
Contribution of Coagulase and Its Regulator SaeRS to Lethality of CA-MRSA 923 Bacteremia


Abstract
:1. Introduction
2. Results
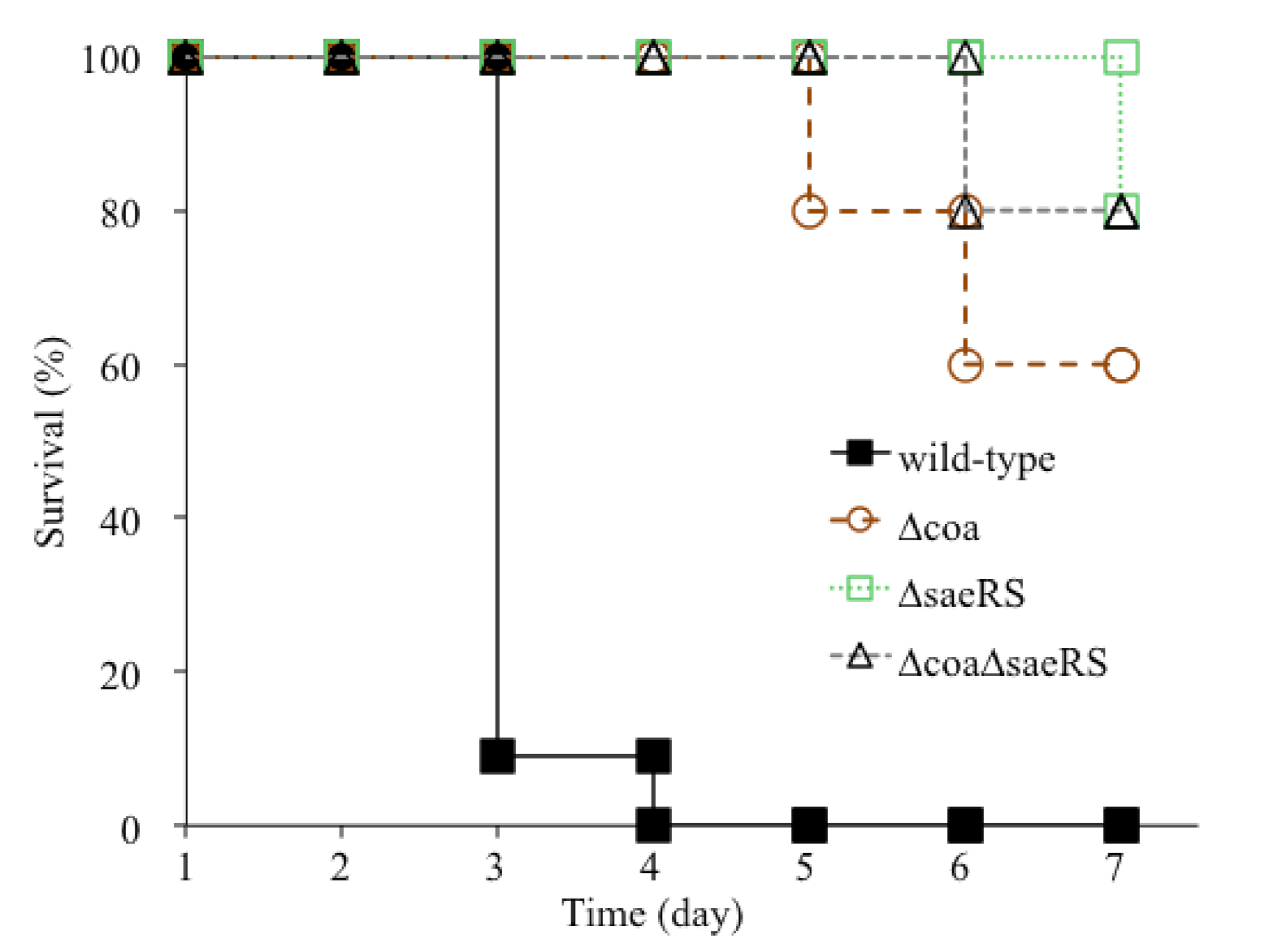
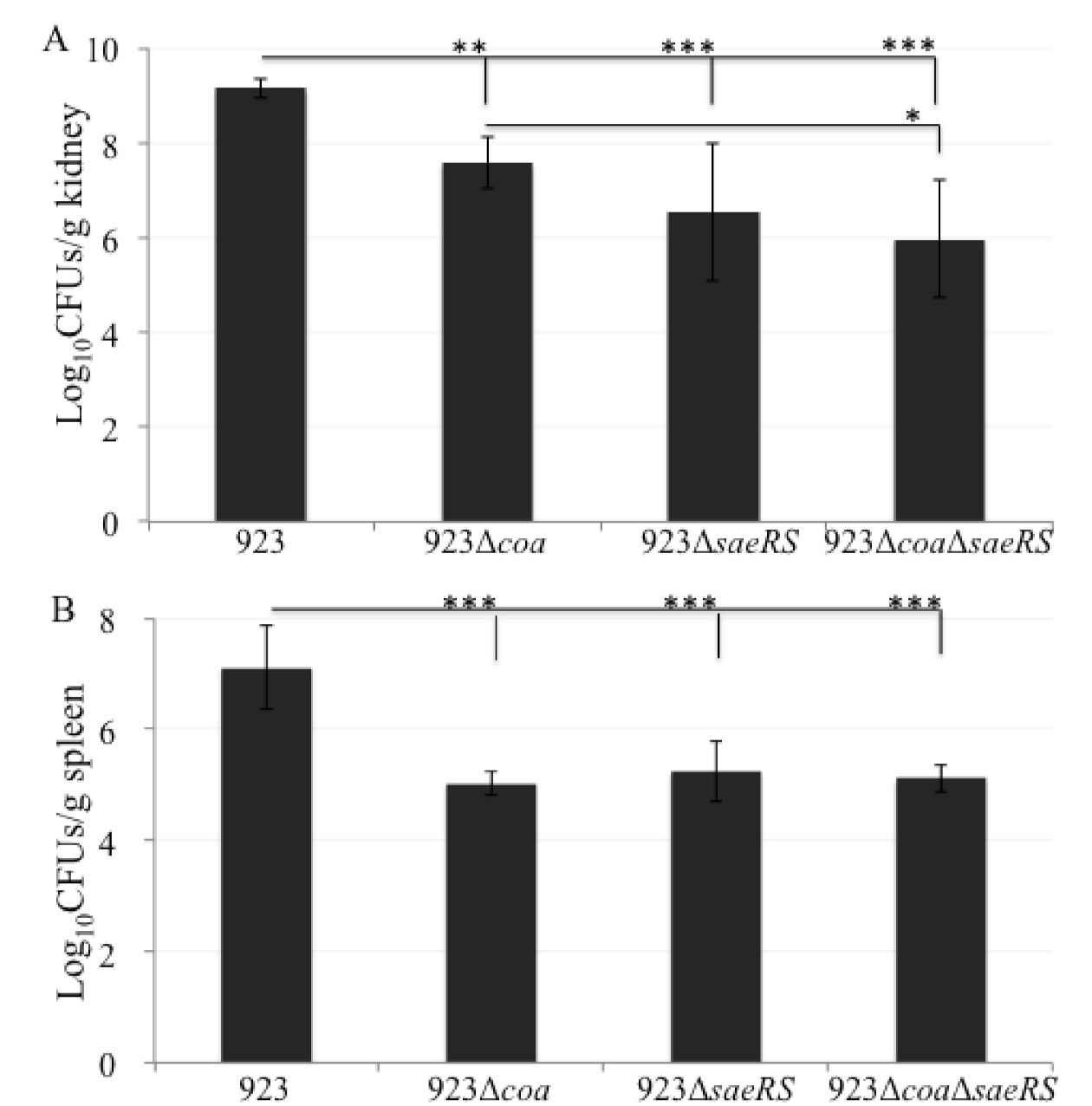
2.1. The Deletion Mutation of coa Significantly Alleviated the Lethality of CA-MRSA 923 in a Murine Blood Infection
2.2. The saeRS Single Knockout and coa/saeRS Double Knockout Significantly Eliminated the Lethality of CA-MRSA 923 in a Murine Blood Infection
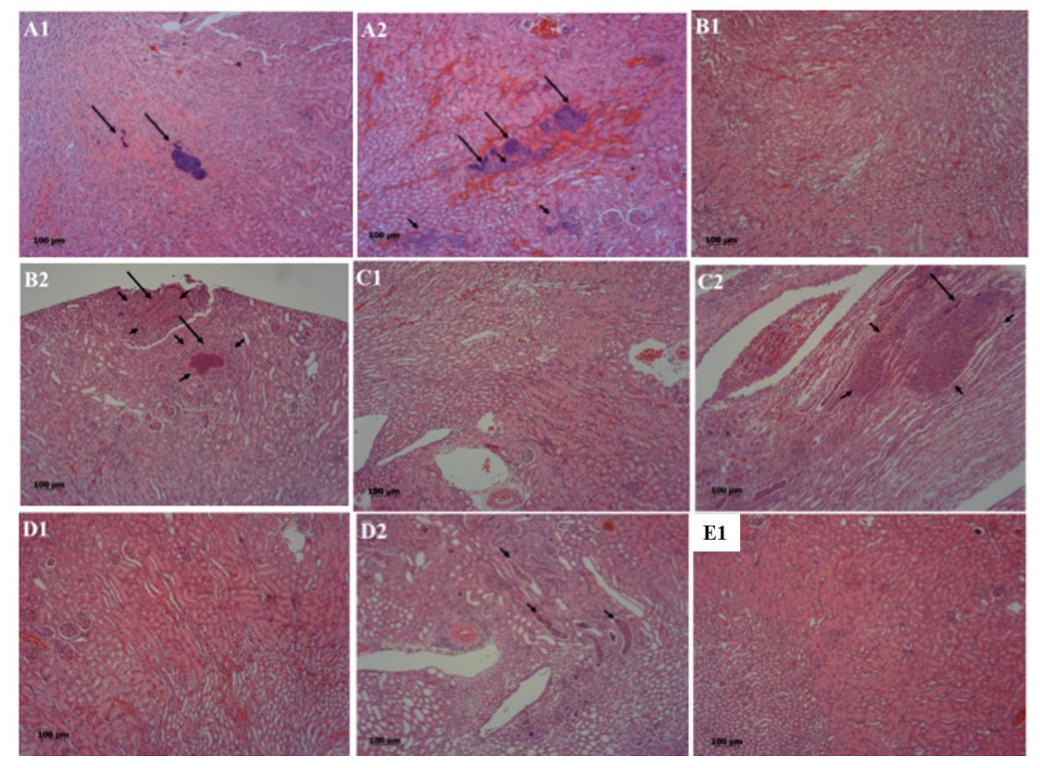
2.3. The Deletion Mutation of saeRS or coa/saeRS Remarkably Diminished the Histologically Pathological Damages in the Kidneys of Mice Infected with CA-MRSA 923
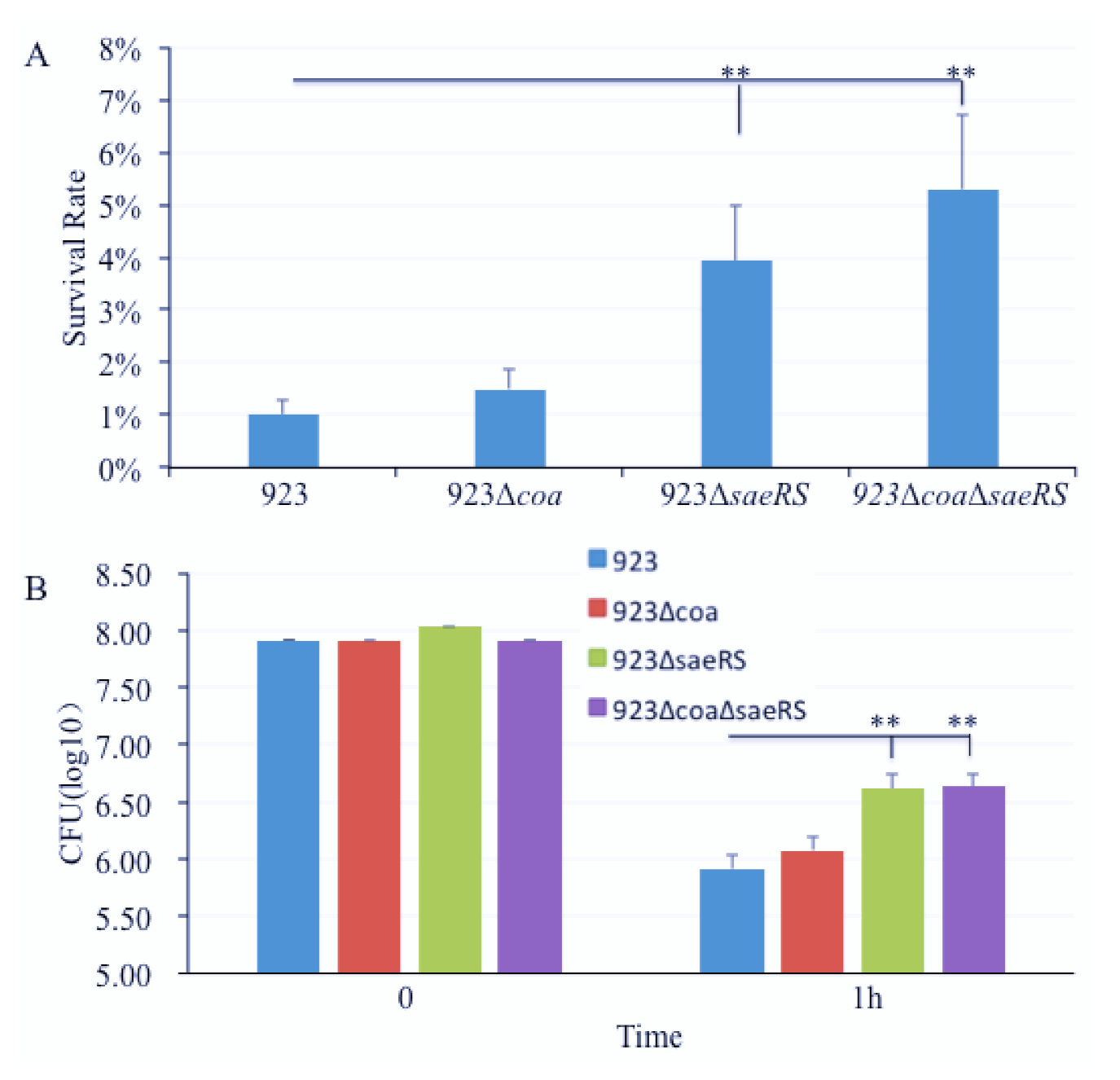
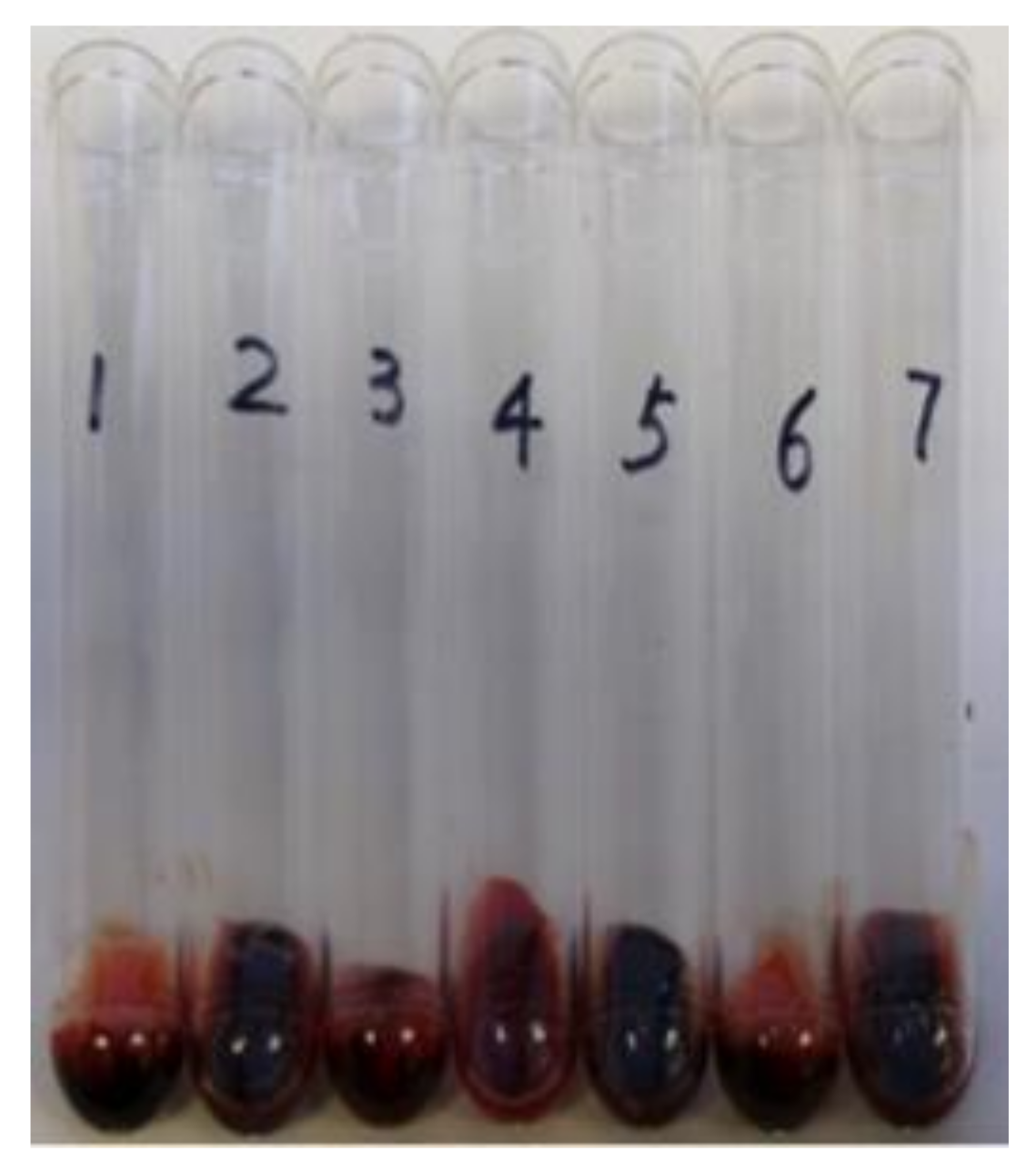
2.4. The Deletion Mutation of saeRS Significantly Increased, but Coagulase Null Mutation Had No Influence on the Ability of CA-MRSA 923 to Survive in Mouse Blood In Vitro
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Media
4.2. S. aureus Survival and Blood Coagulation Assays
4.3. Mouse Intravenous Infection Model and Histopathology
4.4. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klevens, R.M.; Morrison, M.A.; Nadle, J.; Petit, S.; Gershman, K.; Ray, S.; Harrison, L.H.; Lynfield, R.; Dumyati, G.; Townes, J.M.; et al. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA 2007, 298, 1763–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, R.J.; Lowy, F.D. Pathogenesis of methicillin-resistant Staphylococcus aureus infection. Clin. Infect Dis. 2008, 46, S350–S359. [Google Scholar] [CrossRef] [Green Version]
- Sowash, M.G.; Uhlemann, A.-C. Community-associated methicillin-resistant Staphylococcus aureus case studies. Methods Mol. Biol. 2013, 1085, 25–69. [Google Scholar] [CrossRef] [Green Version]
- Chalmers, S.J.; Wylam, M.E. Methicillin-Resistant Staphylococcus aureus Infection and Treatment Options. Methods Mol. Biol. 2019, 2069, 229–251. [Google Scholar] [CrossRef]
- Foster, T.J.; Höök, M. Surface protein adhesins of Staphylococcus aureus. Trends Microbiol. 1998, 6, 484–488. [Google Scholar] [CrossRef]
- Liu, G.Y.; Essex, A.; Buchanan, J.T.; Datta, V.; Hoffman, H.M.; Bastian, J.F.; Fierer, J.; Nizet, V. Staphylococcus aureus golden pigment impairs neutrophil killing and promotes virulence through its antioxidant activity. J. Exp. Med. 2005, 202, 209–215. [Google Scholar] [CrossRef]
- Clauditz, A.; Resch, A.; Wieland, K.-P.; Peschel, A.; Götz, F. Staphyloxanthin Plays a Role in the Fitness of Staphylococcus aureus and Its Ability to Cope with Oxidative Stress. Infect. Immun. 2006, 74, 4950–4953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novick, R.P. Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol. Microbiol. 2003, 48, 1429–1449. [Google Scholar] [CrossRef]
- Montgomery, C.P.; Boyle-Vavra, S.; Daum, R.S. Importance of the Global Regulators Agr and SaeRS in the Pathogenesis of CA-MRSA USA300 Infection. PLoS ONE 2010, 5, e15177. [Google Scholar] [CrossRef] [Green Version]
- Thoendel, M.; Horswill, A.R. Random mutagenesis and topology analysis of the autoinducing peptide biosynthesis proteins in Staphylococcus aureus. Mol. Microbiol. 2012, 87, 318–337. [Google Scholar] [CrossRef] [Green Version]
- Giraudo, A.T.; Calzolari, A.; Cataldi, A.A.; Bogni, C.; Nagel, R. The sae locus of Staphylococcus aureus encodes a two-component regulatory system. FEMS Microbiol. Lett. 1999, 177, 15–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Yu, C.; Sun, J.; Liu, H.; Landwehr, C.; Holmes, D.; Ji, Y. Inactivation of a Two-Component Signal Transduction System, SaeRS, Eliminates Adherence and Attenuates Virulence of Staphylococcus aureus. Infect. Immun. 2006, 74, 4655–4665. [Google Scholar] [CrossRef] [Green Version]
- Voyich, J.M.; Vuong, C.; Dewald, M.; Nygaard, T.; Kocianova, S.; Griffith, S.; Jones, J.; Iverson, C.; Sturdevant, D.E.; Braughton, K.R.; et al. The SaeR/S Gene Regulatory System Is Essential for Innate Immune Evasion by Staphylococcus aureus. J. Infect. Dis. 2009, 199, 1698–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nygaard, T.; Pallister, K.B.; Ruzevich, P.; Griffith, S.; Vuong, C.; Voyich, J.M. SaeR Binds a Consensus Sequence within Virulence Gene Promoters to Advance USA300 Pathogenesis. J. Infect. Dis. 2010, 201, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Zurek, O.W.; Nygaard, T.K.; Watkins, R.L.; Pallister, K.B.; Torres, V.J.; Horswill, A.R.; Voyich, J.M. The role of innate immunity in promoting SaeR/S-mediated virulence in Staphylococcus aureus. J. Innate Immun. 2014, 6, 21–30. [Google Scholar] [CrossRef]
- Liu, Q.; Yeo, W.; Bae, T. The SaeRS Two-Component System of Staphylococcus aureus. Genes 2016, 7, 81. [Google Scholar] [CrossRef] [Green Version]
- Fournier, B.; Klier, A.; Rapoport, G. The two-component system ArlS-ArlR is a regulator of virulence gene expression in Staphylococcus aureus. Mol. Microbiol. 2001, 41, 247–261. [Google Scholar] [CrossRef]
- Bronner, S.; Monteil, H.; Prévost, G. Regulation of virulence determinants in Staphylococcus aureus: Complexity and applications. FEMS Microbiol. Rev. 2004, 28, 183–200. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Zheng, L.; Landwehr, C.; Lunsford, D.; Holmes, D.; Ji, Y. Global Regulation of Gene Expression by ArlRS, a Two-Component Signal Transduction Regulatory System of Staphylococcus aureus. J. Bacteriol. 2005, 187, 5486–5492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, Y.-T.; Manna, A.C.; Projan, S.J.; Cheung, A.L. SarA, a Global Regulator of Virulence Determinants in Staphylococcus aureus, Binds to a Conserved Motif Essential for sar-dependent Gene Regulation. J. Biol. Chem. 1999, 274, 37169–37176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, A.L.; Nishina, K.A.; Trotonda, M.P.; Tamber, S. The SarA protein family of Staphylococcus aureus. Int. J. Biochem. Cell Biol. 2008, 40, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Loughran, A.J.; Gaddy, D.; Beenken, K.E.; Meeker, D.G.; Morello, R.; Zhao, H.; Byrum, S.D.; Tackett, A.J.; Cassat, J.E.; Smeltzer, M.S. Impact of sarA and Phenol-Soluble Modulins on the Pathogenesis of Osteomyelitis in Diverse Clinical Isolates of Staphylococcus aureus. Infect. Immun. 2016, 84, 2586–2594. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Cheung, A.; Bayer, A.S.; Chen, L.; Abdelhady, W.; Kreiswirth, B.N.; Yeaman, M.R.; Xiong, Y.Q. The global regulon sarA regulates β-Lactam antibiotic resistance in methicillin-resistant Staphylococcus aureus in vitro and in endovascular infections. J. Infect. Dis. 2016, 214, 1421–1429. [Google Scholar] [CrossRef] [Green Version]
- Ballal, A.; Ray, B.; Manna, A.C. sarZ, a sarA Family Gene, Is Transcriptionally Activated by MgrA and Is Involved in the Regulation of Genes Encoding Exoproteins in Staphylococcus aureus. J. Bacteriol. 2009, 191, 1656–1665. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.K.; Luong, T.T.; Lee, C.Y. RNAIII of the Staphylococcus aureus agr system activates global regulator MgrA by stabilizing mRNA. Proc. Natl. Acad. Sci. USA 2015, 112, 14036–14041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goerke, C.; Fluckiger, U.; Steinhuber, A.; Bisanzio, V.; Ulrich, M.; Bischoff, M.; Patti, J.M.; Wolz, C. The role of Staphylococcus aureus global regulators sae and σB in virulence gene expression during device-related infection. Infect. Immun. 2005, 73, 3415–3421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goerke, C.; Fluckiger, U.; Steinhuber, A.; Zimmerli, W.; Wolz, C. Impact of the regulatory loci agr, sarA and sae of Staphylococcus aureus on the induction of α-toxin during device-related infection resolved by direct quantitative transcript analysis. Mol. Microbiol. 2001, 40, 1439–1447. [Google Scholar] [CrossRef] [Green Version]
- Steinhuber, A.; Goerke, C.; Bayer, M.G.; Döring, G.; Wolz, C. Molecular Architecture of the Regulatory Locus sae of Staphylococcus aureus and Its Impact on Expression of Virulence Factors. J. Bacteriol. 2003, 185, 6278–6286. [Google Scholar] [CrossRef] [Green Version]
- Rogasch, K.; Rühmling, V.; Pané-Farré, J.; Höper, D.; Weinberg, C.; Fuchs, S.; Schmudde, M.; Bröker, B.; Wolz, C.; Hecker, M.; et al. Influence of the Two-Component System SaeRS on Global Gene Expression in Two Different Staphylococcus aureus Strains. J. Bacteriol. 2006, 188, 7742–7758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.; Jeong, D.-W.; Liu, Q.; Yeo, W.-S.; Vogl, T.; Skaar, E.P.; Chazin, W.J.; Bae, T. Calprotectin Increases the Activity of the SaeRS Two Component System and Murine Mortality during Staphylococcus aureus Infections. PLoS Pathog. 2015, 11, e1005026. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Cheng, B.L.; Boyle-Vavra, S.; Alegre, M.-L.; Daum, R.S.; Chong, A.S.; Montgomery, C.P. Proteomic Identification of saeRS -Dependent Targets Critical for Protective Humoral Immunity against Staphylococcus aureus Skin Infection. Infect. Immun. 2015, 83, 3712–3721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baddour, L.M.; Tayidi, M.M.; Walker, E.; McDevitt, D.; Foster, T.J. Virulence of coagulase-deficient mutants of Staphylococcus aureus in experimental endocarditis. J. Med. Microbiol. 1994, 41, 259–263. [Google Scholar] [CrossRef]
- Moreillon, P.; Entenza, J.M.; Francioli, P.; McDevitt, D.; Foster, T.J.; Francois, P.; Vaudaux, P. Role of Staphylococcus aureus coagulase and clumping factor in pathogenesis of experimental endocarditis. Infect. Immun. 1995, 63, 4738–4743. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.G.; McAdow, M.; Kim, H.K.; Bae, T.; Missiakas, D.M.; Schneewind, O. Contribution of Coagulases towards Staphylococcus aureus Disease and Protective Immunity. PLoS Pathog. 2010, 6, e1001036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Hall, J.; Yang, J.; Ji, Y. The SaeRS Two-Component System Controls Survival of Staphylococcus aureus in Human Blood through Regulation of Coagulase. Front. Cell. Infect. Microbiol. 2017, 7, 204. [Google Scholar] [CrossRef] [Green Version]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-negative staphylococci. Clin. Microbiol. Rev. 2014, 27, 870–926. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Bi, C.; Wang, T.; Xiang, H.; Chen, F.; Hu, J.; Liu, B.; Cai, H.; Zhong, X.; Deng, X.; et al. A coagulase-negative and non-haemolytic strain of Staphylococcus aureus for investigating the roles of SrtA in a murine model of bloodstream infection. Pathog. Dis. 2015, 73, ftv042. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, D.; Lâm, T.-T.; Geiger, T.; Mainiero, M.; Engelmann, S.; Hussain, M.; Bosserhoff, A.; Frosch, M.; Bischoff, M.; Wolz, C.; et al. A Point Mutation in the Sensor Histidine Kinase SaeS of Staphylococcus aureus Strain Newman Alters the Response to Biocide Exposure. J. Bacteriol. 2009, 191, 7306–7314. [Google Scholar] [CrossRef] [Green Version]
- Lei, T.; Zhang, Y.; Yang, J.; Silverstein, K.; Ji, Y. Complete Genome Sequence of Hospital-Acquired Methicillin-Resistant Staphylococcus aureus Strain WCUH29. Microbiol. Resour. Announc. 2019, 8, e00551-19. [Google Scholar] [CrossRef] [Green Version]
- Thomer, L.; Schneewind, O.; Missiakas, D. Multiple Ligands of von Willebrand Factor-binding Protein (vWbp) Promote Staphylococcus aureus Clot Formation in Human Plasma. J. Biol. Chem. 2013, 288, 28283–28292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janhsen, W.K.; Arnold, C.; Hentschel, J.; Lehmann, T.; Pfister, W.; Baier, M.; Böer, K.; Hünniger, K.; Kurzai, O.; Hipler, U.-C.; et al. Colonization of CF patients’ upper airways with S. aureus contributes more decisively to upper airway inflammation than P. aeruginosa. Med. Microbiol. Immunol. 2016, 205, 485–500. [Google Scholar] [CrossRef]
- Okumura, C.Y.; Nizet, V. Subterfuge and Sabotage: Evasion of Host Innate Defenses by Invasive Gram-Positive Bacterial Pathogens. Annu. Rev. Microbiol. 2014, 68, 439–458. [Google Scholar] [CrossRef] [Green Version]
- McGuinness, W.A.; Kobayashi, S.D.; DeLeo, F.R. Evasion of Neutrophil Killing by Staphylococcus aureus. Pathogens 2016, 5, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.; Marra, A.; Rosenberg, M.; Woodnutt, G. Regulated Antisense RNA Eliminates Alpha-Toxin Virulence in Staphylococcus aureus Infection. J. Bacteriol. 1999, 181, 6585–6590. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.G.; DeDent, A.C.; Schneewind, O.; Missiakas, D. A play in four acts: Staphylococcus aureus abscess formation. Trends Microbiol. 2011, 19, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Fowler, T.; Wann, E.R.; Joh, D.; Johansson, S.; Foster, T.J.; Höök, M. Cellular invasion by Staphylococcus aureus involves a fibronectin bridge between the bacterial fibronectin-binding MSCRAMMs and host cell β1 integrins. Eur. J. Cell Biol. 2000, 79, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Li, C.; Jeong, D.-W.; Sohn, C.; He, C.; Bae, T. In the Staphylococcus aureus Two-Component System sae, the Response Regulator SaeR Binds to a Direct Repeat Sequence and DNA Binding Requires Phosphorylation by the Sensor Kinase SaeS. J. Bacteriol. 2010, 192, 2111–2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kernodle, D.S.; Voladri, R.K.; EMenzies, B.; Hager, C.C.; Edwards, K.M. Expression of an antisense hla fragment in Staphylococcus aureus reduces alpha-toxin production in vitro and attenuates lethal activity in a murine model. Infect. Immun. 1997, 65, 179–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Ji, Y. Involvement of alpha5beta1-integrin and TNF-alpha in Staphylococcus aureus alpha-toxin-induced death of epithelial cells. Cell Microbiol. 2007, 9, 1809–1818. [Google Scholar] [CrossRef]
- Hall, J.; Yang, J.; Guo, H.; Ji, Y. The AirSR two-component system contributes to Staphylococcus aureus survival in human blood and transcriptionally regulates sspABC operon. Front. Microbiol. 2015, 6, 682. [Google Scholar] [CrossRef]
- Hall, J.W.; Yang, J.; Guo, H.; Ji, Y. The Staphylococcus aureus AirSR Two-Component System Mediates Reactive Oxygen Species Resistance via Transcriptional Regulation of Staphyloxanthin Production. Infect. Immun. 2017, 85, e00838-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Number of Mice in Trial I | Number of Mice in Trial II |
|---|---|---|
| 923 | 6 | 5 |
| 923Δcoa | 6 | 5 |
| 923ΔsaeRS | 6 | 5 |
| 923ΔcoaΔsaeRS | 6 | 5 |
| Negative control (PBS) | 6 | 5 |
| Strain | Relevant Characteristics | Reference |
|---|---|---|
| 923 | USA300 CA-MRSA | [9] |
| 923ΔsaeRS | 923 saeRS deletion mutant | [35] |
| 923Δcoa | 923 coa deletion mutant | [35] |
| 923ΔcoaΔsaeRS | 923 coa and saeRS double deletion mutant | [35] |
| 923/pYH4 | 923 carrying plasmid pYH4, ErmR | [35] |
| 923Δcoa/pYH4 | 923 coa deletion mutant with empty plasmid | [35] |
| 923Δcoa/pYH4-coa | coa complementary strain, ErmR | [35] |
| 923ΔsaeRS/pYH4 | 923 saeRS deletion mutant with empty plasmid | [35] |
| 923ΔsaeRS/pYH4-coa | ErmR | [35] |
| 923ΔcoaΔsaeRS/pYH4-coa | ErmR | [35] |
| 923ΔcoaΔsaeRS/pYH4-saeRS | ErmR | [35] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Gao, W.; Yang, J.; Guo, H.; Zhang, J.; Ji, Y. Contribution of Coagulase and Its Regulator SaeRS to Lethality of CA-MRSA 923 Bacteremia. Pathogens 2021, 10, 1396. https://doi.org/10.3390/pathogens10111396
Liu Y, Gao W, Yang J, Guo H, Zhang J, Ji Y. Contribution of Coagulase and Its Regulator SaeRS to Lethality of CA-MRSA 923 Bacteremia. Pathogens. 2021; 10(11):1396. https://doi.org/10.3390/pathogens10111396
Chicago/Turabian StyleLiu, Ying, Wei Gao, Junshu Yang, Haiyong Guo, Jiang Zhang, and Yinduo Ji. 2021. "Contribution of Coagulase and Its Regulator SaeRS to Lethality of CA-MRSA 923 Bacteremia" Pathogens 10, no. 11: 1396. https://doi.org/10.3390/pathogens10111396