
Microbiologically Influenced Corrosion Mechanism of Ferrous Alloys in Marine Environment

Abstract
:1. Introduction

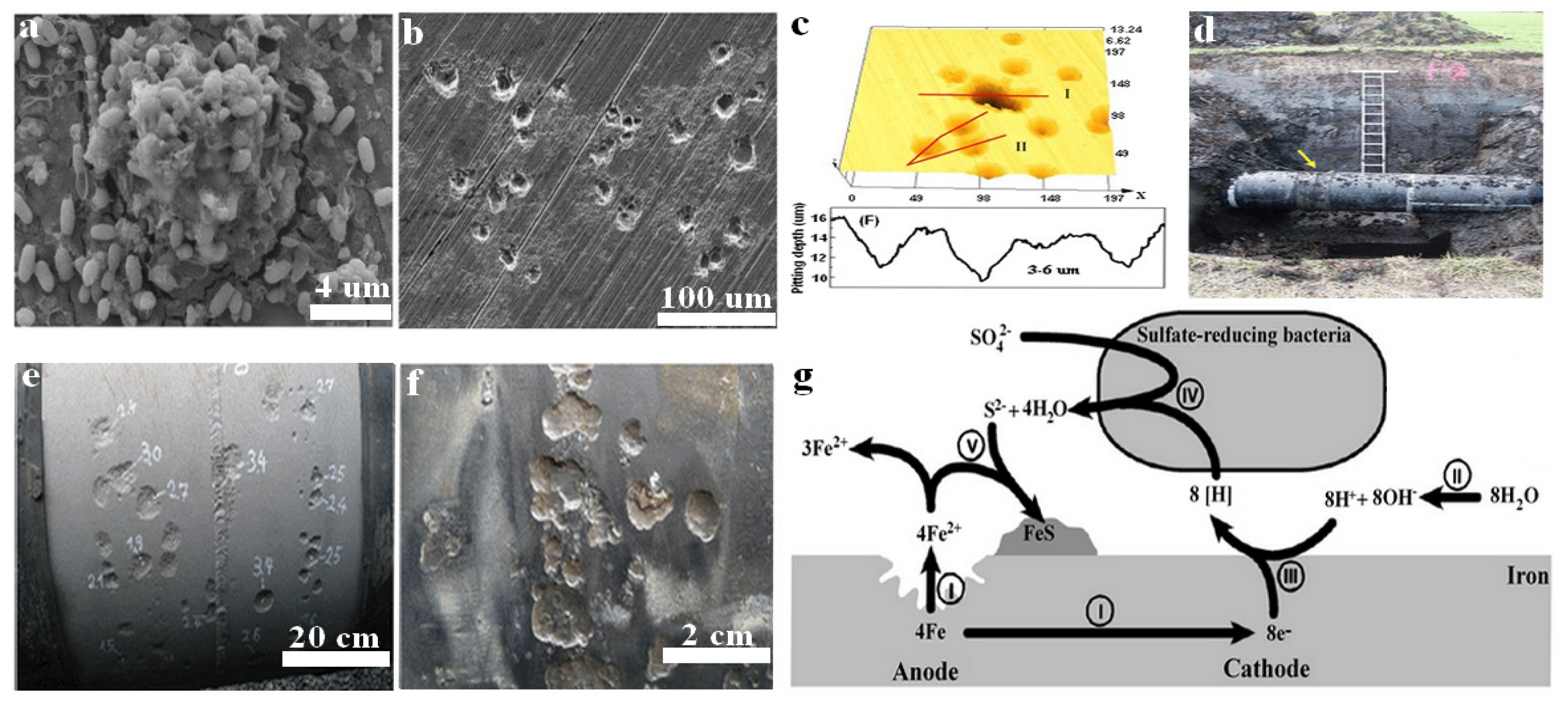{kind=link}
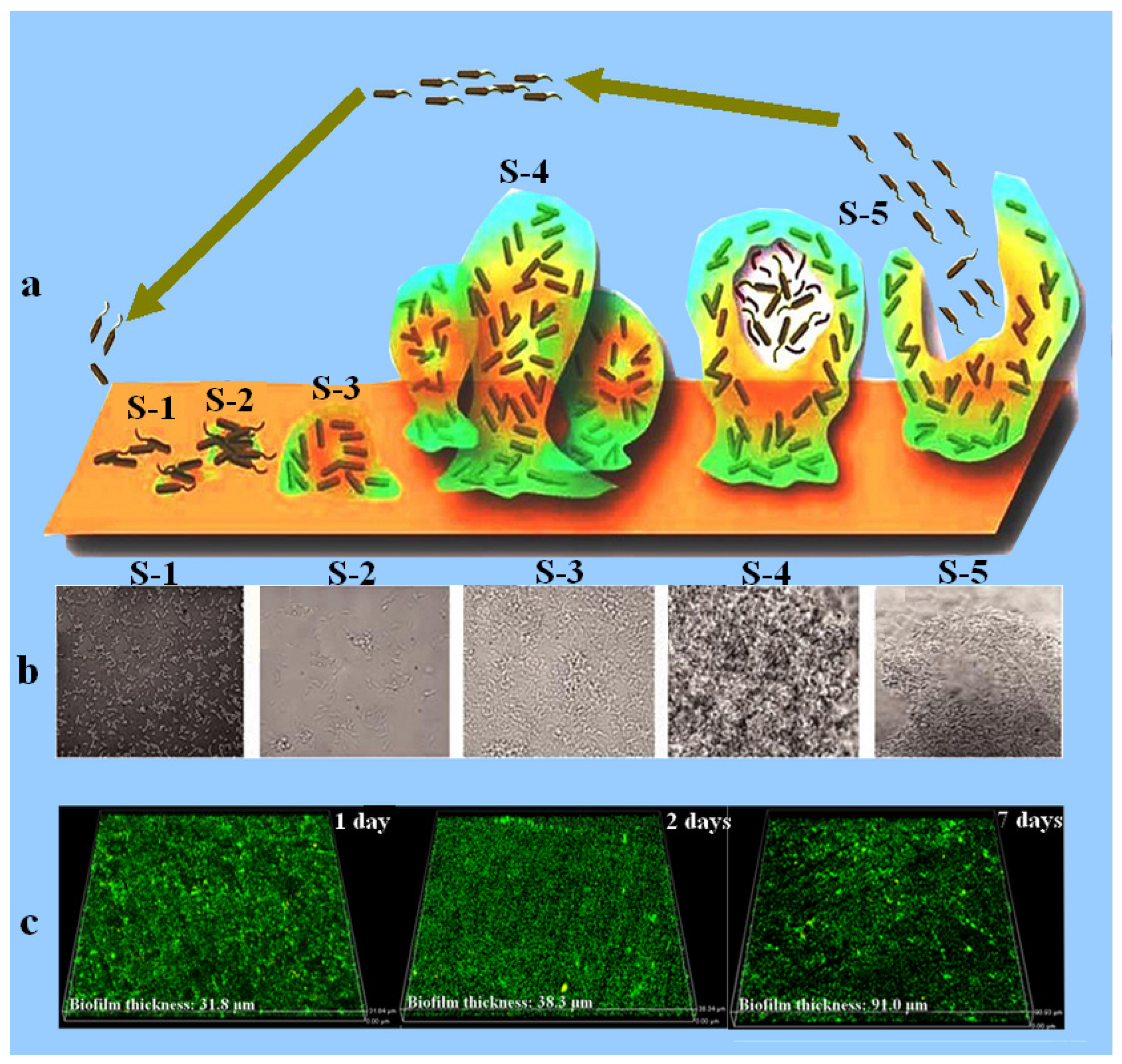{kind=link}
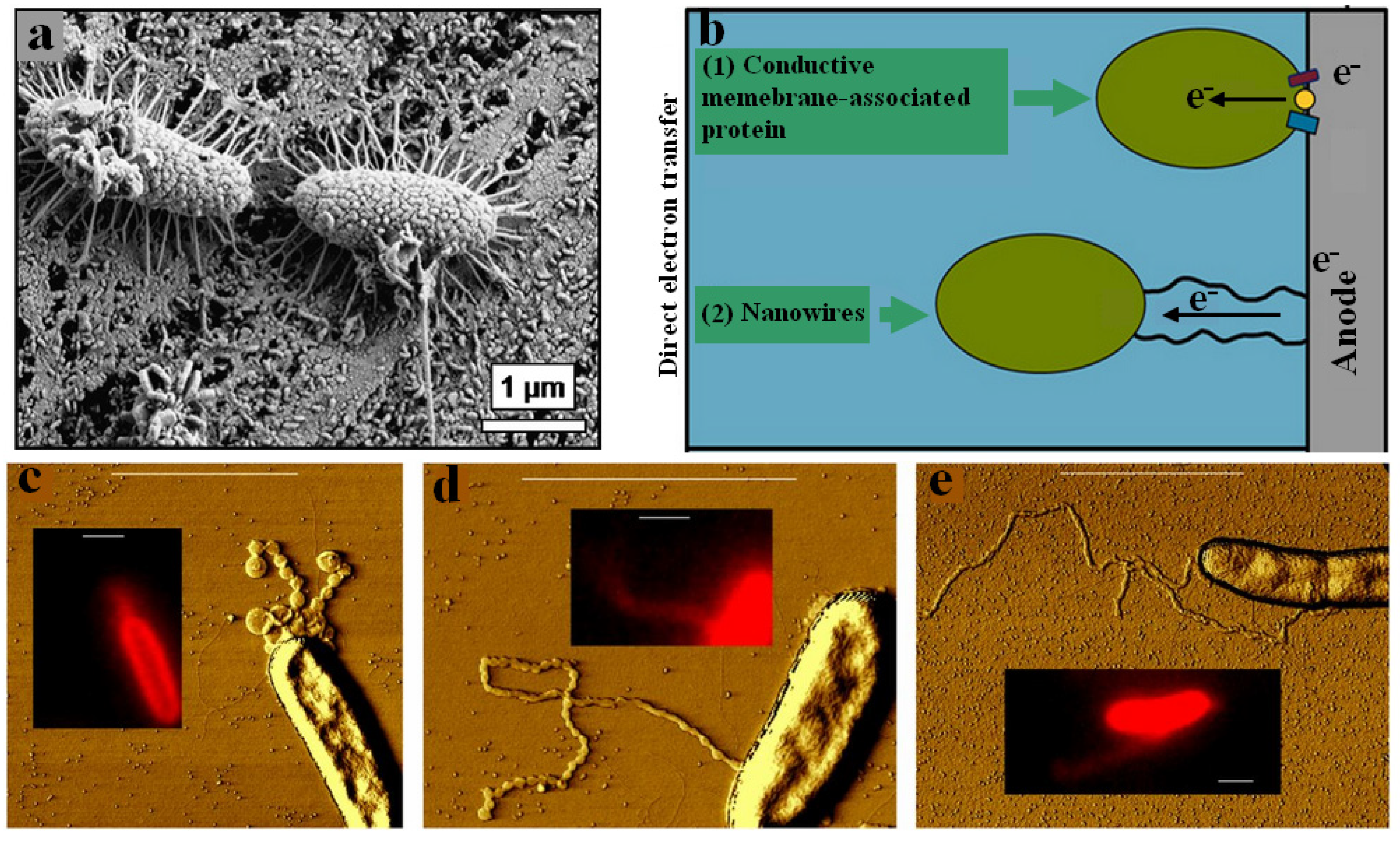{kind=link}
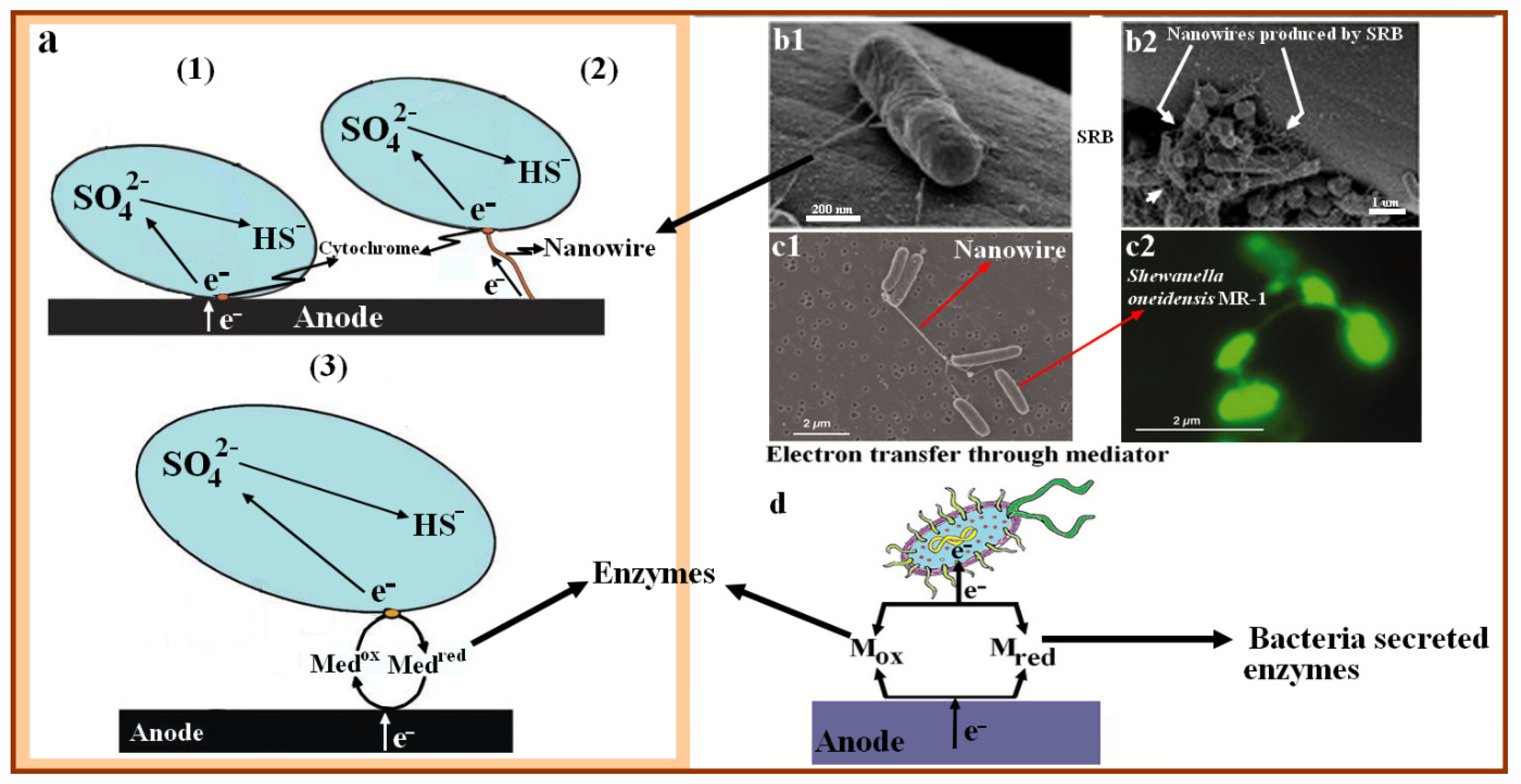{kind=link}
| Materials | Authors | Microbes | Effects |
|---|---|---|---|
| carbon steel | Hamza et al. [4] | P. aeruginosa | Biofilm formation, corrosion damages, high corrosion rate |
| 304 SS | Hamza et al. [5] | P. aeruginosa | Biofilm formation, formation of differential aeration cells, pitting corrosion |
| Titanium | Khan et al. [7] | P. aeruginosa | Biofilm formation, pitting corrosion |
| Carbon steel | Javed et al. [8] | SRB | Biofilm formation, EPS secretion, pitting corrosion attacks, high corrosion rate |
| Iron based oil and gas pipelines | Dennis et al. [22] | SRB | Production of corrosive hydrogen sulfide, increased deterioration of iron |
| Low carbon steel (Ship ballast tank) | Heyer et al. [26] | Slime forming bacteria, sulfur oxidizing (SOB), iron oxidizing bacteria (IOB) and sulphate reducing bacteria (SRB) | Increase pitting corrosion, high corrosion rate |
| 316L stainless steel | Dong et al. [27] | Acidithiobacillus caldus SM-1 | Development of dense biofilm, severe pitting corrosion, high corrosion rate |
| 316L stainless steel | Tang et al. 28 | Geobacter sulfurreducens and Geobacter metallireducens | Direct electron transfer encouraged the corrosion of stainless steel |
| 304 stainless steel | Zhang et al. [29] | Desulfovibrio vulgaris | Electron mediator increased corrosion, weight loss, pitting corrosion |
2. Corrosion Induced by the Concentration Cells
3. Metabolites Induced Corrosion
4. MIC Based on Bioenergetics and Bio-Electrochemistry
4.1. Direct Electron Transfer
4.2. Electron Mediator
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Little, B.J.; Lee, J.S. Microbiologically Influenced Corrosion. In Engineering Materials and Processes; Springer: London, UK, 2015; Volume 2, ISBN 9781119019213. [Google Scholar]
- Marciales, A.; Peralta, Y.; Haile, T.; Crosby, T.; Wolodko, J. Mechanistic microbiologically influenced corrosion modeling—A review. Corros. Sci. 2019, 146, 99–111. [Google Scholar]
- Suarez, E.M.; Lepkova, K.; Kinsella, B.; Machuca, L.L. Aggressive corrosion of steel by a thermophilic microbial consortium in the presence and absence of sand. Int. Biodeterior. Biodegrad. 2019, 137, 137–146. [Google Scholar] [CrossRef]
- Hamzah, E.; Hussain, M.F.; Ibrahim, Z.; Abdolahi, A. Corrosion Behaviour of Carbon Steel in Sea Water Medium in Presence of P. aeruginosa Bacteria. Arab. J. Sci. Eng. 2014, 39, 6863–6870. [Google Scholar] [CrossRef]
- Hamzah, E.; Hussain, M.Z.; Ibrahim, Z.; Abdolahi, A. Influence of Pseudomonas aeruginosa bacteria on corrosion resistance of 304 stainless steel. Corros. Eng. Sci. Technol. 2013, 48, 116–120. [Google Scholar] [CrossRef]
- Farooq, A.; Zubair, M.; Wadood, H.Z.; Deen, K.M. Effect of Pseudomonas aeruginosa Strain ZK Biofilm on the Mechanical and Corrosion Behavior of 316L Stainless Steel and α-brass. J. Electrochem. Sci. Technol. 2021, 12, 431–439. [Google Scholar] [CrossRef]
- Khan, M.S.; Li, Z.; Yang, K.; Xu, D.; Yang, C.; Liu, D.; Lekbach, Y.; Zhou, E.; Kalnaowakul, P. Microbiologically influenced corrosion of titanium caused by aerobic marine bacterium Pseudomonas aeruginosa. J. Mater. Sci. Technol. 2019, 35, 216–222. [Google Scholar] [CrossRef]
- Javed, M.A.; Stoddart, P.R.; Wade, S.A. Corrosion of carbon steel by sulphate reducing bacteria: Initial attachment and the role of ferrous ions. Corros. Sci. 2015, 93, 48–57. [Google Scholar] [CrossRef]
- Bradley, G.; Gaylarde, C.C. Iron uptake by Desulfovibrio vulgaris outer membrane components in artificial vesicles. Curr. Microbiol. 1988, 17, 189–192. [Google Scholar] [CrossRef]
- Javed, M.A.; Neil, W.C.; McAdam, G.; Wade, S.A. Effect of sulphate-reducing bacteria on the microbiologically influenced corrosion of ten different metals using constant test conditions. Int. Biodeterior. Biodegrad. 2017, 125, 73–85. [Google Scholar] [CrossRef]
- Li, Z.; Wang, J.; Dong, Y.; Xu, D.; Zhang, X.; Wu, J.; Gu, T.; Wang, F. Synergistic effect of chloride ion and Shewanella algae accelerates the corrosion of Ti-6Al-4V alloy. J. Mater. Sci. Technol. 2021, 71, 177–185. [Google Scholar] [CrossRef]
- Alabbas, F.M.; Mishra, B. Microbiologically influenced corrosion of pipelines in the oil & gas industry. In Proceedings of the 8th Pacific Rim International Congress on Advanced Materials and Processing 2013, PRICM 8, Waikoloa, Hawaii, 4–9 August 2013; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; Volume 4, pp. 3441–3448. [Google Scholar]
- Zarasvand, K.A.; Rai, V.R. Microorganisms: Induction and inhibition of corrosion in metals. Int. Biodeterior. Biodegrad. 2014, 87, 66–74. [Google Scholar] [CrossRef]
- Beech, I.B. Corrosion of technical materials in the presence of biofilms—Current understanding and state-of-the art methods of study. Int. Biodeterior. Biodegrad. 2004, 53, 177–183. [Google Scholar] [CrossRef]
- Videla, H.A. Prevention and control of biocorrosion. Int. Biodeterior. Biodegrad. 2002, 49, 259–270. [Google Scholar] [CrossRef]
- Zhang, H.-J.; Dexter, S.C. Effect of biofilms on crevice corrosion of stainless steels in coastal seawater. Corrosion 1995, 51, 56–66. [Google Scholar] [CrossRef]
- Machuca, L.L.; Bailey, S.I.; Gubner, R.; Watkin, E.L.J.; Ginige, M.P.; Kaksonen, A.H.; Heidersbach, K. Effect of oxygen and biofilms on crevice corrosion of UNS S31803 and UNS N08825 in natural seawater. Corros. Sci. 2013, 67, 242–255. [Google Scholar] [CrossRef]
- Machuca, L.L.; Bailey, S.I.; Gubner, R. Microbial Corrosion Resistance of Stainless Steels for Marine Energy Installations. Adv. Mater. Res. 2012, 353, 3591–3596. [Google Scholar] [CrossRef]
- Little, B.J.; Lee, J.S.; Ray, R.I. The influence of marine biofilms on corrosion: A concise review. Electrochim. Acta 2008, 54, 2–7. [Google Scholar] [CrossRef]
- Beech, I.B.; Sunner, J. Biocorrosion: Towards understanding interactions between biofilms and metals. Curr. Opin. Biotechnol. 2004, 15, 181–186. [Google Scholar] [CrossRef]
- Iverson, W.P. Research on the mechanisms of anaerobic corrosion. Int. Biodeterior. Biodegrad. 2001, 47, 63–70. [Google Scholar] [CrossRef]
- Enning, D.; Garrelfs, J. Corrosion of iron by sulfate-reducing bacteria: New views of an old problem. Appl. Environ. Microbiol. 2014, 80, 1226–1236. [Google Scholar] [CrossRef]
- Anandkumar, B.; George, R.P.; Maruthamuthu, S.; Parvathavarthini, N.; Mudali, U.K. Corrosion characteristics of sulfate-reducing bacteria (SRB) and the role of molecular biology in SRB studies: An overview. Corros. Rev. 2016, 34, 41–63. [Google Scholar]
- Loto, C.A. Microbiological corrosion: Mechanism, control and impact—A review. Int. J. Adv. Manuf. Technol. 2017, 92, 4241–4252. [Google Scholar] [CrossRef]
- Kip, N.; Van Veen, J.A. The dual role of microbes in corrosion. ISME J. 2015, 9, 542–551. [Google Scholar] [CrossRef]
- Heyer, A.; Souza, F.D.; Morales, C.F.L.; Ferrari, G.; Mol, J.M.C.; Wit, J.H.W. De Ship ballast tanks a review from microbial corrosion and electrochemical point of view. Ocean Eng. 2013, 70, 188–200. [Google Scholar] [CrossRef]
- Dong, Y.; Li, J.; Xu, D.; Song, G.; Liu, D.; Wang, H.; Saleem Khan, M.; Yang, K.; Wang, F. Investigation of microbial corrosion inhibition of Cu-bearing 316L stainless steel in the presence of acid producing bacterium Acidithiobacillus caldus SM-1. J. Mater. Sci. Technol. 2020. [Google Scholar] [CrossRef]
- Tang, H.Y.; Yang, C.; Ueki, T.; Pittman, C.C.; Xu, D.; Woodard, T.L.; Holmes, D.E.; Gu, T.; Wang, F.; Lovley, D.R. Stainless steel corrosion via direct iron-to-microbe electron transfer by Geobacter species. ISME J. 2021. [Google Scholar] [CrossRef]
- Zhang, P.; Xu, D.; Li, Y.; Yang, K.; Gu, T. Electron mediators accelerate the microbiologically influenced corrosion of 304 stainless steel by the Desulfovibrio vulgaris biofilm. Bioelectrochemistry 2015, 101, 14–21. [Google Scholar] [CrossRef]
- Caldwell, D.E.; Korber, D.R.; Lawrence, J.R. Confocal Laser Microscopy and Digital Image Analysis in Microbial Ecology. In Advances in Microbial Ecology; Springer: Boston, MA, USA, 1992; Volume 12, pp. 1–67. [Google Scholar] [CrossRef]
- Alemayehu, T.; Birahane, M. Corrosion and its Protection. IJASR Int. J. Acad. Sci. Res. 2014, 2, 2272–6446. [Google Scholar]
- Ray, R.I.; Lee, J.S.; Little, B.J. Iron-oxidizing bacteria: A review of corrosion mechanisms in fresh water and marine environments. In Proceedings of the NACE—International Corrosion Conference Series, San Antonio, TX, USA, 14–18 March 2010. [Google Scholar]
- Ray, R.; Lee, J.; Little, B. Factors contributing to corrosion of steel pilings in Duluth-Superior Harbor. Corrosion 2009, 65, 707–717. [Google Scholar] [CrossRef]
- Li, Y.; Feng, S.; Liu, H.; Tian, X.; Xia, Y.; Li, M.; Xu, K.; Yu, H.; Liu, Q.; Chen, C. Bacterial distribution in SRB biofilm affects MIC pitting of carbon steel studied using FIB-SEM. Corros. Sci. 2020, 167, 108512. [Google Scholar] [CrossRef]
- Blackwood, D. An Electrochemist Perspective of Microbiologically Influenced Corrosion. Corros. Mater. Degrad. 2018, 1, 59–76. [Google Scholar] [CrossRef]
- Dong, C.; Bai, X.; Yan, X.; Yuan, C. Research status and advances on tribological study of materials under ocean environment. Tribology 2013, 33, 311–320. [Google Scholar]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as Complex Differentiated Communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verderosa, A.D.; Totsika, M.; Fairfull-Smith, K.E. Bacterial Biofilm Eradication Agents: A Current Review. Front. Chem. 2019, 7, 824. [Google Scholar] [CrossRef] [PubMed]
- Little, B.J.; Blackwood, D.J.; Hinks, J.; Lauro, F.M.; Marsili, E.; Okamoto, A.; Rice, S.A.; Wade, S.A.; Flemming, H.C. Microbially influenced corrosion—Any progress? Corros. Sci. 2020, 170, 108641. [Google Scholar] [CrossRef]
- Yuan, S.; Liang, B.; Zhao, Y.; Pehkonen, S.O. Surface chemistry and corrosion behaviour of 304 stainless steel in simulated seawater containing inorganic sulphide and sulphate-reducing bacteria. Corros. Sci. 2013, 74, 353–366. [Google Scholar] [CrossRef]
- Ćwiek, J. Prevention methods against hydrogen degradation of steel. J. Achiev. Mater. Manuf. Eng. 2010, 43, 214–221. [Google Scholar]
- Deckena, S.; Blotevogel, K.H. Growth of methanogenic and sulfate-reducing bacteria with cathodic hydrogen. Biotechnol. Lett. 1990, 12, 615–620. [Google Scholar] [CrossRef]
- Gu, T.; Jia, R.; Unsal, T.; Xu, D. Toward a better understanding of microbiologically influenced corrosion caused by sulfate reducing bacteria. J. Mater. Sci. Technol. 2019, 35, 631–636. [Google Scholar] [CrossRef]
- Xu, D.; Li, Y.; Song, F.; Gu, T. Laboratory investigation of microbiologically influenced corrosion of C1018 carbon steel by nitrate reducing bacterium Bacillus licheniformis. Corros. Sci. 2013, 77, 385–390. [Google Scholar] [CrossRef]
- Gu, T. New understandings of biocorrosion mechanisms and their classifications. J. Microb. Biochem. Technol. 2012, 4, iii–vi. [Google Scholar] [CrossRef]
- Lampreia, J.; Fauque, G.; Speich, N.; Dahl, C.; Moura, I.; Truper, H.G.; Moura, J.J.G. Spectroscopic studies on APS reductase isolated from the hyperthermophilic sulfate-reducing archaebacterium Archaeglobus fulgidus. Biochem. Biophys. Res. Commun. 1991, 181, 342–347. [Google Scholar] [CrossRef]
- Xu, D.; Gu, T. Carbon source starvation triggered more aggressive corrosion against carbon steel by the Desulfovibrio vulgaris biofilm. Int. Biodeterior. Biodegrad. 2014, 91, 74–81. [Google Scholar] [CrossRef]
- Jia, R.; Yang, D.; Xu, J.; Xu, D.; Gu, T. Microbiologically influenced corrosion of C1018 carbon steel by nitrate reducing Pseudomonas aeruginosa biofilm under organic carbon starvation. Corros. Sci. 2017, 127, 1–9. [Google Scholar] [CrossRef]
- Lovley, D.R. The microbe electric: Conversion of organic matter to electricity. Curr. Opin. Biotechnol. 2008, 19, 564–571. [Google Scholar] [CrossRef]
- Kato, S.; Hashimoto, K.; Watanabe, K. Microbial interspecies electron transfer via electric currents through conductive minerals. Proc. Natl. Acad. Sci. USA 2012, 109, 10042–10046. [Google Scholar] [CrossRef]
- Kato, S.; Yumoto, I.; Kamagata, Y. Isolation of acetogenic bacteria that induce biocorrosion by utilizing metallic iron as the sole electron donor. Appl. Environ. Microbiol. 2015, 81, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Rabaey, K.; Rozendal, R.A. Microbial electrosynthesis—Revisiting the electrical route for microbial production. Nat. Rev. Microbiol. 2010, 8, 706–716. [Google Scholar] [CrossRef]
- Lovley, D.R. Powering microbes with electricity: Direct electron transfer from electrodes to microbes. Environ. Microbiol. Rep. 2011, 3, 27–35. [Google Scholar] [CrossRef]
- Nevin, K.P.; Hensley, S.A.; Franks, A.E.; Summers, Z.M.; Ou, J.; Woodard, T.L.; Snoeyenbos-West, O.L.; Lovley, D.R. Electrosynthesis of organic compounds from carbon dioxide is catalyzed by a diversity of acetogenic microorganisms. Appl. Environ. Microbiol. 2011, 77, 2882–2886. [Google Scholar] [CrossRef]
- Pfeffer, C.; Larsen, S.; Song, J.; Dong, M.; Besenbacher, F.; Meyer, R.L.; Kjeldsen, K.U.; Schreiber, L.; Gorby, Y.A.; El-Naggar, M.Y.; et al. Filamentous bacteria transport electrons over centimetre distances. Nature 2012, 491, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.P.; Risgaard-Petersen, N.; Fossing, H.; Christensen, P.B.; Sayama, M. Electric currents couple spatially separated biogeochemical processes in marine sediment. Nature 2010, 463, 1071–1074. [Google Scholar] [CrossRef] [PubMed]
- Bond, D.R.; Lovley, D.R. Electricity production by Geobacter sulfurreducens attached to electrodes. Appl. Environ. Microbiol. 2003, 69, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- Gregory, K.B.; Bond, D.R.; Lovley, D.R. Graphite electrodes as electron donors for anaerobic respiration. Environ. Microbiol. 2004, 6, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Enning, D.; Venzlaff, H.; Garrelfs, J.; Dinh, H.T.; Meyer, V.; Mayrhofer, K.; Hassel, A.W.; Stratmann, M.; Widdel, F. Marine sulfate-reducing bacteria cause serious corrosion of iron under electroconductive biogenic mineral crust. Environ. Microbiol. 2012, 14, 1772–1787. [Google Scholar] [CrossRef]
- Lohner, S.T.; Deutzmann, J.S.; Logan, B.E.; Leigh, J.; Spormann, A.M. Hydrogenase-independent uptake and metabolism of electrons by the archaeon Methanococcus maripaludis. ISME J. 2014, 8, 1673–1681. [Google Scholar] [CrossRef]
- Dinh, H.T.; Kuever, J.; Mußmann, M.; Hassel, A.W.; Stratmann, M.; Widdel, F. Iron corrosion by novel anaerobic microorganisms. Nature 2004, 427, 829–832. [Google Scholar] [CrossRef]
- Deutzmann, J.S.; Sahin, M.; Spormann, A.M. Extracellular Enzymes Facilitate Electron Uptake in Biocorrosion and Bioelectrosynthesis. MBio 2015, 6, e00496-15. [Google Scholar] [CrossRef]
- Kato, S. Microbial extracellular electron transfer and its relevance to iron corrosion. Microb. Biotechnol. 2016, 9, 141–148. [Google Scholar] [CrossRef]
- Palacios, P.A.; Snoeyenbos-West, O.; Löscher, C.R.; Thamdrup, B.; Rotaru, A.E. Baltic Sea methanogens compete with acetogens for electrons from metallic iron. ISME J. 2019, 13, 3011–3023. [Google Scholar] [CrossRef]
- Venzlaff, H.; Enning, D.; Srinivasan, J.; Mayrhofer, K.J.J.; Hassel, A.W.; Widdel, F.; Stratmann, M. Accelerated cathodic reaction in microbial corrosion of iron due to direct electron uptake by sulfate-reducing bacteria. Corros. Sci. 2013, 66, 88–96. [Google Scholar] [CrossRef]
- Sharma, M.; Bajracharya, S.; Gildemyn, S.; Patil, S.A.; Alvarez-Gallego, Y.; Pant, D.; Rabaey, K.; Dominguez-Benetton, X. A critical revisit of the key parameters used to describe microbial electrochemical systems. Electrochim. Acta 2014, 140, 191–208. [Google Scholar] [CrossRef]
- Vigneron, A.; Head, I.M.; Tsesmetzis, N. Damage to offshore production facilities by corrosive microbial biofilms. Appl. Microbiol. Biotechnol. 2018, 102, 2525–2533. [Google Scholar] [CrossRef] [PubMed]
- Philips, J.; Van den Driessche, N.; De Paepe, K.; Prévoteau, A.; Gralnick, J.A.; Arends, J.B.A.; Rabaey, K. A novel Shewanella isolate enhances corrosion by using metallic iron as the electron donor with fumarate as the electron acceptor. Appl. Environ. Microbiol. 2018, 84, e01154-18. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.Y.; Holmes, D.E.; Ueki, T.; Palacios, P.A.; Lovley, D.R. Iron corrosion via direct metal-microbe electron transfer. MBio 2019, 10, e00303-19. [Google Scholar] [CrossRef] [PubMed]
- Philips, J.; Monballyu, E.; Georg, S.; De Paepe, K.; Prévoteau, A.; Rabaey, K.; Arends, J.B.A. An Acetobacterium strain isolated with metallic iron as electron donor enhances iron corrosion by a similar mechanism as Sporomusa sphaeroides. FEMS Microbiol. Ecol. 2019, 95, fiy222. [Google Scholar] [CrossRef]
- Mehanna, M.; Basséguy, R.; Délia, M.L.; Bergel, A. Effect of Geobacter sulfurreducens on the microbial corrosion of mild steel, ferritic and austenitic stainless steels. Corros. Sci. 2009, 51, 2596–2604. [Google Scholar] [CrossRef]
- Pirbadian, S.; Barchinger, S.E.; Leung, K.M.; Byun, H.S.; Jangir, Y.; Bouhenni, R.A.; Reed, S.B.; Romine, M.F.; Saffarini, D.A.; Shi, L.; et al. Shewanella oneidensis MR-1 nanowires are outer membrane and periplasmic extensions of the extracellular electron transport components. Proc. Natl. Acad. Sci. USA 2014, 111, 12883–12888. [Google Scholar] [CrossRef]
- Gorby, Y.A.; Yanina, S.; McLean, J.S.; Rosso, K.M.; Moyles, D.; Dohnalkova, A.; Beveridge, T.J.; Chang, I.S.; Kim, B.H.; Kim, K.S.; et al. Electrically conductive bacterial nanowires produced by Shewanella oneidensis strain MR-1 and other microorganisms. Proc. Natl. Acad. Sci. USA 2006, 103, 11358–11363. [Google Scholar] [CrossRef]
- Okamoto, A.; Nakamura, R.; Hashimoto, K. Electrochimica Acta In-vivo identification of direct electron transfer from Shewanella oneidensis MR-1 to electrodes via outer-membrane OmcA—MtrCAB protein complexes. Electrochim. Acta 2011, 56, 5526–5531. [Google Scholar] [CrossRef]
- Reguera, G.; McCarthy, K.D.; Mehta, T.; Nicoll, J.S.; Tuominen, M.T.; Lovley, D.R. Extracellular electron transfer via microbial nanowires. Nature 2005, 435, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Summers, Z.M.; Fogarty, H.E.; Leang, C.; Franks, A.E.; Malvankar, N.S.; Lovley, D.R. Direct exchange of electrons within aggregates of an evolved syntrophic coculture of anaerobic bacteria. Science 2010, 330, 1413–1415. [Google Scholar] [CrossRef] [PubMed]
- Vargas, M.; Malvankar, N.S.; Tremblay, P.L.; Leang, C.; Smith, J.A.; Patel, P.; Synoeyenbos-West, O.; Nevin, K.P.; Lovley, D.R. Aromatic amino acids required for pili conductivity and long-range extracellular electron transport in Geobacter sulfurreducens. MBio 2013, 4, e00105-13. [Google Scholar] [CrossRef]
- Tremblay, P.L.; Summers, Z.M.; Glaven, R.H.; Nevin, K.P.; Zengler, K.; Barrett, C.L.; Qiu, Y.; Palsson, B.O.; Lovley, D.R. A c-type cytochrome and a transcriptional regulator responsible for enhanced extracellular electron transfer in Geobacter sulfurreducens revealed by adaptive evolution. Environ. Microbiol. 2011, 13, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Reguera, G.; Nevin, K.P.; Nicoll, J.S.; Covalla, S.F.; Woodard, T.L.; Lovley, D.R. Biofilm and nanowire production leads to increased current in Geobacter sulfurreducens fuel cells. Appl. Environ. Microbiol. 2006, 72, 7345–7348. [Google Scholar] [CrossRef] [PubMed]
- Shoesmith, D.W.; Mitlin, S.; Power, I.M.; Sherar, B.W.A.; Southam, G.; Keech, P.G. Characterizing the effect of carbon steel exposure in sulfide containing solutions to microbially induced corrosion. Corros. Sci. 2010, 53, 955–960. [Google Scholar] [CrossRef]
- Heydorn, R.L.; Engel, C.; Krull, R.; Dohnt, K. Strategies for the Targeted Improvement of Anodic Electron Transfer in Microbial Fuel Cells. ChemBioEng Rev. 2020, 7, 4–17. [Google Scholar] [CrossRef]
- Chen, J.J.; Chen, W.; He, H.; Li, D.B.; Li, W.W.; Xiong, L.; Yu, H.Q. Manipulation of microbial extracellular electron transfer by changing molecular structure of phenazine-type redox mediators. Environ. Sci. Technol. 2013, 47, 1033–1039. [Google Scholar] [CrossRef]
- Min, D.; Cheng, L.; Zhang, F.; Huang, X.N.; Li, D.B.; Liu, D.F.; Lau, T.C.; Mu, Y.; Yu, H.Q. Enhancing Extracellular Electron Transfer of Shewanella oneidensis MR-1 through Coupling Improved Flavin Synthesis and Metal-Reducing Conduit for Pollutant Degradation. Environ. Sci. Technol. 2017, 51, 5082–5089. [Google Scholar] [CrossRef]
- Qiao, Y.J.; Qiao, Y.; Zou, L.; Wu, X.S.; Liu, J.H. Biofilm promoted current generation of Pseudomonas aeruginosa microbial fuel cell via improving the interfacial redox reaction of phenazines. Bioelectrochemistry 2017, 117, 34–39. [Google Scholar] [CrossRef]
- Jia, R.; Yang, D.; Xu, D.; Gu, T. Electron transfer mediators accelerated the microbiologically influence corrosion against carbon steel by nitrate reducing Pseudomonas aeruginosa biofilm. Bioelectrochemistry 2017, 118, 38–46. [Google Scholar] [CrossRef]
- Huang, L.; Tang, J.; Chen, M.; Liu, X.; Zhou, S. Two modes of riboflavin-mediated extracellular electron transfer in Geobacter uraniireducens. Front. Microbiol. 2018, 9, 2886. [Google Scholar] [CrossRef] [PubMed]
- Brutinel, E.D.; Gralnick, J.A. Shuttling happens: Soluble flavin mediators of extracellular electron transfer in Shewanella. Appl. Microbiol. Biotechnol. 2012, 93, 41–48. [Google Scholar] [CrossRef]
- Booth, G.H.; Tiller, A.K. Polarization studies of mild steel in cultures of sulphate-reducing bacteria. Trans. Faraday Soc. 1960, 56, 1689–1696. [Google Scholar] [CrossRef]
- Schröder, U. Anodic electron transfer mechanisms in microbial fuel cells and their energy efficiency. Phys. Chem. Chem. Phys. 2007, 9, 2619–2629. [Google Scholar] [CrossRef]
- Eaktasang, N.; Kang, C.S.; Lim, H.; Kwean, O.S.; Cho, S.; Kim, Y.; Kim, H.S. Production of electrically-conductive nanoscale filaments by sulfate-reducing bacteria in the microbial fuel cell. Bioresour. Technol. 2016, 210, 61–67. [Google Scholar] [CrossRef]
- Khan, M.S.; Yang, C.; Zhao, Y.; Pan, H.; Zhao, J.; Shahzad, M.B.; Kolawole, S.K.; Ullah, I.; Yang, K. An induced corrosion inhibition of X80 steel by using marine bacterium Marinobacter salsuginis. Colloids Surf. B Biointerfaces 2020, 189, 110858. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.S.; Liang, T.; Liu, Y.; Shi, Y.; Zhang, H.; Li, H.; Guo, S.; Pan, H.; Yang, K.; Zhao, Y. Microbiologically Influenced Corrosion Mechanism of Ferrous Alloys in Marine Environment. Metals 2022, 12, 1458. https://doi.org/10.3390/met12091458
Khan MS, Liang T, Liu Y, Shi Y, Zhang H, Li H, Guo S, Pan H, Yang K, Zhao Y. Microbiologically Influenced Corrosion Mechanism of Ferrous Alloys in Marine Environment. Metals. 2022; 12(9):1458. https://doi.org/10.3390/met12091458
Chicago/Turabian StyleKhan, M. Saleem, Tao Liang, Yuzhi Liu, Yunzhu Shi, Huanhuan Zhang, Hongyu Li, Shifeng Guo, Haobo Pan, Ke Yang, and Ying Zhao. 2022. "Microbiologically Influenced Corrosion Mechanism of Ferrous Alloys in Marine Environment" Metals 12, no. 9: 1458. https://doi.org/10.3390/met12091458