschel, D.; Petersen, C.; Wahl, K. Effect of culture conditions on microorganism identification by matrix-assisted laser desorption ionization mass spectrometry. Appl. Environ. Microbiol. 2005, 71, 58–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wunschel, D.S.; Hill, E.A.; McLean, J.S.; Jarman, K.; Gorby, Y.A.; Valentine, N.; Wahl, K. Effects of varied pH, growth rate and temperature using controlled fermentation and batch culture on matrix assisted laser desorption/ionization whole cell protein fingerprints. J. Microbiol. Methods 2005, 62, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Mazzeo, M.F.; Sorrentino, A.; Gaita, M.; Cacace, G.; Di Stasio, M.; Facchiano, A.; Comi, G.; Malorni, A.; Siciliano, R.A. Matrix-assisted laser desorption ionization–time of flight mass spectrometry for the discrimination of food-borne microorganisms. Appl. Environ. Microbiol. 2006, 72, 1180–1189. [Google Scholar] [CrossRef] [Green Version]
- Alispahic, M.; Hummel, K.; Jandreski-Cvetkovic, D.; Nöbauer, K.; Razzazi-Fazeli, E.; Hess, M.; Hess, C. Species-specific identification and differentiation of Arcobacter, Helicobacter and Campylobacter by full-spectral matrix-associated laser desorption/ionization time of flight mass spectrometry analysis. J. Med. Microbiol. 2010, 59, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Balážová, T.; Makovcová, J.; Šedo, O.; Slaný, M.; Faldyna, M.; Zdráhal, Z. The influence of culture conditions on the identification of Mycobacterium species by MALDI-TOF MS profiling. FEMS Microbiol. Lett. 2014, 353, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Lagacé-Wiens, P.R.S.; Abbott, A.A.; Karlowsky, J.A. CHROMagarTM orientation urine culture medium produces matrix-assisted laser desorption-time-of-flight mass spectrometry spectra misidentified as Mycoplasma arginini and Mycoplasma alkalescens. Diagn. Microbiol. Infect. Dis. 2019, 94, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Bokma, J.; Pardon, B.; Deprez, P.; Haesebrouck, F.; Boyen, F. Non-specific, agar medium-related peaks can result in false positive Mycoplasma alkalescens and Mycoplasma arginini identification by MALDI-TOF MS. Res. Vet. Sci. 2020, 130, 139–143. [Google Scholar] [CrossRef]
- Parisi, D.; Magliulo, M.; Nanni, P.; Casale, M.; Forina, M.; Roda, A. Analysis and classification of bacteria by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry and a chemometric approach. Anal. Bioanal. Chem. 2008, 391, 2127–2134. [Google Scholar] [CrossRef]
- Barbuddhe, S.B.; Maier, T.; Schwarz, G.; Kostrzewa, M.; Hof, H.; Domann, E.; Chakraborty, T.; Hain, T. Rapid identification and typing of Listeria species by matrix-assisted laser desorption ionization-time of flight mass spectrometry. Appl. Environ. Microbiol. 2008, 74, 5402–5407. [Google Scholar] [CrossRef] [Green Version]
- Baele, M.; Decostere, A.; Vandamme, P.; Ceelen, L.; Hellemans, A.; Mast, J.; Chiers, K.; Ducatelle, R.; Haesebrouck, F. Isolation and characterization of Helicobacter suis sp. nov. from pig stomachs. Int. J. Syst. Evol. Microbiol. 2008, 58, 1350–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Ducatelle, R.; Pasmans, F.; Smet, A.; Haesebrouck, F.; Flahou, B. Multilocus sequence typing of the porcine and human gastric pathogen Helicobacter suis. J. Clin. Microbiol. 2013, 51, 920–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joosten, M.; Lindén, S.; Rossi, M.; Tay, A.C.Y.; Skoog, E.; Padra, M.; Peters, F.; Perkins, T.; Vandamme, P.; Van Nieuwerburgh, F.; et al. Divergence between the highly virulent zoonotic pathogen Helicobacter heilmannii and its closest relative, the low-virulence “Helicobacter ailurogastricus” sp. nov. Infect. Immun. 2016, 84, 293–306. [Google Scholar] [CrossRef] [Green Version]
- Harper, C.G.; Feng, Y.; Xu, S.; Taylor, N.S.; Kinsel, M.; Dewhirst, F.E.; Paster, B.J.; Greenwell, M.; Levine, G.; Rogers, A.; et al. Helicobacter cetorum sp. nov., a urease-positive Helicobacter species isolated from dolphins and whales. J. Clin. Microbiol. 2002, 40, 4536–4543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harper, C.G.; Whary, M.T.; Feng, Y.; Rhinehart, H.L.; Wells, R.S.; Xu, S.; Taylor, N.S.; Fox, J.G. Comparison of diagnostic techniques for Helicobacter cetorum infection in Wild Atlantic Bottlenose Dolphins (Tursiops truncates). J. Clin. Microbiol. 2003, 41, 2842–2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]



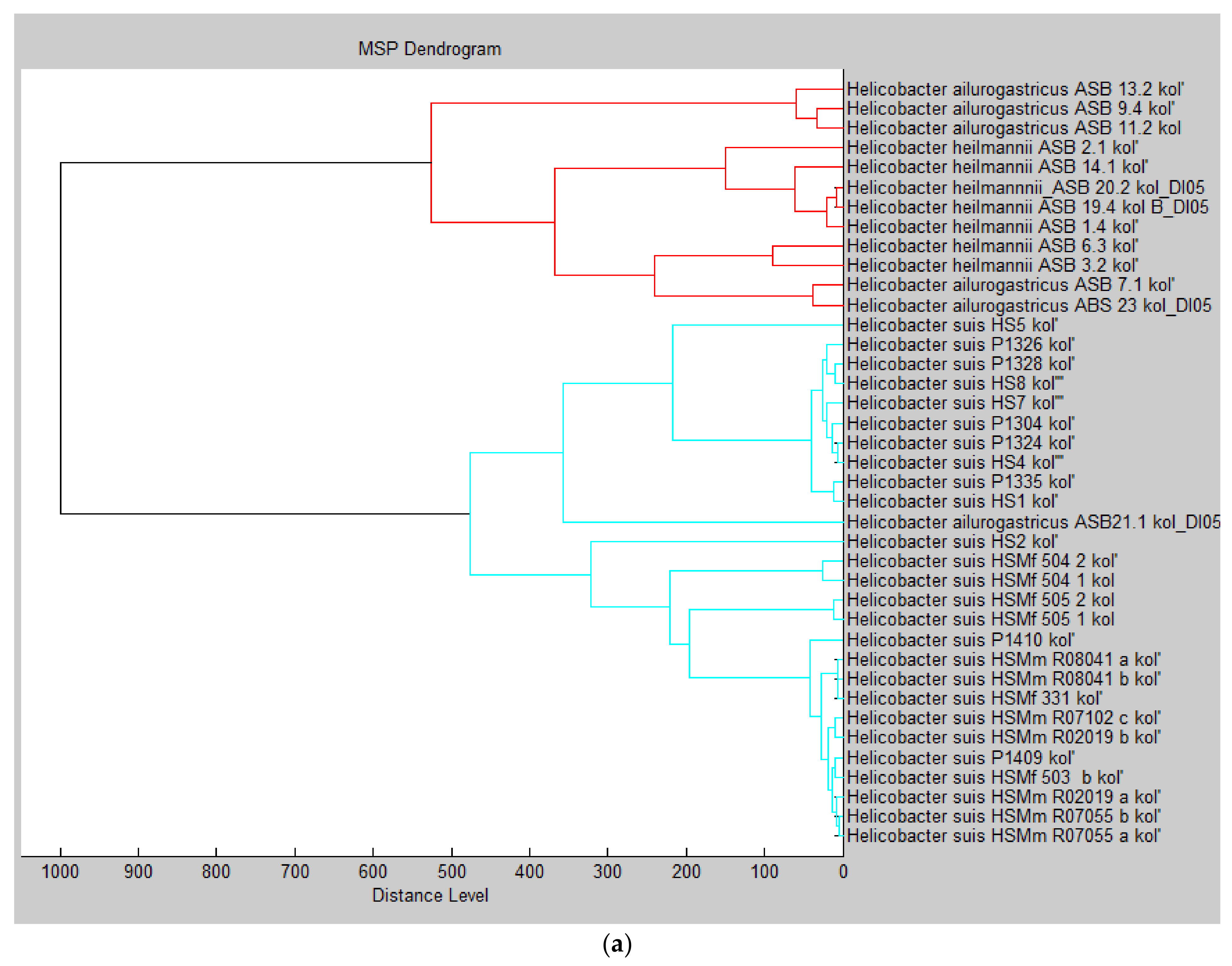{kind=link}
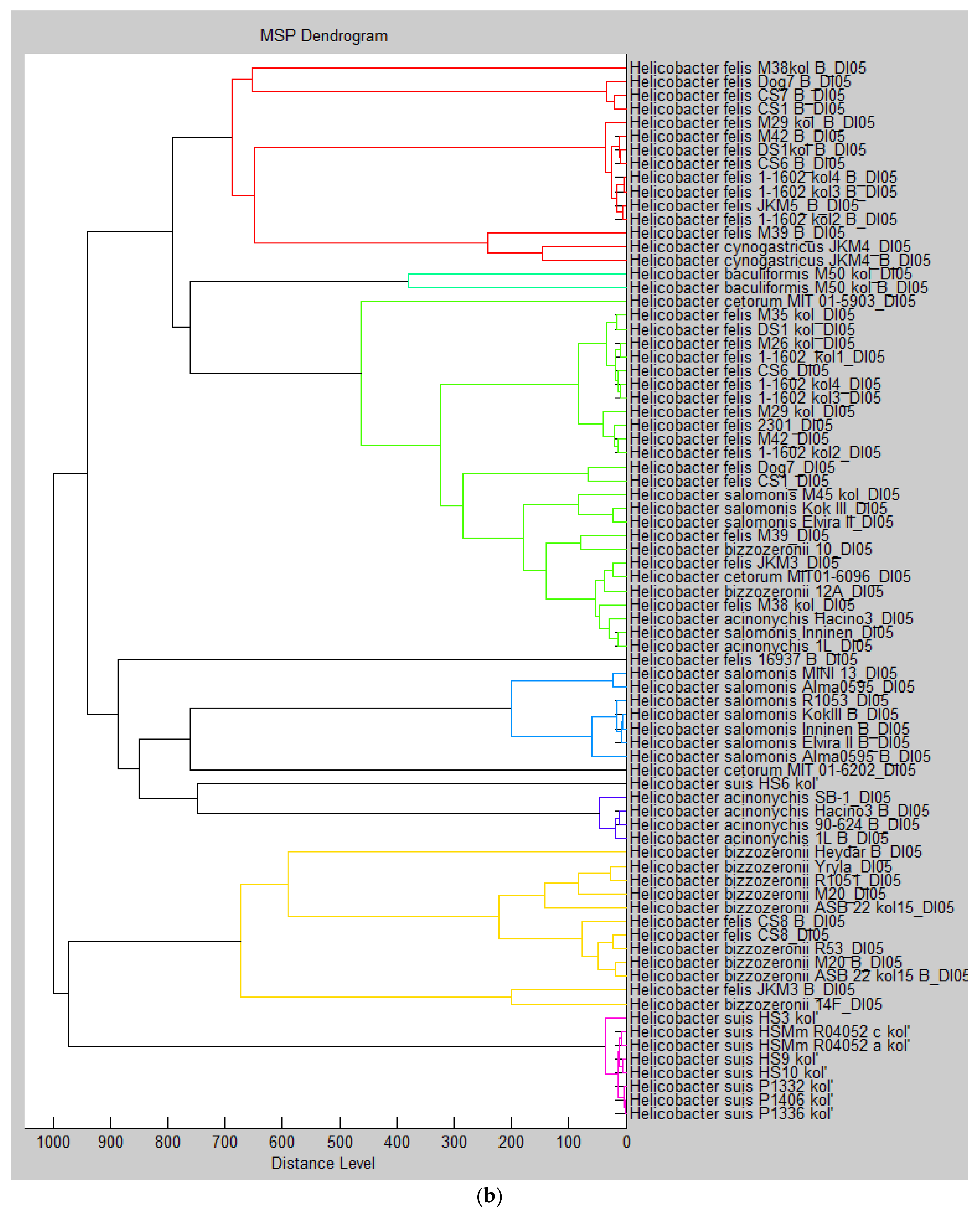{kind=link}
| Species | Number of Isolates | Amount of MSPs Created | Hosts of Isolation | MALDI-TOF log Score Best Correct Match Range | Best Incorrect Match (Maximum log Score) | Log difference Best Correct Match − Best Incorrect Match |
|---|---|---|---|---|---|---|
| H. acinonychis | 4 | 6 | Wild felines | 2.58–2.79 | 3rd match | 0.12 |
| H. salomonis (2.42) | ||||||
| H. ailurogastricus | 6 | 6 | Cat | 2.49–2.82 | 7th match | 0.80 |
| H. heilmannii (2.02) | ||||||
| H. baculiformis | 1 | 2 | Cat | 2.69–2.75 | 3rd match | 0.67 |
| H. cetorum (2.02) | ||||||
| H. bizzozeronii | 9 | 11 | Dog, cat, human | 2.40–2.81 | 4th match H. felis (2.46) | 0.08 |
| H. cetorum | 3 | 3 | Dolphin | 2.22–2.74 | 2nd match H. bizzozeronii (2.16) | 0.06 |
| H. cynogastricus | 1 | 2 | Dog | 2.67–2.85 | 3rd match H. felis (2.23) | 0.62 |
| H. felis | 20 | 33 | Dog, cat | 2.32–2.81 | 2nd match H. bizzozeronii (2.45) | 0.14 |
| H. heilmannii | 7 | 7 | Cat | 2.56–2.82 | 6th match H. ailurogastricus (1.96) | 0.64 |
| H. salomonis | 7 | 11 | Dog | 2.48–2.81 | 2nd match H. acinonychis (2.61) | 0.05 |
| H. suis | 35 | 35 | Pigs, non-human primates | 2.45–2.88 | 5th match H. felis (2.08) | 0.66 |
| Total | 93 | 116 | 2.22–2.88 | 1.96–2.61 | 0.05–0.80 |
| Individual Spectra of Isolate | Log Score Best MSP Match | Second-Best Match | Log Score Second-Best MSP Match * | Log Difference |
|---|---|---|---|---|
| H. ailurogastricus ASB 21.1 D | 2.49 | H. heilmannii ASB1.4 kol B | 1.91 | 0.58 |
| H. bizzozeronii 10 D | 2.46 | H. felis JKM3 D | 2.34 | 0.12 |
| H. bizzozeronii 12A D | 2.40 | H. cetorum MIT 01-6096 D | 2.22 | 0.18 |
| H. bizzozeronii Heydar B | 2.78 | H. felis CS8 B | 1.87 | 0.91 |
| H. bizzozeronii R1051 D | 2.60 | H. felis CS8 D | 2.42 | 0.18 |
| H. cetorum MIT 01-5903 D | 2.74 | H. felis M29 kol D | 1.90 | 0.84 |
| H. cetorum MIT 01-6096 D | 2.22 | H. bizzozeronii 12A D | 2.16 | 0.06 |
| H. felis CS8 D | 2.59 | H. bizzozeronii R1051 D | 2.45 | 0.14 |
| H. felis JKM3 D | 2.57 | H. cetorum MIT 01-6096 D | 2.41 | 0.16 |
| H. felis M38 D | 2.45 | H. acinonychis Hacino3 D | 1.80 | 0.65 |
| H. felis M39 D | 2.54 | H. cetorum MIT 01-6096 D | 2.19 | 0.35 |
| H. salomonis Inkinen D | 2.66 | H. acinonychis 1L D | 2.61 | 0.05 |
| Species | Isolate | Log Score B to D | Log Score D to B |
|---|---|---|---|
| H. acinonychis | 1L | 0.76 | 1.48 |
| Hacino3 | 1.10 | 1.79 | |
| H. baculiformis | M50 | 2.45 | 2.44 |
| H. bizzozeronii | M20 | 1.97 | 2.13 |
| ASB 22 kol 15 | 1.72 | 2.16 | |
| H. cynogastricus | JKM4 | 2.34 | 2.43 |
| H. felis | CS1 | 2.22 | 2.06 |
| CS6 | 1.83 | 1.65 | |
| CS8 | 2.31 | 1.92 | |
| DS1 | 2.12 | 2.30 | |
| Dog7 | 1.97 | 1.97 | |
| JKM3 | 1.67 | 1.02 | |
| M29 | 2.09 | 2.31 | |
| M38 | 1.76 | 1.31 | |
| M39 | < 0 | 1.04 | |
| M42 | 1.56 | 1.66 | |
| 1-1602 kol2 | 1.78 | 1.30 | |
| 1-1602 kol3 | 2.05 | 2.00 | |
| 1-1602 kol4 | 2.16 | 1.75 | |
| H. salomonis | KokIII | 1.90 | 1.56 |
| Alma0595 | 2.10 | 1.51 | |
| Inkinen | 0.69 | <0 | |
| Elvira II | 1.98 | 1.90 | |
| <0–2.45 | <0–2.44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berlamont, H.; De Witte, C.; De Bruyckere, S.; Fox, J.G.; Backert, S.; Smet, A.; Boyen, F.; Haesebrouck, F. Differentiation of Gastric Helicobacter Species Using MALDI-TOF Mass Spectrometry. Pathogens 2021, 10, 366. https://doi.org/10.3390/pathogens10030366
Berlamont H, De Witte C, De Bruyckere S, Fox JG, Backert S, Smet A, Boyen F, Haesebrouck F. Differentiation of Gastric Helicobacter Species Using MALDI-TOF Mass Spectrometry. Pathogens. 2021; 10(3):366. https://doi.org/10.3390/pathogens10030366
Chicago/Turabian StyleBerlamont, Helena, Chloë De Witte, Sofie De Bruyckere, James G. Fox, Steffen Backert, Annemieke Smet, Filip Boyen, and Freddy Haesebrouck. 2021. "Differentiation of Gastric Helicobacter Species Using MALDI-TOF Mass Spectrometry" Pathogens 10, no. 3: 366. https://doi.org/10.3390/pathogens10030366