Molecular Evidence of Novel Spotted Fever Group Rickettsia Species in Amblyomma albolimbatum Ticks from the Shingleback Skink (Tiliqua rugosa) in Southern Western Australia

Abstract
:1. Introduction
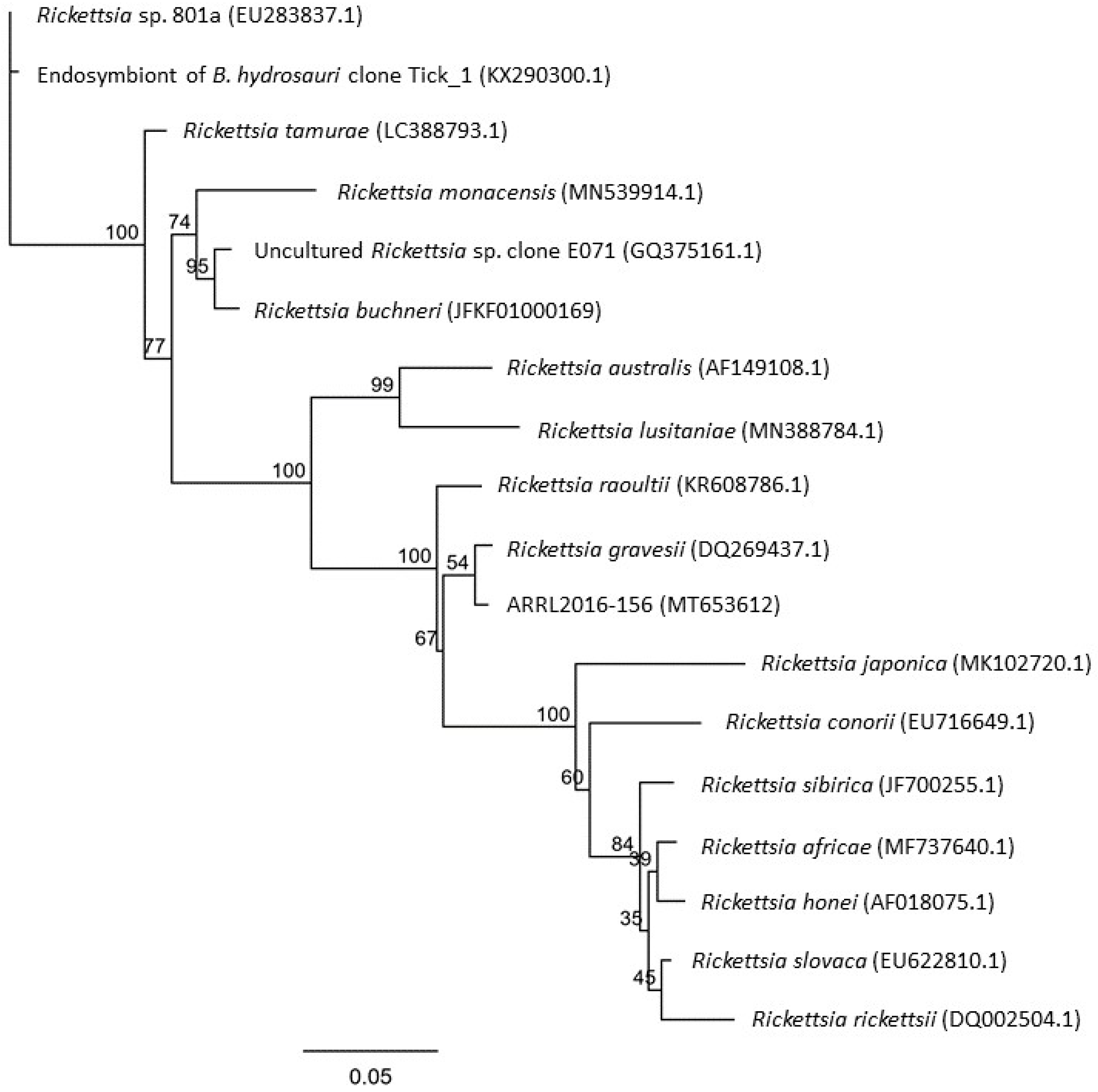
2. Results
3. Discussion
4. Materials and Methods
4.1. Tick Collection and Identification
4.2. DNA Extraction and Rickettsia-Specific qPCR Detection
4.3. Molecular Identification and Characterization of Detected Rickettsia spp.
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Madison-Antenucci, S.; Kramer, L.D.; Gebhardt, L.L.; Kauffman, E. Emerging Tick-Borne Diseases. Clin. Microbiol. Rev. 2020, 33. [Google Scholar] [CrossRef] [PubMed]
- Laroche, M.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.-E.; et al. Update on Tick-Borne Rickettsioses around the World: A Geographic Approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [Green Version]
- Barker, S.C.; Walker, A.R.; Campelo, D. A list of the 70 species of Australian ticks; diagnostic guides to and species accounts of Ixodes holocyclus (paralysis tick), Ixodes cornuatus (southern paralysis tick) and Rhipicephalus australis (Australian cattle tick); and consideration of the place of Australia in the evolution of ticks with comments on four controversial ideas. Int. J. Parasitol. 2014, 44, 941–953. [Google Scholar] [CrossRef] [PubMed]
- Graves, S.; Stenos, J. Tick-borne infectious diseases in Australia. Med. J. Aust. 2017, 206, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Andrew, R.; Bonnin, J.M.; Williams, S. Tick typhus in north queensland. Med. J. Aust. 1946, 2, 253–258. [Google Scholar] [CrossRef]
- Graves, S.R.; Stewart, L.; Stenos, J.; Stewart, R.S.; Schmidt, E.; Hudson, S.; Banks, J.; Huang, Z.; Dwyer, B. Spotted fever group rickettsial infection in South-Eastern Australia: Isolation of rickettsiae. Comp. Immunol. Microbiol. Infect. Dis. 1993, 16, 223–233. [Google Scholar] [CrossRef]
- Stewart, A.; Armstrong, M.; Graves, S.; Hajkowicz, K. Epidemiology and Characteristics of Rickettsia australis (Queensland Tick Typhus) Infection in Hospitalized Patients in North Brisbane, Australia. Trop. Med. Infect. Dis. 2017, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Unsworth, N.B.; Stenos, J.; Graves, S.R.; Faa, A.G.; Cox, G.E.; Dyer, J.R.; Boutlis, C.S.; Lane, A.M.; Shaw, M.D.; Robson, J.; et al. Flinders Island Spotted Fever Rickettsioses Caused by “marmionii” Strain of Rickettsia honei, Eastern Australia. Emerg. Infect. Dis. 2007, 13, 566–573. [Google Scholar] [CrossRef]
- Hone, F.S. A SERIES OF CASES CLOSELY RESEMBLING TYPHUS FEVER. Med. J. Aust. 1922, 1, 1–13. [Google Scholar] [CrossRef]
- Williams, M.; Izzard, L.; Graves, S.; Stenos, J.; Kelly, J.J. First probable Australian cases of human infection with Rickettsia felis (cat-flea typhus). Med. J. Aust. 2011, 194, 41–43. [Google Scholar] [CrossRef]
- Izzard, L.; Graves, S.; Cox, E.; Fenwick, S.; Unsworth, N.; Stenos, J. Novel Rickettsia in Ticks, Tasmania, Australia. Emerg. Infect. Dis. 2009, 15, 1654–1656. [Google Scholar] [CrossRef] [PubMed]
- Abdad, M.Y.; Abdallah, R.A.; El Karkouri, K.; Beye, M.; Stenos, J.; Owen, H.; Unsworth, N.; Robertson, I.D.; Blacksell, S.D.; Nguyen, T.-T.; et al. Rickettsia gravesii sp. nov.: A novel spotted fever group rickettsia in Western Australian Amblyomma triguttatum triguttatum ticks. Int. J. Syst. Evol. Microbiol. 2017, 67, 3156–3161. [Google Scholar] [CrossRef] [PubMed]
- Paris, D.H.; Dumler, J.S. State of the art of diagnosis of rickettsial diseases. Curr. Opin. Infect. Dis. 2016, 29, 433–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.T.; Satjanadumrong, J.; Hughes, T.; Stenos, J.; Blacksell, S.D. Diagnosis of spotted fever group Rickettsia infections: The Asian perspective. Epidemiol. Infect. 2019, 147, e286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilcins, I.-M.E.; Old, J.M.; Deane, E.M. Detection of a spotted fever group Rickettsia in the tick Ixodes tasmani collected from koalas in Port Macquarie, Australia. J. Med. Èntomol. 2008, 45, 745–750. [Google Scholar] [CrossRef]
- Stenos, J.; Walker, D.H.; Popov, V.L.; Graves, S. APONOMMA HYDROSAURI, THE REPTILE-ASSOCIATED TICK RESERVOIR OF RICKETTSIA HONEI ON FLINDERS ISLAND, AUSTRALIA. Am. J. Trop. Med. Hyg. 2003, 69, 314–317. [Google Scholar] [CrossRef]
- Vilcins, I.-M.E.; Fournier, P.-E.; Old, J.M.; Deane, E. Evidence for the Presence of Francisella and Spotted Fever Group Rickettsia DNA in the Tick Amblyomma fimbriatum (Acari: Ixodidae), Northern Territory, Australia. J. Med. Èntomol. 2009, 46, 926–933. [Google Scholar] [CrossRef]
- Whiley, H.; Custance, G.; Graves, S.; Stenos, J.; Taylor, M.J.; Ross, K.E.; Gardner, M.G. Rickettsia Detected in the Reptile Tick Bothriocroton hydrosauri from the Lizard Tiliqua rugosa in South Australia. Pathogens 2016, 5, 41. [Google Scholar] [CrossRef] [Green Version]
- Fournier, P.-E.; Dumler, J.S.; Greub, G.; Zhang, J.; Wu, Y.; Raoult, D. Gene Sequence-Based Criteria for Identification of New Rickettsia Isolates and Description of Rickettsia heilongjiangensis sp. nov. J. Clin. Microbiol. 2003, 41, 5456–5465. [Google Scholar] [CrossRef] [Green Version]
- Vayssier-Taussat, M.; Cosson, J.; Degeilh, B.; Eloit, M.; Fontanet, A.; Moutailler, S.; Raoult, D.; Sellal, E.; Ungeheuer, M.-N.; Zylbermann, P.; et al. How a multidisciplinary ‘One Health’ approach can combat the tick-borne pathogen threat in Europe. Future Microbiol. 2015, 10, 809–818. [Google Scholar] [CrossRef]
- Cabezas-Cruz, A.; Vayssier-Taussat, M.; Greub, G. Tick-borne pathogen detection: What’s new? Microbes Infect. 2018, 20, 441–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalada, M.J.; Stenos, J.; Vincent, G.; Barker, D.; Bradbury, R.S. A Molecular Survey of Tick-Borne Pathogens from Ticks Collected in Central Queensland, Australia. Vector-Borne Zoonotic Dis. 2018, 18, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Graves, S.; Jackson, C.; Hussain-Yusuf, H.; Vincent, G.; Nguyen, C.; Stenos, J.; Webster, M. Ixodes holocyclus Tick-Transmitted Human Pathogens in North-Eastern New South Wales, Australia. Trop. Med. Infect. Dis. 2016, 1, 4. [Google Scholar] [CrossRef] [PubMed]
- Hussain-Yusuf, H.; Stenos, J.; Vincent, G.; Shima, A.; Abell, S.; Preece, N.D.; Tadepalli, M.; Hii, S.F.; Bowie, N.; Mitram, K.; et al. Screening for Rickettsia, Coxiella and Borrelia Species in Ticks from Queensland, Australia. Pathogens 2020, 9, 1016. [Google Scholar] [CrossRef]
- Bonnet, S.I.; Binetruy, F.; Hernández-Jarguín, A.M.; Duron, O. The Tick Microbiome: Why Non-pathogenic Microorganisms Matter in Tick Biology and Pathogen Transmission. Front. Cell. Infect. Microbiol. 2017, 7, 236. [Google Scholar] [CrossRef]
- Weinert, L.A.; Araujo-Jnr, E.V.; Ahmed, M.Z.; Welch, J.J. The incidence of bacterial endosymbionts in terrestrial arthropods. Proc. R. Soc. Lond. 2015, 282, 20150249. [Google Scholar] [CrossRef]
- Barker, S.C.; Walker, A.R. Ticks of Australia. The species that infest domestic animals and humans. Zootaxa 2014, 3816, 1–144. [Google Scholar] [CrossRef]
- Roberts, F.H.S. Australian Ticks; Commonwealth Scientific and Industrial Research Organization: Melbourne, Victoria, Australia, 1970; p. 267.
- Stenos, J.; Unsworth, N.B.; Graves, S.R. A highly sensitive and specific real-time pcr assay for the detection of spotted fever and typhus group rickettsiae. Am. J. Trop. Med. Hyg. 2005, 73, 1083–1085. [Google Scholar] [CrossRef]
- Ishikura, M.; Ando, S.; Shinagawa, Y.; Matsuura, K.; Hasegawa, S.; Nakayama, T.; Fujita, H.; Watanabe, M. Phylogenetic Analysis of Spotted Fever Group Rickettsiae Based on gltA, 17-kDa, and rOmpA Genes Amplified by Nested PCR from Ticks in Japan. Microbiol. Immunol. 2003, 47, 823–832. [Google Scholar] [CrossRef] [Green Version]
- Roux, V.; Raoult, D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int. J. Syst. Evol. Microbiol. 2000, 50, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- Sekeyova, Z.; Roux, V.; Raoult, D. Phylogeny of Rickettsia spp. inferred by comparing sequences of ‘gene D’, which encodes an intracytoplasmic protein. Int. J. Syst. Evol. Microbiol. 2001, 51, 1353–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Locality/Coordinates | Life Cycle Stage | Ticks (n) | Rickettsia spp. gltA qPCR n (%) |
|---|---|---|---|
| Moulyinning (33.2290° S, 117.9377° E) | Adult (female) Adult (male) | 2 2 | 2 (100) 2 (100) |
| Wagin (33.3054° S, 117.3474° E) | Adult (male) | 3 | 3 (100) |
| Yanchep (31.5464° S, 115.6327° E) | Adult (female) Adult (male) Nymph | 3 4 1 | 3 (100) 4 (100) 1 (100) |
| Esperance (33.8608° S, 121.8896° E) | Adult (female) Adult (male) | 1 1 | 1 (100) 1 (100) |
| Southern Cross (31.2306° S, 119.3278° E) | Adult (female) Adult (male) Nymph | 6 11 1 | 6 (100) 9 (82) 1 (100) |
| Yellowdine (31.0656° S, 119.8116° E) | Adult (female) Adult (male) Nymph | 2 4 2 | 2 (100) 3 (75) 2 (100) |
| Tammin (31.6410° S, 117.4865° E) | Adult (female) Adult (male) | 1 5 | 1 (100) 4 (80) |
| Youndegin (31.8227° S, 117.2518° E) | Adult (female) Adult (male) | 1 2 | 1 (100) 2 (100) |
| Doodlakine (31.6020° S, 117.8976° E) | Adult (male) | 1 | 1 (100) |
| Korbel (31.6082° S, 118.1272° E) | Adult (female) Adult (male) | 5 6 | 5 (100) 5 (100) |
| Totals | 64 | 59 (92.2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tadepalli, M.; Vincent, G.; Hii, S.F.; Watharow, S.; Graves, S.; Stenos, J. Molecular Evidence of Novel Spotted Fever Group Rickettsia Species in Amblyomma albolimbatum Ticks from the Shingleback Skink (Tiliqua rugosa) in Southern Western Australia. Pathogens 2021, 10, 35. https://doi.org/10.3390/pathogens10010035
Tadepalli M, Vincent G, Hii SF, Watharow S, Graves S, Stenos J. Molecular Evidence of Novel Spotted Fever Group Rickettsia Species in Amblyomma albolimbatum Ticks from the Shingleback Skink (Tiliqua rugosa) in Southern Western Australia. Pathogens. 2021; 10(1):35. https://doi.org/10.3390/pathogens10010035
Chicago/Turabian StyleTadepalli, Mythili, Gemma Vincent, Sze Fui Hii, Simon Watharow, Stephen Graves, and John Stenos. 2021. "Molecular Evidence of Novel Spotted Fever Group Rickettsia Species in Amblyomma albolimbatum Ticks from the Shingleback Skink (Tiliqua rugosa) in Southern Western Australia" Pathogens 10, no. 1: 35. https://doi.org/10.3390/pathogens10010035