
Chlamydia pecorum Ovine Abortion: Associations between Maternal Infection and Perinatal Mortality

 and
and
Abstract
:1. Introduction
2. Results
2.1. Prevalence of C. pecorum
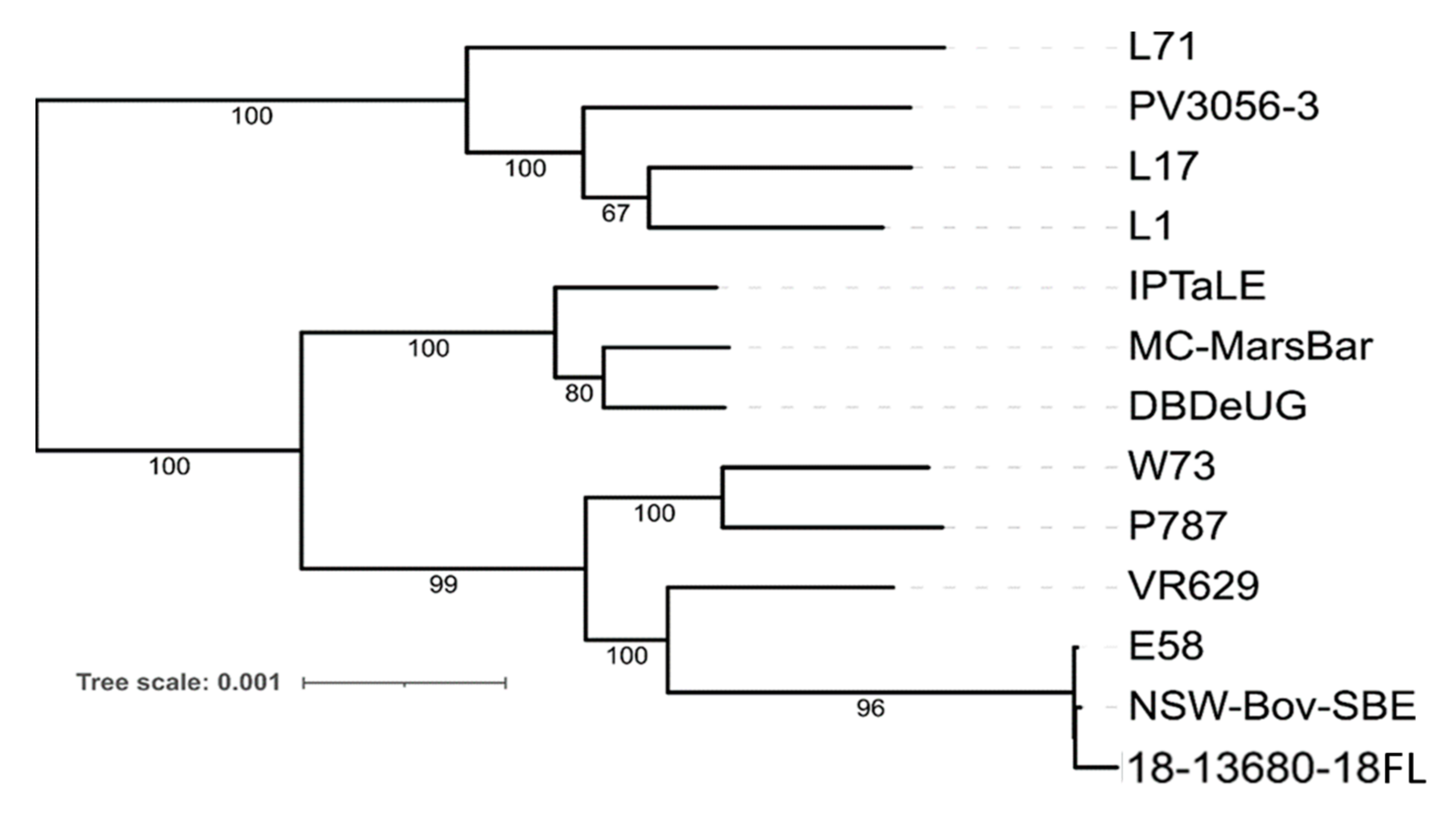
2.2. Whole Genome Analysis of Ovine Abortion Strain 18-13680-18FL
2.3. MLST of C. pecorum from Vaginal, Rectal and Conjunctival Sites
3. Discussion
4. Materials and Methods
4.1. Farm History and Sample Collection
4.2. Serological Testing
4.3. Quantitative PCR (qPCR)
4.4. Statistical Analysis
4.5. Multilocus Sequence Typing (MLST)
4.6. Whole Genome Sequencing (WGS)
4.7. Genome Assembly and Analysis
4.8. Detection of the C. pecorum Plasmid
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sánchez Romano, J.; Leijon, M.; Hagström, Å.; Jinnerot, T.; Rockström, U.K.; Tryland, M. Chlamydia pecorum associated with an outbreak of infectious keratoconjunctivitis in semi-domesticated reindeer in Sweden. Front. Vet. Sci. 2019, 6, 14. [Google Scholar] [CrossRef]
- Jelocnik, M.; Self, R.; Timms, P.; Borel, N.; Polkinghorne, A. Novel sequence types of Chlamydia pecorum infect free-ranging Alpine ibex (Capra ibex) and red deer (Cervus elaphus) in Switzerland. J. Wildl. Dis. 2015, 51, 479–483. [Google Scholar] [CrossRef] [Green Version]
- Cheong, H.C.; Lee, C.Y.Q.; Cheok, Y.Y.; Tan, G.M.Y.; Looi, C.Y.; Wong, W.F. Chlamydiaceae: Diseases in primary hosts and zoonosis. Microorganisms 2019, 7, 146. [Google Scholar] [CrossRef] [Green Version]
- Jelocnik, M.; Frentiu, F.D.; Timms, P.; Polkinghorne, A. Multilocus sequence analysis provides insights into molecular epidemiology of Chlamydia pecorum infections in australian sheep, cattle, and koalas. J. Clin. Microbiol. 2013, 51, 2625–2632. [Google Scholar] [CrossRef] [Green Version]
- Burnard, D.; Polkinghorne, A. Chlamydial infections in wildlife-conservation threats and/or reservoirs of ‘spill-over’ infections? Vet. Microbiol. 2016, 196, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Walker, E.; Lee, E.J.; Timms, P.; Polkinghorne, A. Chlamydia pecorum infections in sheep and cattle: A common and under-recognised infectious disease with significant impact on animal health. Vet. J. 2015, 206, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Poudel, A.; Elsasser, T.H.; Rahman, K.S.; Chowdhury, E.U.; Kaltenboeck, B. Asymptomatic endemic Chlamydia pecorum infections reduce growth rates in calves by up to 48 percent. PLoS ONE 2012, 7, e44961. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Jacobson, C.; Gardner, G.; Carmichael, I.; Campbell, A.J.D.; Ryan, U. Longitudinal prevalence and faecal shedding of Chlamydia pecorum in sheep. Vet. J. 2014, 201, 322–326. [Google Scholar] [CrossRef] [Green Version]
- Clarkson, M.J.; Philips, H.L. Isolation of faecal chlamydia from sheep in Britain andtheir characterization by cultural properties. Vet. J. 1997, 153, 307–310. [Google Scholar] [CrossRef]
- Lenzko, H.; Moog, U.; Henning, K.; Lederbach, R.; Diller, R.; Menge, C.; Sachse, K.; Sprague, L.D. High frequency of chlamydial co-infections in clinically healthy sheep flocks. BMC Vet. Res. 2011, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Philips, H.L.; Clarkson, M.J. Experimental infection of pregnant ewes with Chlamydia pecorum. Infect. Immun. 1998, 66, 2818–2821. [Google Scholar] [CrossRef] [Green Version]
- Westermann, T.; Jenkins, C.; Onizawa, E.; Gestier, S.; McNally, J.; Kirkland, P.; Zhang, J.; Bogema, D.; Manning, L.K.; Walker, K.; et al. Chlamydia pecorum–Associated sporadic ovine abortion. Vet. Pathol. 2021, 58, 114–122. [Google Scholar] [CrossRef]
- Essig, A.; Longbottom, D. Chlamydia abortus: New aspects of infectious abortion in sheep and potential risk for pregnant women. Curr. Clin. Microbiol. Rep. 2015, 2, 22–34. [Google Scholar] [CrossRef] [Green Version]
- Jelocnik, M.; Bachmann, N.L.; Kaltenboeck, B.; Waugh, C.; Woolford, L.; Speight, K.N.; Gillett, A.; Higgins, D.P.; Flanagan, C.; Myers, G.S.A.; et al. Genetic diversity in the plasticity zone and the presence of the chlamydial plasmid differentiates Chlamydia pecorum strains from pigs, sheep, cattle, and koalas. BMC Genom. 2015, 16, 893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jelocnik, M.; Bachmann, N.L.; Seth-Smith, H.; Thomson, N.R.; Timms, P.; Polkinghorne, A.M. Molecular characterisation of the Chlamydia pecorum plasmid from porcine, ovine, bovine, and koala strains indicates plasmid-strain co-evolution. PeerJ 2016, 4, e1661. [Google Scholar] [CrossRef] [PubMed]
- Sait, M.; Livingstone, M.; Clark, E.M.; Wheelhouse, N.; Spalding, L.; Markey, B.; Magnino, S.; Lainson, F.A.; Myers, G.S.; Longbottom, D. Genome sequencing and comparative analysis of three Chlamydia pecorum strains associated with different pathogenic outcomes. BMC Genom. 2014, 15, 23. [Google Scholar] [CrossRef] [Green Version]
- Rekiki, A.; Bouakane, A.; Hammami, S.; El Idrissi, A.H.; Bernard, F.; Rodolakis, A. Efficacy of live Chlamydophila abortus vaccine 1B in protecting mice placentas and foetuses against strains of Chlamydophila pecorum isolated from cases of abortion. Vet. Microbiol. 2004, 99, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Berri, M.; Rekiki, A.; Boumedine, K.S.; Rodolakis, A. Simultaneous differential detection of Chlamydophila abortus, Chlamydophila pecorum and Coxiella burnetii from aborted ruminant’s clinical samples using multiplex PCR. BMC Microbiol. 2009, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Giannitti, F.; Anderson, M.; Miller, M.; Rowe, J.; Sverlow, K.; Vasquez, M.; Canton, G. Chlamydia pecorum: Fetal and placental lesions in sporadic caprine abortion. J. Vet. Diagn. Investig. 2016, 28, 184–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clune, T.; Besier, S.; Hair, S.; Hancock, S.; Lockwood, A.; Thompson, A.; Jelocnik, M.; Jacobson, C. Chlamydia pecorum detection in aborted and stillborn lambs from Western Australia. Vet. Res. 2021, 52, 84. [Google Scholar] [CrossRef]
- Walker, E.; Moore, C.; Shearer, P.; Jelocnik, M.; Bommana, S.; Timms, P.; Polkinghorne, A. Clinical, diagnostic and pathologic features of presumptive cases of Chlamydia pecorum-associated arthritis in Australian sheep flocks. BMC Vet. Res. 2016, 12, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bommana, S.; Walker, E.; Desclozeaux, M.; Jelocnik, M.; Timms, P.; Polkinghorne, A.; Carver, S. Molecular and serological dynamics of Chlamydia pecorum infection in a longitudinal study of prime lamb production. PeerJ 2018, 6, e4296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polkinghorne, A.; Borel, N.; Becker, A.; Lu, Z.H.; Zimmermann, D.R.; Brugnera, E.; Pospischil, A.; Vaughan, L. Molecular evidence for chlamydial infections in the eyes of sheep. Vet. Microbiol. 2009, 135, 142–146. [Google Scholar] [CrossRef]
- Jelocnik, M.; Walker, E.; Pannekoek, Y.; Ellem, J.; Timms, P.; Polkinghorne, A. Evaluation of the relationship between Chlamydia pecorum sequence types and disease using a species-specific multi-locus sequence typing scheme (MLST). Vet. Microbiol. 2014, 174, 214–222. [Google Scholar] [CrossRef]
- Mygind, P.H.; Christiansen, G.; Roepstorff, P.; Birkelund, S. Membrane proteins PmpG and PmpH are major constituents of Chlamydia trachomatis L2 outer membrane complex. FEMS Microbiol. Lett. 2000, 186, 163–169. [Google Scholar] [CrossRef]
- Becker, E.; Hegemann, J.H. All subtypes of the Pmp adhesin family are implicated in chlamydial virulence and show species-specific function. Microbiologyopen 2014, 3, 544–556. [Google Scholar] [CrossRef]
- Molleken, K.; Schmidt, E.; Hegemann, J.H. Members of the Pmp protein family of Chlamydia pneumoniae mediate adhesion to human cells via short repetitive peptide motifs. Mol. Microbiol. 2010, 78, 1004–1017. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Pais, R.; Ohandjo, A.; He, C.; He, Q.; Omosun, Y.; Igietseme, J.U.; Eko, F.O. Comparative evaluation of the protective efficacy of two formulations of a recombinant Chlamydia abortus subunit candidate vaccine in a mouse model. Vaccine 2015, 33, 1865–1872. [Google Scholar] [CrossRef] [Green Version]
- Gomes, J.P.; Nunes, A.; Bruno, W.J.; Borrego, M.J.; Florindo, C.; Dean, D. Polymorphisms in the nine polymorphic membrane proteins of Chlamydia trachomatis across all serovars: Evidence for serovar Da recombination and correlation with tissue tropism. J. Bacteriol. 2006, 188, 275–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karunakaran, K.P.; Yu, H.; Jiang, X.; Chan, Q.; Moon, K.M.; Foster, L.J.; Brunham, R.C. Outer membrane proteins preferentially load MHC class II peptides: Implications for a Chlamydia trachomatis T cell vaccine. Vaccine 2015, 33, 2159–2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longbottom, D.; Russell, M.; Jones, G.E.; Lainson, F.A.; Herring, A.J. Identification of a multigene family coding for the 90 kDa proteins of the ovine abortion subtype of Chlamydia psittaci. FEMS Microbiol. Lett. 1996, 142, 277–281. [Google Scholar] [CrossRef]
- Johnson, R.M.; Yu, H.; Kerr, M.S.; Slaven, J.E.; Karunakaran, K.P.; Brunham, R.C. PmpG303-311, a protective vaccine epitope that elicits persistent cellular immune responses in Chlamydia muridarum-immune mice. Infect. Immun. 2012, 80, 2204–2211. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Liu, G.; Kickhoefer, V.A.; Rome, L.H.; Li, L.X.; McSorley, S.J.; Kelly, K.A. A protective vaccine against Chlamydia genital infection using vault nanoparticles without an added adjuvant. Vaccines 2017, 5, 3. [Google Scholar] [CrossRef]
- Nunes, A.; Gomes, J.P.; Karunakaran, K.P.; Brunham, R.C. Bioinformatic analysis of Chlamydia trachomatis polymorphic membrane proteins PmpE, PmpF, PmpG and PmpH as potential vaccine antigens. PLoS ONE 2015, 10, e0131695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desclozeaux, M.; Robbins, A.; Jelocnik, M.; Khan, S.A.; Hanger, J.; Gerdts, V.; Potter, A.; Polkinghorne, A.; Timms, P. Immunization of a wild koala population with a recombinant Chlamydia pecorum Major Outer Membrane Protein (MOMP) or Polymorphic Membrane Protein (PMP) based vaccine: New insights into immune response, protection and clearance. PLoS ONE 2017, 12, e0178786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Favaroni, A.; Hegemann, J.H. Chlamydia trachomatis polymorphic membrane Proteins (Pmps) form functional homomeric and heteromeric oligomers. Front. Microbiol. 2021, 12, 709724. [Google Scholar] [CrossRef]
- Ntountoumi, C.; Vlastaridis, P.; Mossialos, D.; Stathopoulos, C.; Iliopoulos, I.; Promponas, V.; Oliver, S.G.; Amoutzias, G.D. Low complexity regions in the proteins of prokaryotes perform important functional roles and are highly conserved. Nucleic Acids Res. 2019, 47, 9998–10009. [Google Scholar] [CrossRef] [PubMed]
- Watt, B.; McNally, J.; Bourke, J.; Cavanagh, C.; Masters, A.; McCarthy, T.; Jenkins, C.; Staples, P. Chlamydia pecorum abortion in five widely dispersed NSW ewe flocks. Flock Herd Case Notes 2021, 9, 20–23. Available online: http://www.flockandherd.net.au/sheep/ireader/chlamydial-abortion.html (accessed on 21 September 2021).
- Thomas, N.S.; Lusher, M.; Storey, C.C.; Clarke, I.N. Plasmid diversity in Chlamydia. Microbiology 1997, 143 Pt 6, 1847–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escobedo-Guerra, M.R.; Katoku-Herrera, M.; Lopez-Hurtado, M.; Villagrana-Zesati, J.R.; de Haro-Cruz, M.J.; Guerra-Infante, F.M. Identification of a new variant of Chlamydia trachomatis in Mexico. Enferm. Infecc. Microbiol. Clin. 2019, 37, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Seth-Smith, H.M.B.; Harris, S.R.; Persson, K.; Marsh, P.; Barron, A.; Bignell, A.; Bjartling, C.; Clark, L.; Cutcliffe, L.T.; Lambden, P.R.; et al. Co-evolution of genomes and plasmids within Chlamydia trachomatis and the emergence in Sweden of a new variant strain. BMC Genom. 2009, 10, 239. [Google Scholar] [CrossRef] [Green Version]
- Szabo, K.V.; O’Neill, C.E.; Clarke, I.N. Diversity in Chlamydial plasmids. PLoS ONE 2020, 15, e0233298. [Google Scholar] [CrossRef]
- Herrmann, B. A new genetic variant of Chlamydia trachomatis. Sex Transm. Infect. 2007, 83, 253–254. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Lyu, Z.; Fan, Y.; Evans, C.R.; Barber, K.W.; Banerjee, K.; Igoshin, O.A.; Rinehart, J.; Ling, J. Metabolic stress promotes stop-codon readthrough and phenotypic heterogeneity. Proc. Natl. Acad. Sci. USA 2020, 117, 22167–22172. [Google Scholar] [CrossRef]
- Ehricht, R.; Slickers, S.; Goellner, S.; Hotzel, H.; Sachse, K. Optimized DNA microarray assay allows detection and genotyping of single PCR-amplifiable target copies. Mol. Cell. Probes 2006, 20, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Pantchev, A.; Sting, R.; Bauerfeind, R.; Tyczka, J.; Sachse, K. Detection of all Chlamydophila and Chlamydia spp. of veterinary interest using species-specific real-time PCR assays. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 473–484. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, J.; Croucher, N.J.; Goater, R.J.; Abudahab, K.; Aanensen, D.M.; Harris, S.R. Phandango: An interactive viewer for bacterial population genomics. Bioinformatics 2017, 34, 292–293. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 1 March 2010).
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.A.; Korobeynikov, A.; Lapidus, A.; Prjibelski, A.D.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling single-cell genomes and mini-metagenomes from chimeric MDA products. J. Comput. Biol. A J. Comput. Mol. Cell Biol. 2013, 20, 714–737. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Laetsch, D.R.; Blaxter, M.L. BlobTools: Interrogation of genome assemblies. F1000Research 2017, 6, 1287. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Marine, R.L.; Magaña, L.C.; Castro, C.J.; Zhao, K.; Montmayeur, A.M.; Schmidt, A.; Diez-Valcarce, M.; Ng, T.F.F.; Vinjé, J.; Burns, C.C.; et al. Comparison of illumina miseq and the ion torrent PGM and S5 platforms for whole-genome sequencing of picornaviruses and caliciviruses. J. Virol. Methods 2020, 280, 113865. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Cock, P.J.; Antao, T.; Chang, J.T.; Chapman, B.A.; Cox, C.J.; Dalke, A.; Friedberg, I.; Hamelryck, T.; Kauff, F.; Wilczynski, B.; et al. Biopython: Freely available Python tools for computational molecular biology and bioinformatics. Bioinformatics 2009, 25, 1422–1423. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef]
- Mai, U.; Sayyari, E.; Mirarab, S. Minimum variance rooting of phylogenetic trees and implications for species tree reconstruction. PLoS ONE 2017, 12, e0182238. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.; Robbins, A.; Loader, J.; Hanger, J.; Booth, R.; Jelocnik, M.; Polkinghorne, A.; Timms, P. Chlamydia pecorum gastrointestinal tract infection associations with urogenital tract infections in the koala (Phascolarctos cinereus). PLoS ONE 2018, 13, e0206471. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Host (ID) | Sample Type | PNM | ST | Reference | pCpec | CDS1 Deletion |
|---|---|---|---|---|---|---|---|
| 18-18549-63R | Ewe (7) | Rectal swab | + | 307 * | This study | + | − |
| 18-17161-75R | Ewe (24) | Rectal swab | + | 23 | This study | + | + |
| 18-18549-93R | Ewe (36) | Rectal swab | + | 305 * | This study | + | − |
| 18-18549-105R | Ewe (48) | Rectal swab | + | 206 | This study | + | − |
| 18-17161-147R | Ewe (25) | Rectal swab | − | 23 | This study | + | + |
| 18-17161-76V | Ewe (24) | Vaginal swab | + | 23 | This study | + | + |
| 18-18549-115V | Ewe (4) | Vaginal swab | + | 23 | This study | − | N/A |
| 18-18549-137V | Ewe (25) | Vaginal swab | − | 23 | This study | − | N/A |
| 18-18549-143V | Ewe (31) | Vaginal swab | − | 23 | This study | + | + |
| 18-17161-184V | Ewe (31) | Vaginal swab | − | 23 | This study | + | + |
| 18-18549-190C | Ewe (23) | Conjunctival swab | − | 23 | This study | + | + |
| 18-13680-29FM | Foetus 1 | Foetal Membranes | + | 23 | [12] | + | + |
| 18-13680-30FM | Foetus 2 | Foetal Membranes | + | 23 | [12] | + | + |
| 18-13680-22FK | Foetus 3 | Foetal Kidney | + | 23 | [12] | + | + |
| 18-13680-35FB | Foetus 3 | Foetal Brain | + | 23 | [12] | + | + |
| 18-13680-18FL | Foetus 1 | Foetal Lung Culture | + | 23 | [12] | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jenkins, C.; Jelocnik, M.; Onizawa, E.; McNally, J.; Coilparampil, R.; Pinczowski, P.; Bogema, D.; Westermann, T. Chlamydia pecorum Ovine Abortion: Associations between Maternal Infection and Perinatal Mortality. Pathogens 2021, 10, 1367. https://doi.org/10.3390/pathogens10111367
Jenkins C, Jelocnik M, Onizawa E, McNally J, Coilparampil R, Pinczowski P, Bogema D, Westermann T. Chlamydia pecorum Ovine Abortion: Associations between Maternal Infection and Perinatal Mortality. Pathogens. 2021; 10(11):1367. https://doi.org/10.3390/pathogens10111367
Chicago/Turabian StyleJenkins, Cheryl, Martina Jelocnik, Emily Onizawa, Justine McNally, Ronald Coilparampil, Pedro Pinczowski, Daniel Bogema, and Thomas Westermann. 2021. "Chlamydia pecorum Ovine Abortion: Associations between Maternal Infection and Perinatal Mortality" Pathogens 10, no. 11: 1367. https://doi.org/10.3390/pathogens10111367