
A Mathematical Model for the Action Spectrum of Steady-State Pupil Size in Photopic Vision with Insight into Healthful Lighting



Abstract
:1. Introduction
2. Methods
2.1. Participants
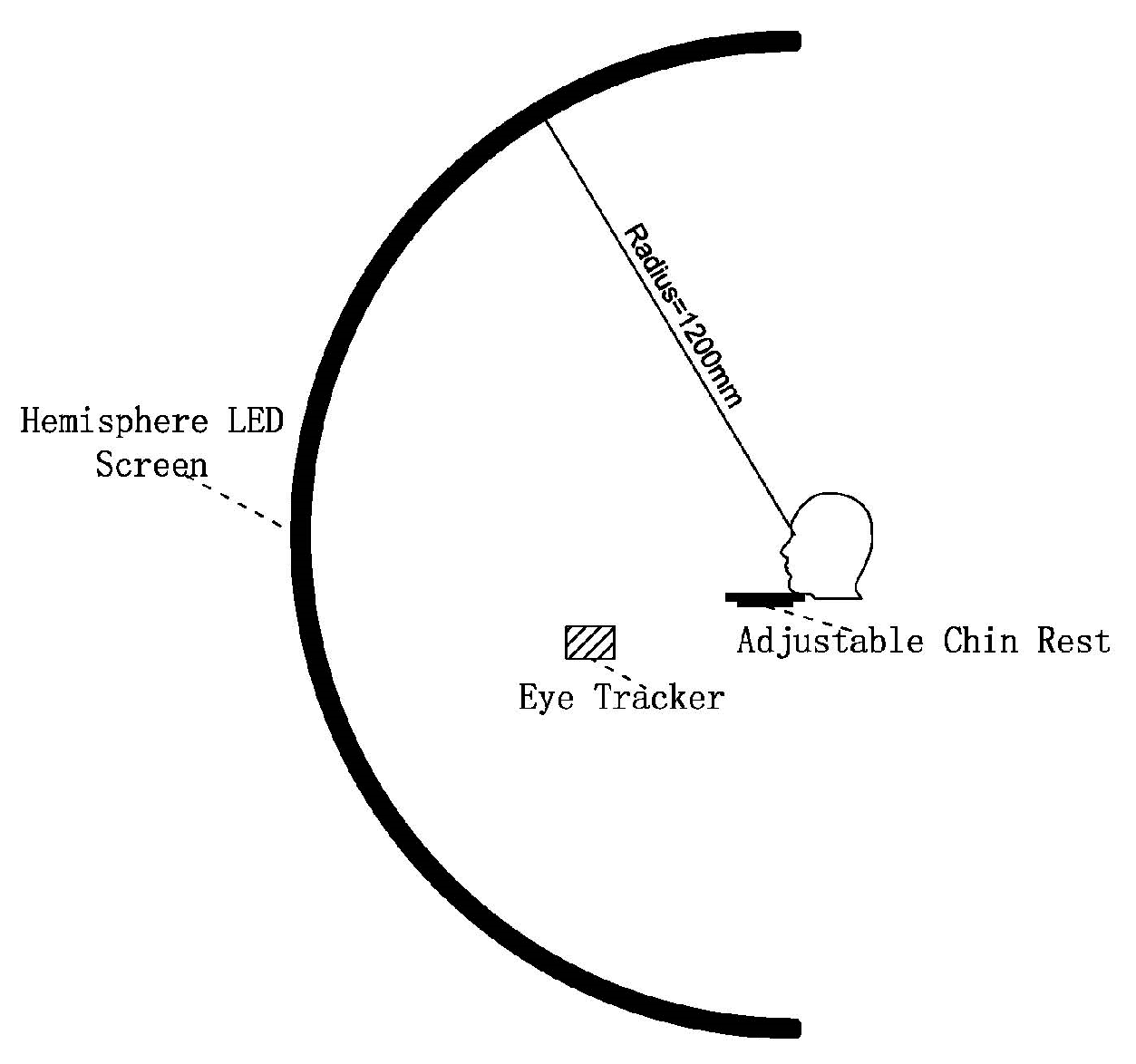
2.2. Apparatus
2.3. Experiment Protocol
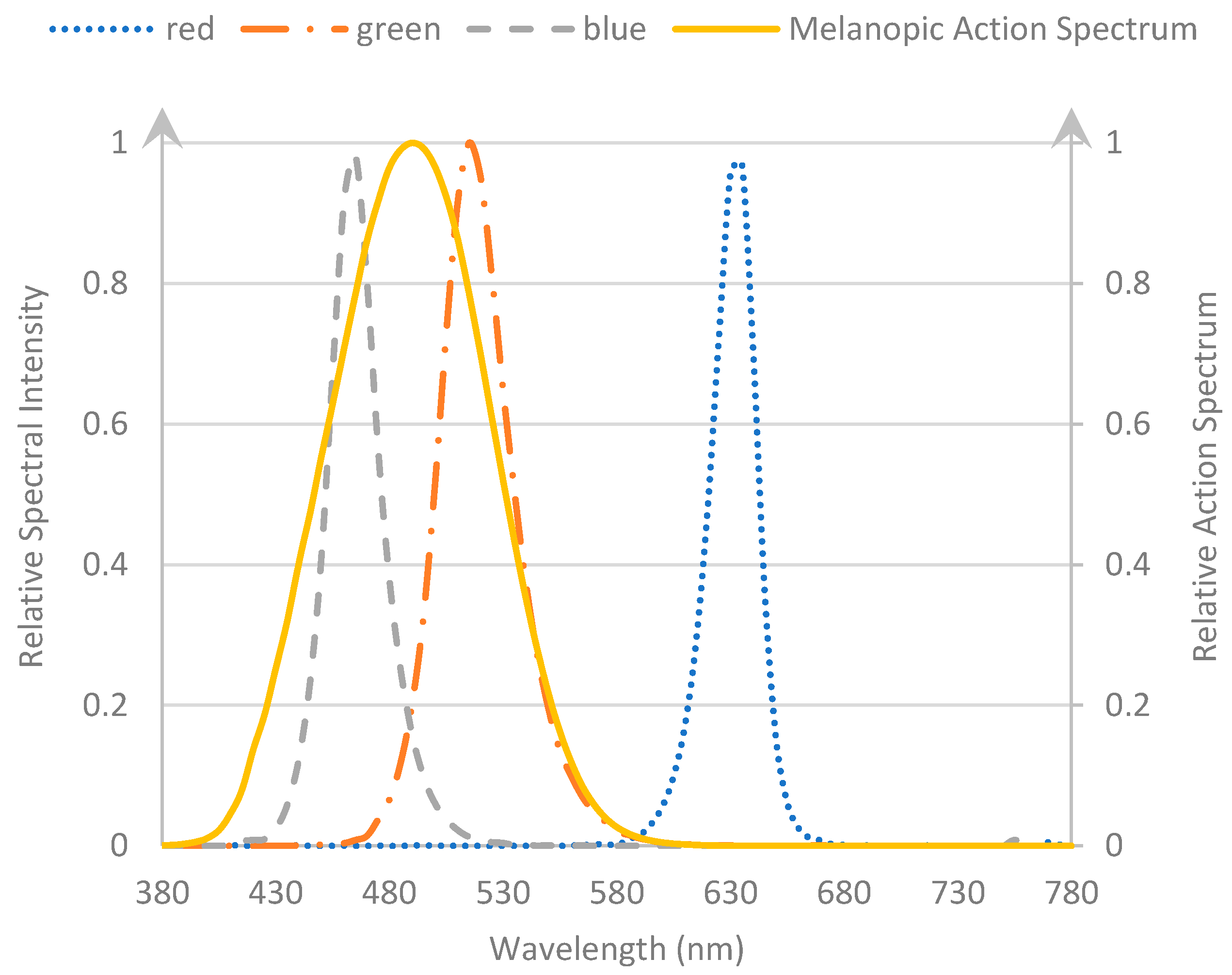
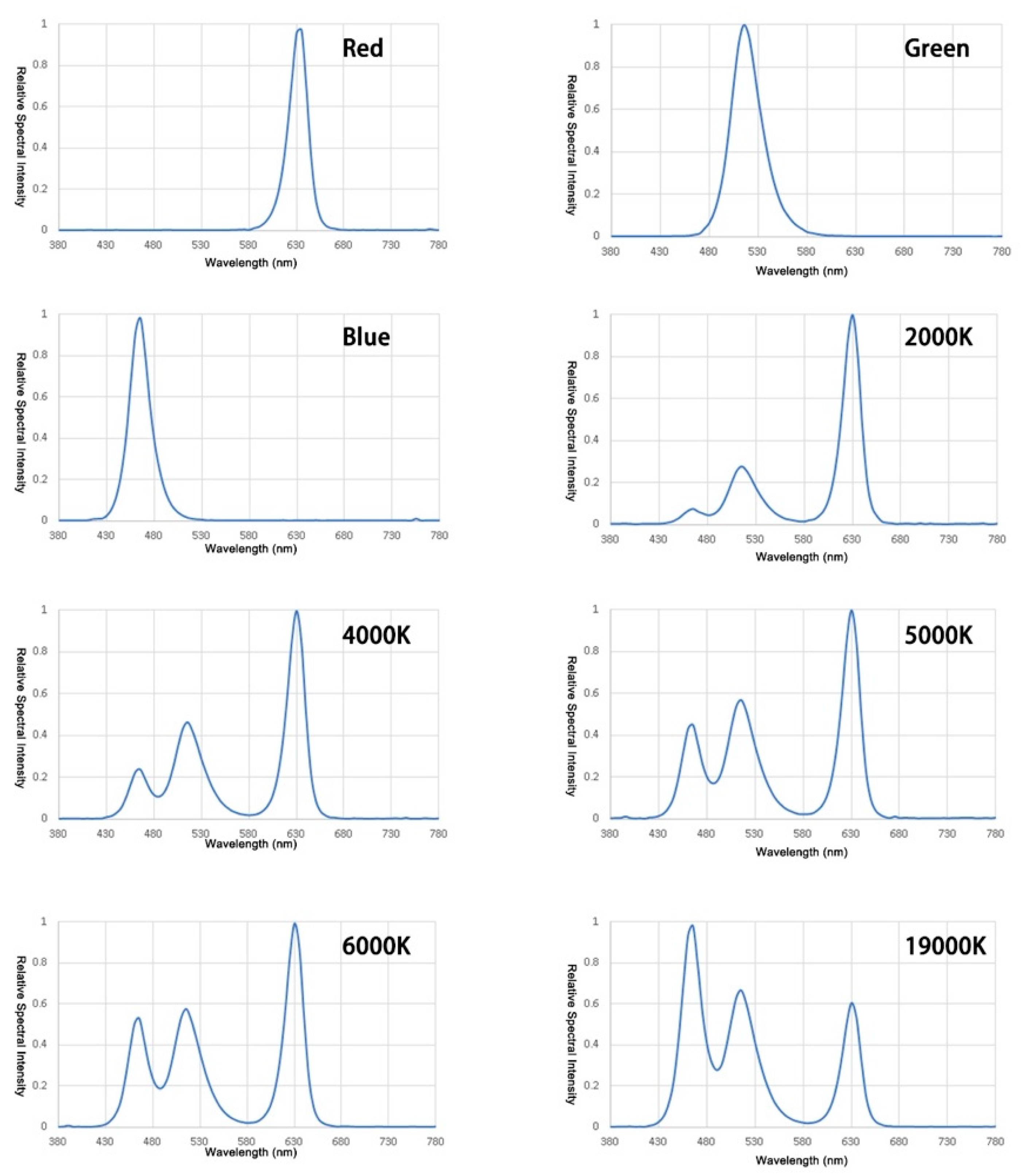
2.4. Stimuli
3. Results
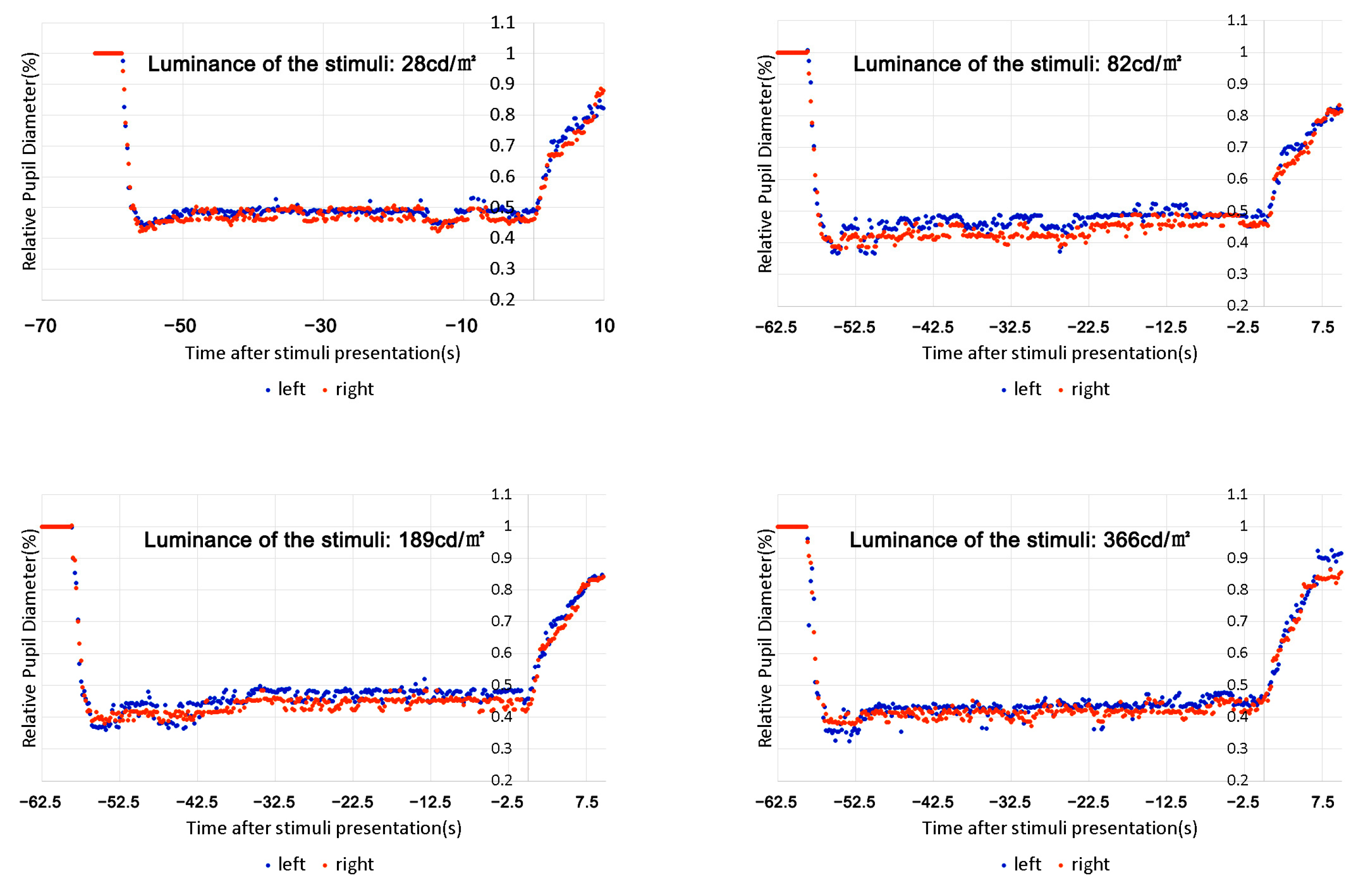
3.1. Verifying the Contribution of the Intrinsic Signal and External Signal to the Steady State of PLR
3.2. Development of a Mathematical Model for the Action Spectrum of Steady-State Pupil Size in Photopic Vision
3.3. Explanation for the Mathematic Model
4. Discussion and Conclusions
- (1)
- We did not find continued dilation of the pupil size after the initial constriction under continuous light presentation for a relatively long time (up to 30 min) but found that the pupil size could reach balance in no more than 10 s, even under a red stimulus that only activated the cones. This suggests that both intrinsic and extrinsic signals participated in the process of regulating pupil size under continuous lighting conditions, unlike the previous report indicating an underestimated contribution of extrinsic signals [21,22,23].
- (2)
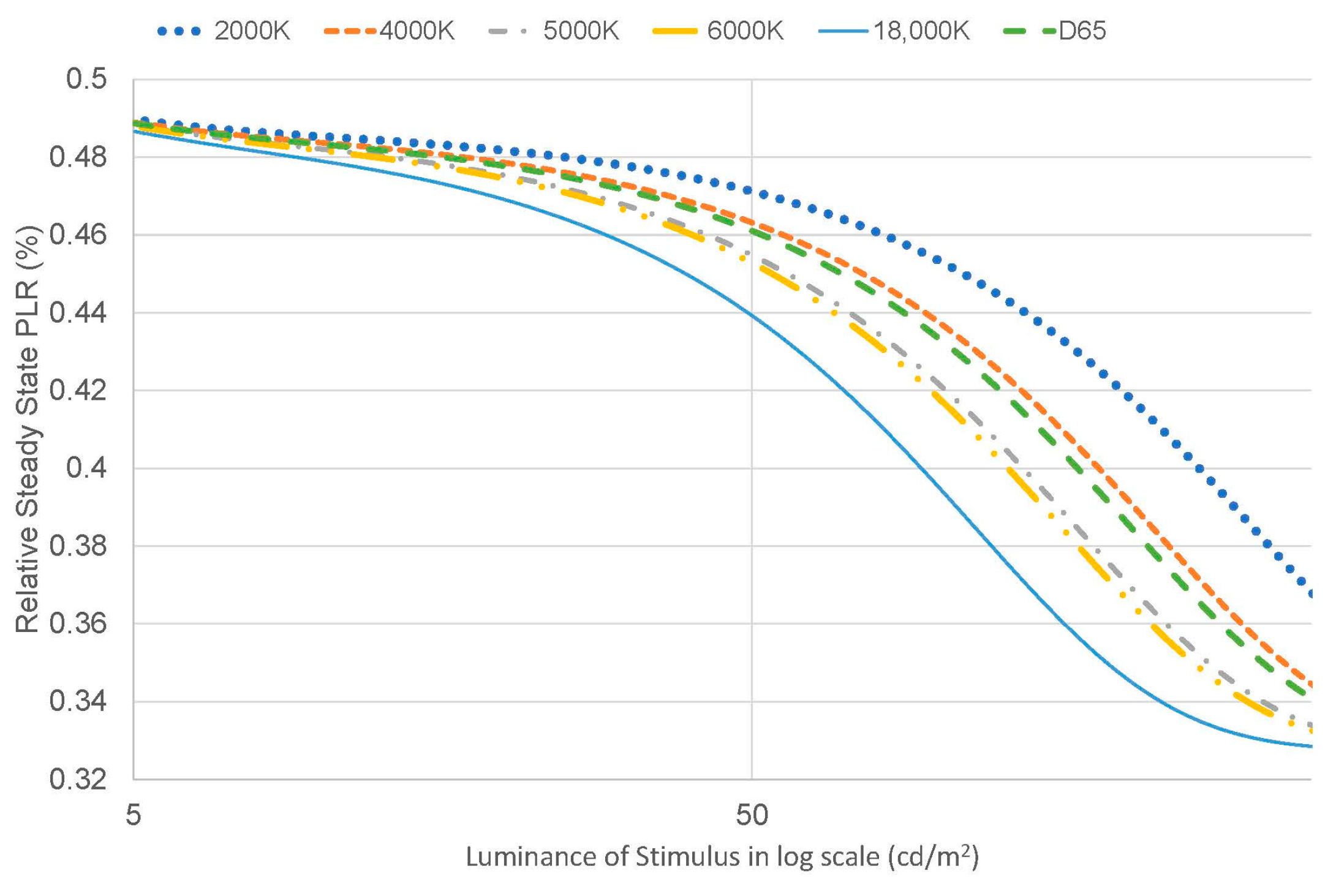
- The contribution of the signals might change with the intensity of melanopic radiance. For melanopic radiance with higher intensity, the intrinsic signal solely determines the pupil size under sustained light presentation, which is consistent with the data from Manuel [20]; however, for photopic conditions with lower melanopic radiance, the contribution of the intrinsic signal decreases, and the extrinsic signal increases with reducing intensity (see Figure 12).
- (3)
- The hypothesis for the potential function of melanopsin in regulating the sensitivity of cones was proposed and verified according to the two experiments. This hypothesis is also supported by the research of Prigge and Mark, who found that M1 ipRGCs, which also regulate the function OPN for the PLR and SCN for behavior, send signals retrogradely to dopaminergic amacrine cells (DACs) within the retina, providing a possible mechanism by which ipRGCs may influence light adaptation via the retinal dopaminergic system [36,38,43]. This may provide new insight into how to explain the mechanism of visual perception.
- (1)
- As M1 ipRGCs are responsible for both the circadian clock and the PLR, research on the PLR might offer us a quick and easy tool to evaluate the impact of different lighting conditions on the nonvisual effect of eyes, such as circadian behaviors.
- (2)
- Many studies have shown that ipRGC dysfunction may have strong relevance to human diseases such as Alzheimer’s disease, Parkinson’s disease, diabetic retinopathy, glaucoma, etc. [44]. Research on the PLR may develop a standard protocol for pupillometry as an emerging method for the direct assessment of ipRGC function in healthy and diseased retinas, which may be used for early detection and clinical examination of these ipRGC-related diseases.
- (3)
- The measurement or estimation of adaptation levels in natural environments with a complex luminance distribution is a very important topic, but there have been no studies directly dealing with this issue [45]. Historically, the steady-state PLR has been seen as an indicator for luminance adaptation, but recent studies on ipRGCs offered us a better mechanistic explanation of the relationship between light adaptation of the visual system and the PLR. This suggests that the same class of ipRGCs for the PLR also functions analogously to a photographer’s light meter, providing a measure to regulate luminance adaptation by synapsing with bipolar and amacrine cells [36,37,46]. Research on the PLR might give us a new perspective on developing a model for light adaptation in a natural environment.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- David, L.D.; Kevin, W.H.; Richard, G.M.; Steffy, G.R. The Lighting Handbook, 10th ed.; Illuminating Engineering Society: New York, NY, USA, 2011. [Google Scholar]
- Hall, C.A.; Chilcott, R.P. Eyeing up the Future of the Pupillary Light Reflex in Neurodiagnostics. Diagnostics 2018, 8, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDougal, D.H.; Gamlin, P.D. The influence of intrinsically-photosensitive retinal ganglion cells on the spectral sensitivity and response dynamics of the human pupillary light reflex. Vis. Res. 2010, 50, 72–87. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.B.; Yellott, J.I. A unified formula for light-adapted pupil size. J. Vis. 2012, 12, 12. [Google Scholar] [CrossRef]
- Berman, S.; Fein, G.; Jewett, D.; Saika, G.; Ashford, F. Spectral Determinants of Steady-State Pupil Size with Full Field of View. J. Illum. Eng. Soc. 1992, 21, 3–13. [Google Scholar] [CrossRef]
- Bouma, H. Size of the Static Pupil as a Function of Wave-length and Luminosity of the Light Incident on the Human Eye. Nature 1962, 193, 690–691. [Google Scholar] [CrossRef]
- Lucas, R.J.; Douglas, R.H.; Foster, R.G. Characterization of an ocular photopigment capable of driving pupillary constriction in mice. Nat. Neurosci. 2001, 4, 621–626. [Google Scholar] [CrossRef]
- Joyce, D.S. Temporal, Spatial and Adaptation Characteristics of Melanopsin Inputs to the Human Pupil Light Reflex. Ph.D. Thesis, Queensland University of Technology, Brisbane City, QLD, Australia, 2016. [Google Scholar]
- Berson, D.M. Strange vision: Ganglion cells as circadian photoreceptors. Trends Neurosci. 2003, 26, 314–320. [Google Scholar] [CrossRef]
- CIE S 026/E:2018; CIE System for Metrology of Optical Radiation for IpRGC-Influenced Responses to Light. Commission Internationale De L’eclairage(Cie) CIE: Vienna, Austria, 2018.
- Dacey, D.; Liao, H.; Peterson, B.; Robinson, F.; Smith, V.; Pokorny, J.; Yau, K.-W.; Gamlin, P.D. Melanopsin-expressing ganglion cells in primate retina signal color and irradiance and project to the lgn. Nature 2005, 433, 749–754. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, J. New prospectives on light adaptation of visual system research with the emerging knowledge on non-imageforming effect. Front. Built Environ. 2022, 8, 1019460. [Google Scholar] [CrossRef]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by Retinal Ganglion Cells That Set the Circadian Clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattar, S.; Liao, H.-W.; Takao, M.; Berson, D.M.; Yau, K.-W. Melanopsin-containing retinal ganglion cells: Architecture, projections, and intrinsic photosensitivity. Science 2002, 295, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Hattar, S.; Lucas, R.J.; Mrosovsky, N.; Thompson, S.; Douglas, R.H.; Hankins, M.W.; Lem, J.; Biel, M.; Hofmann, F.; Foster, R.G.; et al. Melanopsin and rod–cone photoreceptive systems account for all major accessory visual functions in mice. Nature 2003, 424, 75–81. [Google Scholar] [CrossRef] [Green Version]
- Zele, A.J.; Feigl, B.; Adhikari, P.; Maynard, M.L.; Cao, D. Melanopsin photoreception contributes to human visual detection, temporal and colour processing. Sci. Rep. 2018, 8, 3842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Münch, M.; Léon, L.; Crippa, S.V.; Kawasaki, A. Circadian and Wake-Dependent Effects on the Pupil Light Reflex in Response to Narrow-Bandwidth Light Pulses. Investig. Opthalmolo. Vis. Sci. 2012, 53, 4546–4555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, P.; Feigl, B.; Zele, A.J. Rhodopsin and Melanopsin Contributions to the Early Redilation Phase of the Post-Illumination Pupil Response (PIPR). PLoS ONE 2016, 11, e0161175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, F.; Chan, A.; Zhu, X.-F. Effects of photopic and cirtopic illumination on steady state pupil sizes. Vis. Res. 2017, 137, 24–28. [Google Scholar] [CrossRef]
- Spitschan, M. Photoreceptor inputs to pupil control. J. Vis. 2019, 19, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gooley, J.J.; Mien, I.H.; Hilaire, M.A.S.; Yeo, S.C.; Chua, E.C.P.; Van Reen, E.; Hanley, C.J.; Hull, J.T.; Czeisler, C.A.; Lockley, S.W. Melanopsin and Rod-Cone Photoreceptors Play Different Roles in Mediating Pupillary Light Responses during Exposure to Continuous Light in Humans. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 14242–14253. [Google Scholar] [CrossRef] [Green Version]
- Park, J.C.; Moura, A.L.; Raza, A.S.; Rhee, D.W.; Kardon, R.H.; Hood, D.C. Toward a Clinical Protocol for Assessing Rod, Cone, and Melanopsin Contributions to the Human Pupil Response. Investig. Opthalmology Vis. Sci. 2011, 52, 6624–6635. [Google Scholar] [CrossRef] [Green Version]
- Gamlin, P.D.; McDougal, D.H.; Pokorny, J.; Smith, V.C.; Yau, K.-W.; Dacey, D.M. Human and macaque pupil responses driven by melanopsin-containing retinal ganglion cells. Vis. Res. 2007, 47, 946–954. [Google Scholar] [CrossRef] [Green Version]
- Drouyer, E.; Rieux, C.; Hut, R.A.; Cooper, H.M. Responses of Suprachiasmatic Nucleus Neurons to Light and Dark Adaptation: Relative Contributions of Melanopsin and Rod–Cone Inputs. J. Neurosci. 2007, 27, 9623–9631. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.Y.; Dunn, F.A.; Graham, D.M.; Berson, D.M. Synaptic influences on rat ganglion-cell photoreceptors. J. Physiol. 2007, 582, 279–296. [Google Scholar] [CrossRef]
- Lucas, R.J. Diminished Pupillary Light Reflex at High Irradiances in Melanopsin-Knockout Mice. Science 2003, 299, 245–247. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Guo, J.; Wang, S. Clinical Nursing of Correlation between Pupil Changes and Diseases. Chin. Gen. Pract. Nurs. 2014, 12, 1123–1124. (In Chinese) [Google Scholar]
- McDougal, D.H.; Gamlin, P.D.R. Pupillary Control Pathways. In The Senses: A Comprehensive Reference; Academic Press: New York, NY, USA, 2008; pp. 521–536. [Google Scholar]
- Yu, Y.; Jiang, Y.; Wang, Y.; Yu, M. Pupil size as a biomarker of Memory Processing. Adv. Psychol. Sci. 2020, 28, 10. Available online: http://kns.cnki.net/kcms/detail/11.4766.R.20200117.1029.012.html (accessed on 5 January 2023). (In Chinese).
- Dabbs, J.M. Testosterone and Pupillary Response to Auditory Sexual Stimuli. Physiol. Behav. 1997, 62, 909–912. [Google Scholar] [CrossRef]
- Akashi, Y.; Muramatsu, R.; Kanaya, S. Unified Glare Rating (UGR) and subjective appraisal of discomfort glare. Light. Res. Technol. 1996, 28, 199–206. [Google Scholar] [CrossRef]
- Sperandio, I.; Bond, N.; Binda, P. Pupil Size as a Gateway Into Conscious Interpretation of Brightness. Front. Neurol. 2018, 9, 1070. [Google Scholar] [CrossRef]
- Zele, A.J.; Adhikari, P.; Cao, D.; Feigl, B. Melanopsin and Cone Photoreceptor Inputs to the Afferent Pupil Light Response. Front. Neurol. 2019, 10, 529. [Google Scholar] [CrossRef] [PubMed]
- William, A.H.; Rushton, F.R.S. The Plotting of Dark-Adaptation Curves in Rod-Cone Investigations. Am. J. Ophthalmol. 1963, 56, 748–751. [Google Scholar]
- Hung, G.K.; Ciuffreda, K.J. Model of Visual System; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2002; pp. 125–127. [Google Scholar]
- Hankins, M.W.; Hughes, S. Vision: Melanopsin as a novel irradiance detector at the heart of vision. Curr. Biol. 2014, 24, R1055–R1057. [Google Scholar]
- Allen, A.E.; Storchi, R.; Martial, F.P.; Petersen, R.S.; Montemurro, M.A.; Brown, T.M.; Lucas, R.J. Melanopsin-Driven Light Adaptation in Mouse Vision. Curr. Biol. 2014, 24, 2481–2490. [Google Scholar] [CrossRef] [Green Version]
- Prigge, C.L.; Yeh, P.-T.; Liou, N.-F.; Lee, C.-C.; You, S.-F.; Liu, L.-L.; McNeill, D.S.; Chew, K.S.; Hattar, S.; Chen, S.-K.; et al. M1 ipRGCs Influence Visual Function through Retrograde Signaling in the Retina. J. Neurosci. 2016, 36, 7184–7197. [Google Scholar] [CrossRef] [Green Version]
- Brainard, G.C.; Hanifin, J.P.; Greeson, J.; Byrne, B.; Glickman, G.; Gerner, E.; Rollag, M.D. Action Spectrum for Melatonin Regulation in Humans: Evidence for a Novel Circadian Photoreceptor. J. Neurosci. 2001, 21, 6405–6412. [Google Scholar] [CrossRef] [Green Version]
- van Bommel, W. Interior Lighting: Fundamentals, Technology and Application; Springer International Publishing: Cham, Switzerland, 2019; p. 114. [Google Scholar]
- Shapley, R.; Enroth-Cugell, C. Chapter 9 Visual adaptation and retinal gain controls. Prog. Retin. Res. 1984, 3, 263–346. [Google Scholar] [CrossRef]
- Clarke, R.J.; Zhang, H.; Gamlin, P.D.R. Characteristics of the Pupillary Light Reflex in the Alert Rhesus Monkey. J. Neurophysiol. 2003, 89, 3179–3189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, T.M.; Chen, S.-K.; Hattar, S. Intrinsically photosensitive retinal ganglion cells: Many subtypes, diverse functions. Trends Neurosci. 2011, 34, 572–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esquiva, G.; Hannibal, J. Melanopsin-expressing retinal ganglion cells in aging and disease. Histol. Histopathol. 2019, 34, 1299–1311. [Google Scholar] [PubMed]
- International Commission On Illumination (CIE). CIE 135/5:1999 Visual Adaptation to Complex Luminance; CIE: Vienna, Austria, 1999. [Google Scholar]
- Besenecker, U.C. Investigating Melanopsin Contribution to Scene Brightness Perception. Ph.D. Thesis, Rensselaer Polytechnic Institute, Troy, NY, USA, 2013. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Performance |
|---|---|
| Diameter | 2.4 m |
| Number of pixels | 723,360 pixels |
| Color | R/G/B |
| Maximum luminance | >1000 cd/m2 |
| Luminance uniformity | >95% |
| Refresh rate | ≥2000 Hz |
| Bit depth | 12 bit |
| Scenario NO. | Color/ CCT | Measured Luminance (cd/m2) | Melanopic Radiance (W/m2/Sr) | Scenario No. | Color/ CCT | Measured Luminance (cd/m2) | Melanopic Radiance (W/m2/Sr) |
|---|---|---|---|---|---|---|---|
| 1 | Red | 5 | 0.000 | 45 | 3642 | 104 | 0.085 |
| 2 | Red | 13 | 0.000 | 46 | 3594 | 151 | 0.117 |
| 3 | Red | 28 | 0.000 | 47 | 3574 | 208 | 0.158 |
| 4 | Red | 52 | 0.000 | 48 | 3599 | 266 | 0.217 |
| 5 | Red | 82 | 0.000 | 49 | 3619 | 323 | 0.250 |
| 6 | Red | 131 | 0.000 | 50 | 3755 | 448 | 0.326 |
| 7 | Red | 189 | 0.001 | 51 | 3744 | 505 | 0.389 |
| 8 | Red | 261 | 0.001 | 52 | 6685 | 11 | 0.010 |
| 9 | Red | 366 | 0.001 | 53 | 6462 | 18 | 0.017 |
| 10 | Green | 18 | 0.017 | 54 | 6148 | 26 | 0.025 |
| 11 | Green | 46 | 0.045 | 55 | 5833 | 44 | 0.040 |
| 12 | Green | 101 | 0.098 | 56 | 5429 | 69 | 0.063 |
| 13 | Green | 187 | 0.180 | 57 | 5385 | 95 | 0.095 |
| 14 | Green | 310 | 0.299 | 58 | 5270 | 153 | 0.135 |
| 15 | Green | 477 | 0.460 | 59 | 5168 | 220 | 0.187 |
| 16 | Green | 691 | 0.665 | 60 | 5188 | 285 | 0.246 |
| 17 | Green | 961 | 0.926 | 61 | 5192 | 354 | 0.300 |
| 18 | Green | 1353 | 1.303 | 62 | 5280 | 503 | 0.466 |
| 19 | Blue | 5 | 0.035 | 63 | 5350 | 649 | 0.619 |
| 20 | Blue | 7 | 0.057 | 64 | 7095 | 10 | 0.010 |
| 21 | Blue | 16 | 0.124 | 65 | 6959 | 19 | 0.019 |
| 22 | Blue | 30 | 0.230 | 66 | 6406 | 29 | 0.031 |
| 23 | Blue | 50 | 0.382 | 67 | 6026 | 50 | 0.051 |
| 24 | Blue | 77 | 0.587 | 68 | 5749 | 68 | 0.067 |
| 25 | Blue | 112 | 0.851 | 69 | 5757 | 102 | 0.103 |
| 26 | Blue | 156 | 1.184 | 70 | 5648 | 147 | 0.147 |
| 27 | Blue | 220 | 1.672 | 71 | 5571 | 202 | 0.196 |
| 28 | 4336 | 10 | 0.007 | 72 | 5671 | 263 | 0.264 |
| 29 | 3193 | 19 | 0.012 | 73 | 5832 | 302 | 0.302 |
| 30 | 2951 | 29 | 0.019 | 74 | 5740 | 446 | 0.467 |
| 31 | 2734 | 53 | 0.034 | 75 | 5874 | 536 | 0.533 |
| 32 | 2440 | 72 | 0.044 | 76 | 11,091 | 9 | 0.011 |
| 33 | 2123 | 122 | 0.062 | 77 | 12,717 | 14 | 0.019 |
| 34 | 2190 | 153 | 0.086 | 78 | 14,339 | 21 | 0.028 |
| 35 | 2152 | 211 | 0.110 | 79 | 16,877 | 35 | 0.049 |
| 36 | 2126 | 270 | 0.148 | 80 | 19,003 | 55 | 0.079 |
| 37 | 2099 | 307 | 0.167 | 81 | 19,739 | 74 | 0.106 |
| 38 | 2080 | 462 | 0.246 | 82 | 19,560 | 123 | 0.177 |
| 39 | 2101 | 508 | 0.268 | 83 | 19,430 | 175 | 0.254 |
| 40 | 5545 | 10 | 0.009 | 84 | 19,457 | 229 | 0.337 |
| 41 | 4979 | 18 | 0.019 | 85 | 19,756 | 287 | 0.403 |
| 42 | 4302 | 32 | 0.027 | 86 | 19,652 | 403 | 0.591 |
| 43 | 3836 | 53 | 0.039 | 87 | 19,613 | 540 | 0.804 |
| 44 | 3854 | 70 | 0.059 | —— | —— | —— | —— |
| LSD | |||
|---|---|---|---|
| (I) Time after the Stimulus Onset | (J) Time after the Stimulus Onset | Mean Difference (I–J) | Sig. |
| 1.00 | 11.00 | 0.42839 * | 0.000 |
| 21.00 | 0.42766 * | 0.000 | |
| 31.00 | 0.42221 * | 0.000 | |
| 41.00 | 0.42215 * | 0.000 | |
| 55.00 | 0.41890 * | 0.000 | |
| 11.00 | 1.00 | −0.42839 * | 0.000 |
| 21.00 | −0.00073 | 0.980 | |
| 31.00 | −0.00618 | 0.831 | |
| 41.00 | −0.00624 | 0.829 | |
| 55.00 | −0.00949 | 0.743 | |
| 21.00 | 1.00 | −0.42766 * | 0.000 |
| 11.00 | 0.00073 | 0.980 | |
| 31.00 | −0.00545 | 0.851 | |
| 41.00 | −0.00551 | 0.849 | |
| 55.00 | −0.00875 | 0.762 | |
| 31 | 1.00 | −0.42221 * | 0.000 |
| 11.00 | 0.00618 | 0.831 | |
| 21.00 | 0.00545 | 0.851 | |
| 41.00 | −0.00006 | 0.998 | |
| 55.00 | −0.00331 | 0.909 | |
| 41.00 | 1.00 | −0.42215 * | 0.000 |
| 11.00 | 0.00624 | 0.829 | |
| 21.00 | 0.00551 | 0.849 | |
| 31.00 | 0.00006 | 0.998 | |
| 55.00 | −0.00324 | 0.911 | |
| 55.00 | 1.00 | −0.41890 * | 0.000 |
| 11.00 | 0.00949 | 0.743 | |
| 21.00 | 0.00875 | 0.762 | |
| 31.00 | 0.00331 | 0.909 | |
| 41.00 | 0.00324 | 0.911 | |
| Dependent Factor: Relative Pupil Diameter | ||||||
|---|---|---|---|---|---|---|
| LSD | ||||||
| (I) Luminance of Stimulus (cd/m2) | (J) Luminance of Stimulus (cd/m2) | Mean Difference (I–J) | Std. Error | Sig. | 95% Confidence Interval | |
| Lower Bound | Upper Bound | |||||
| 5 | 28 | 0.05306 * | 0.003044 | 00.000 | 0.04708 | 0.05903 |
| 82 | 0.08648 * | 0.003033 | 0.000 | 0.08052 | 0.09243 | |
| 189 | 0.11041 * | 0.003033 | 0.000 | 0.10446 | 0.11636 | |
| 366 | 0.11060 * | 0.003037 | 0.000 | 0.10464 | 0.11655 | |
| 28 | 5 | −0.05306 * | 0.003044 | 0.000 | −0.05903 | −0.04708 |
| 82 | 0.03342 * | 0.003129 | 0.000 | 0.02728 | 0.03956 | |
| 189 | 0.05736 * | 0.003129 | 0.000 | 0.05122 | 0.06349 | |
| 366 | 0.05754 * | 0.003132 | 0.000 | 0.05140 | 0.06369 | |
| 82 | 5 | −0.08648 * | 0.003033 | 0.000 | −0.09243 | −0.08052 |
| 28 | −0.03342 * | 0.003129 | 0.000 | −0.03956 | −0.02728 | |
| 189 | 0.02394 * | 0.003118 | 0.000 | 0.01782 | 0.03005 | |
| 366 | 0.02412 * | 0.003122 | 0.000 | 0.01800 | 0.03025 | |
| 189 | 5 | −0.11041 * | 0.003033 | 0.000 | −0.11636 | −0.10446 |
| 28 | −0.05736 * | 0.003129 | 0.000 | −0.06349 | −0.05122 | |
| 82 | −0.02394 * | 0.003118 | 0.000 | −0.03005 | −0.01782 | |
| 366 | 0.00019 | 0.003122 | 0.953 | −0.00594 | 0.00631 | |
| 366 | 5 | −0.11060 * | 0.003037 | 0.000 | −0.11655 | −0.10464 |
| 28 | −0.05754 * | 0.003132 | 0.000 | −0.06369 | −0.05140 | |
| 82 | −0.02412 * | 0.003122 | 0.000 | −0.03025 | −0.01800 | |
| 189 | −0.00019 | 0.003122 | 0.953 | −0.00631 | 0.00594 | |
| Source | Sum of Squares | df | Mean Squares |
|---|---|---|---|
| Regression | 38.364 | 4 | 9.591 |
| Residual | 0.231 | 83 | 0.003 |
| Uncorrected Total | 38.594 | 87 | |
| Corrected Total | 1.271 | 86 |
| Source | Sum of Squares | df | Mean Squares |
|---|---|---|---|
| Regression | 15.145 | 4 | 3.786 |
| Residual | 0.024 | 83 | 0.000 |
| Uncorrected Total | 15.169 | 87 | |
| Corrected Total | 0.342 | 86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Zhao, J.; Wang, L.; Hu, W.; Yan, F. A Mathematical Model for the Action Spectrum of Steady-State Pupil Size in Photopic Vision with Insight into Healthful Lighting. Buildings 2023, 13, 781. https://doi.org/10.3390/buildings13030781
Wang S, Zhao J, Wang L, Hu W, Yan F. A Mathematical Model for the Action Spectrum of Steady-State Pupil Size in Photopic Vision with Insight into Healthful Lighting. Buildings. 2023; 13(3):781. https://doi.org/10.3390/buildings13030781
Chicago/Turabian StyleWang, Shuxiao, Jianping Zhao, Lixiong Wang, Wenye Hu, and Fanfang Yan. 2023. "A Mathematical Model for the Action Spectrum of Steady-State Pupil Size in Photopic Vision with Insight into Healthful Lighting" Buildings 13, no. 3: 781. https://doi.org/10.3390/buildings13030781