Influence of Sports Training in Foothills on the Professional Athlete’s Immunity

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
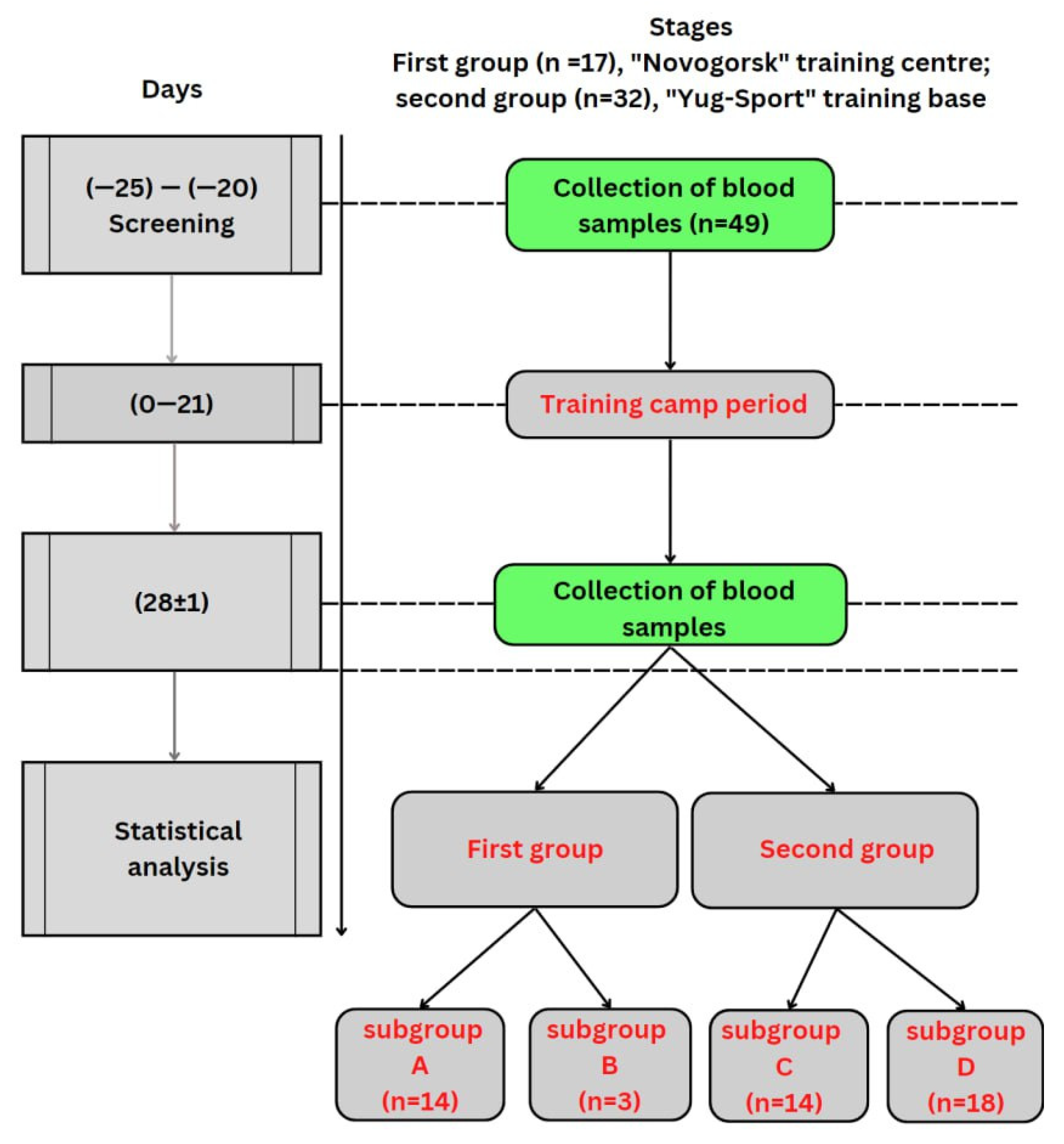
2. Materials and Methods
2.1. Demography
2.2. Study Design
2.3. Laboratory
2.4. Statistics
2.5. Ethics
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pustovojt, V.; Klyuchnikov, M.; Nazaryan, S.; Eroyan, I.; Samojlov, A. Heart rate variability as the main method of assessing the functional state of athletes participating in extreme sports. Mod. Issues Biomed. 2021, 5, 40. [Google Scholar] [CrossRef] [PubMed]
- Samoilov, A.S.; Nikonov, R.V.; Pustovoit, V.I. Stress in Extreme Occupational Activity: A Monograph, State Research Center—Burnasyan Federal Medical Biophysical Center of Federal Medical Biological Agency; SSC FMBC: Moscow, Russia, 2022. [Google Scholar]
- Ding, J.X.; Rudak, P.T.; Inoue, W.; Haeryfar, S.M. Physical restraint mouse models to assess immune responses under stress with or without habituation. STAR Protoc. 2021, 2, 100838. [Google Scholar] [CrossRef]
- Rudak, P.T.; Gangireddy, R.; Choi, J.; Burhan, A.M.; Summers, K.L.; Jackson, D.N.; Inoue, W.; Haeryfar, S.M. Stress-elicited glucocorticoid receptor signaling upregulates TIGIT in innate-like invariant T lymphocytes. Brain Behav. Immun. 2019, 80, 793–804. [Google Scholar] [CrossRef]
- Rudak, P.T.; Choi, J.; Parkins, K.M.; Summers, K.L.; Jackson, D.N.; Foster, P.J.; Skaro, A.I.; Leslie, K.; McAlister, V.C.; Kuchroo, V.K.; et al. Chronic stress physically spares but functionally impairs innate-like invariant T cells. Cell Rep. 2021, 35, 108979. [Google Scholar] [CrossRef] [PubMed]
- Sarin, H.V.; Gudelj, I.; Honkanen, J.; Ihalainen, J.K.; Vuorela, A.; Lee, J.H.; Jin, Z.; Terwilliger, J.D.; Isola, V.; Ahtiainen, J.P.; et al. Molecular Pathways Mediating Immunosuppression in Response to Prolonged Intensive Physical Training, Low-Energy Availability, and Intensive Weight Loss. Front. Immunol. 2019, 10, 907. [Google Scholar] [CrossRef] [Green Version]
- Dalvie, S.; Chatzinakos, C.; Al Zoubi, O.; Georgiadis, F.; Lancashire, L.; Daskalakis, N.P. From genetics to systems biology of stress-related mental disorders. Neurobiol. Stress 2021, 15, 100393. [Google Scholar] [CrossRef] [PubMed]
- Flow Cytofluorimeter BD FACSCantoTM. II. Available online: https://bioline.ru// (accessed on 29 December 2021).
- IMK Kit. Available online: https://www.bdbiosciences.com/en-eu/products/reagents/flow-cytometry-reagents/clinical-diagnostics/multicolor-cocktails-and-kits-ivd-ce-ivds/imk-kit.340503 (accessed on 29 December 2021).
- Wójcik, R. The Effect of Leiber Beta-S (1,3-1,6-β-D-Glucan) on the Phagocytic Activity and Oxidative Metabolism of Peripheral Blood Granulocytes and Monocytes in Calves. Acta Vet. Brno 2014, 83, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Glycotope|Clinical-Stage Immuno-Oncology Company in Berlin. Glycotope. Available online: https://www.glycotope.com/ (accessed on 29 December 2021).
- Modular Platform Cobas® 6000. Available online: https://diagnostics.roche.com/ru/ru/products/systems/cobas_-6000-analyzer-series.html (accessed on 22 June 2022).
- Romero-Garcia, S.; Moreno-Altamirano, M.M.B.; Prado-Garcia, H.; Sánchez-García, F.J. Lactate Contribution to the Tumor Microenvironment: Mechanisms, Effects on Immune Cells and Therapeutic Relevance. Front. Immunol. 2016, 7, 52. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.Y.; Huynh, H.; Chen, P.; Peña-Llopis, S.; Wan, Y. Macrophage PPARγ inhibits Gpr132 to mediate the anti-tumor effects of rosiglitazone. eLife 2016, 5, e18501. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, J.; Dong, R.; Ji, X.; Jiang, Y.; Cen, J.; Bai, Z.; Hong, K.; Li, H.; Chen, J.; et al. Lactate as a metabolite from probiotic Lactobacilli mitigates ethanol-induced gastric mucosal injury: An in vivo study. BMC Complement. Med. Ther. 2021, 21, 26. [Google Scholar] [CrossRef]
- Petrovsky, D.V.; Pustovoyt, V.I.; Nikolsky, K.S.; Malsagova, K.A.; Kopylov, A.T.; Stepanov, A.A.; Rudnev, V.R.; Balakin, E.I.; Kaysheva, A.L. Tracking Health, Performance and Recovery in Athletes Using Machine Learning. Sports 2022, 10, 160. [Google Scholar] [CrossRef] [PubMed]
- Pustovoit, V.I.; Samoilov, A.S.; Nikonov, R.V. Divers’ infectious pathologies in severe climate. Sports Med. Res. Pract. 2020, 1, 67–75. [Google Scholar] [CrossRef]
- Wculek, S.K.; Khouili, S.C.; Priego, E.; Heras-Murillo, I.; Sancho, D. Metabolic Control of Dendritic Cell Functions: Digesting Information. Front. Immunol. 2019, 10, 775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazon, A.; Tyrakis, P.A.; Macias, D.; Veliça, P.; Rundqvist, H.; Fitzpatrick, S.; Vojnovic, N.; Phan, A.T.; Loman, N.; Hedenfalk, I.; et al. An HIF-1α/VEGF-A Axis in Cytotoxic T Cells Regulates Tumor Progression. Cancer Cell 2017, 32, 669–683.e5. [Google Scholar] [CrossRef] [Green Version]
- Rollings, C.M.; Sinclair, L.V.; Brady, H.J.M.; Cantrell, D.A.; Ross, S.H. Interleukin-2 shapes the cytotoxic T cell proteome and immune environment–sensing programs. Sci. Signal. 2018, 11, eaap8112. [Google Scholar] [CrossRef] [Green Version]
- Phan, A.T.; Doedens, A.L.; Palazon, A.; Tyrakis, P.A.; Cheung, K.P.; Johnson, R.S.; Goldrath, A.W. Constitutive Glycolytic Metabolism Supports CD8+ T Cell Effector Memory Differentiation during Viral Infection. Immunity 2016, 45, 1024–1037. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, Y.; Atakan, M.; Kuang, J.; Hu, Y.; Bishop, D.; Yan, X. The Molecular Adaptive Responses of Skeletal Muscle to High-Intensity Exercise/Training and Hypoxia. Antioxidants 2020, 9, 656. [Google Scholar] [CrossRef]
- Ho, J.D.; Schatz, J.H.; Uniacke, J.; Lee, S. Jekyll and Hyde: Activating the Hypoxic Translational Machinery. Trends Biochem. Sci. 2020, 46, 171–174. [Google Scholar] [CrossRef]
- Masson, N.; Keeley, T.P.; Giuntoli, B.; White, M.D.; Lavilla Puerta, M.; Perata, P.; Hopkinson, R.J.; Flashman, E.; Licausi, F.; Ratcliffe, P.J. Conserved N-terminal cysteine dioxygenases transduce responses to hypoxia in animals and plants. Science 2019, 365, 65–69. [Google Scholar] [CrossRef]
- Strowitzki, M.J.; Cummins, E.P.; Taylor, C.T. Protein Hydroxylation by Hypoxia-Inducible Factor (HIF) Hydroxylases: Unique or Ubiquitous? Cells 2019, 8, 384. [Google Scholar] [CrossRef]
- Dimeloe, S.; Gubser, P.; Loeliger, J.; Frick, C.; Develioglu, L.; Fischer, M.; Marquardsen, F.; Bantug, G.R.; Thommen, D.; Lecoultre, Y.; et al. Tumor-derived TGF-β inhibits mitochondrial respiration to suppress IFN-γ production by human CD4 + T cells. Sci. Signal. 2019, 12, eaav3334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caslin, H.L.; Abebayehu, D.; Qayum, A.A.; Haque, T.T.; Taruselli, M.T.; Paez, P.A.; Pondicherry, N.; Barnstein, B.O.; Hoeferlin, L.A.; Chalfant, C.E.; et al. Lactic Acid Inhibits Lipopolysaccharide-Induced Mast Cell Function by Limiting Glycolysis and ATP Availability. J. Immunol. 2019, 203, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Kiran, D.; Basaraba, R.J. Lactate Metabolism and Signaling in Tuberculosis and Cancer: A Comparative Review. Front. Cell. Infect. Microbiol. 2021, 11, 624607. [Google Scholar] [CrossRef] [PubMed]
- Abebayehu, D.; Spence, A.J.; Qayum, A.A.; Taruselli, M.T.; McLeod, J.J.A.; Caslin, H.L.; Chumanevich, A.P.; Kolawole, E.M.; Paranjape, A.; Baker, B.; et al. Lactic Acid Suppresses IL-33–Mediated Mast Cell Inflammatory Responses via Hypoxia-Inducible Factor-1α–Dependent miR-155 Suppression. J. Immunol. 2016, 197, 2909–2917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatib-Massalha, E.; Bhattacharya, S.; Massalha, H.; Biram, A.; Golan, K.; Kollet, O.; Kumari, A.; Avemaria, F.; Petrovich-Kopitman, E.; Gur-Cohen, S.; et al. Lactate released by inflammatory bone marrow neutrophils induces their mobilization via endothelial GPR81 signaling. Nat. Commun. 2020, 11, 3547. [Google Scholar] [CrossRef]
- Yan, L.; Tan, Y.; Chen, G.; Fan, J.; Zhang, J. Harnessing Metabolic Reprogramming to Improve Cancer Immunotherapy. Int. J. Mol. Sci. 2021, 22, 10268. [Google Scholar] [CrossRef]
- Suzuki, K.; Hayashida, H. Effect of Exercise Intensity on Cell-Mediated Immunity. Sports 2021, 9, 8. [Google Scholar] [CrossRef]
- Suzuki, K. Recent Progress in Applicability of Exercise Immunology and Inflammation Research to Sports Nutrition. Nutrients 2021, 13, 4299. [Google Scholar] [CrossRef]
- Beshgetoor, D.; Arrues, S.; McGuire, K. Effect of Competitive Training on T-cell Mediated Immune Function in Master’s Female Athletes. Int. J. Sports Med. 2004, 25, 553–558. [Google Scholar] [CrossRef]
- Howden, A.J.M.; Hukelmann, J.L.; Brenes, A.; Spinelli, L.; Sinclair, L.V.; Lamond, A.I.; Cantrell, D.A. Quantitative analysis of T cell proteomes and environmental sensors during T cell differentiation. Nat. Immunol. 2019, 20, 1542–1554. [Google Scholar] [CrossRef]
- Johnson, M.O.; Siska, P.J.; Contreras, D.C.; Rathmell, J.C. Nutrients and the microenvironment to feed a T cell army. Semin. Immunol. 2016, 28, 505–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voss, K.; Hong, H.S.; Bader, J.E.; Sugiura, A.; Lyssiotis, C.A.; Rathmell, J.C. A Guide to Interrogating Immunometabolism. Nat. Rev. Immunol. 2022, 21, 637–652. [Google Scholar] [CrossRef] [PubMed]
- Geltink, R.I.K.; Kyle, R.L.; Pearce, E.L. Unraveling the Complex Interplay Between T Cell Metabolism and Function. Annu. Rev. Immunol. 2018, 36, 461–488. [Google Scholar] [CrossRef] [PubMed]
- Gropper, Y.; Feferman, T.; Shalit, T.; Salame, T.-M.; Porat, Z.; Shakhar, G. Culturing CTLs under Hypoxic Conditions Enhances Their Cytolysis and Improves Their Anti-tumor Function. Cell Rep. 2017, 20, 2547–2555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murthy, A.; Gerber, S.A.; Koch, C.J.; Lord, E.M. Intratumoral Hypoxia Reduces IFN-γ–Mediated Immunity and MHC Class I Induction in a Preclinical Tumor Model. Immunohorizons 2019, 3, 149–160. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-N.; Yang, J.-F.; Huang, D.-J.; Ni, H.-H.; Zhang, C.-X.; Zhang, L.; He, J.; Gu, J.-M.; Chen, H.-X.; Mai, H.-Q.; et al. Hypoxia Induces Mitochondrial Defect That Promotes T Cell Exhaustion in Tumor Microenvironment Through MYC-Regulated Pathways. Front. Immunol. 2020, 11, 1906. [Google Scholar] [CrossRef]
- Yu, Y.-R.; Imrichova, H.; Wang, H.; Chao, T.; Xiao, Z.; Gao, M.; Rincon-Restrepo, M.; Franco, F.; Genolet, R.; Cheng, W.-C.; et al. Disturbed mitochondrial dynamics in CD8+ TILs reinforce T cell exhaustion. Nat. Immunol. 2020, 21, 1540–1551. [Google Scholar] [CrossRef]
- Scharping, N.E.; Rivadeneira, D.B.; Menk, A.V.; Vignali, P.D.A.; Ford, B.R.; Rittenhouse, N.L.; Peralta, R.; Wang, Y.; Wang, Y.; DePeaux, K.; et al. Mitochondrial stress induced by continuous stimulation under hypoxia rapidly drives T cell exhaustion. Nat. Immunol. 2021, 22, 205–215. [Google Scholar] [CrossRef]


{kind=link}
| Cell Type | Norm (%) | Norm (109/L) |
|---|---|---|
| Leukocytes | - | 4.0–9.0 |
| Lymphocytes | 19.0–37.0 | 1.2–3.0 |
| Stab neutrophils | 1.0–6.0 | - |
| Segmented neutrophils | 47.0–72.0 | - |
| Eosinophils | 0.5–5.0 | 0.1–0.3 |
| Basophils | 0–1.0 | 0–0.07 |
| Monocytes | 3.0–11.0 | 0.1–0.9 |
| T-lymphocytes CD45+ CD3+ | 55.0–80.0 | 0.95–1.8 |
| T-helpers CD45+ CD3+ CD4+ | 31.0–51.0 | 0.57–1.1 |
| T-killers (CTL) CD45+ CD3+ CD8+ | 19.0–35.0 | 0.45–0.85 |
| NK cells CD45+ CD3− CD(16 + 56)+ | 7.0–20.0 | 0.18–0.42 |
| B- lymphocytes CD45+ CD3− CD19+ | 6.0–19.0 | 0.15–0.4 |
| Index CD4+/CD8+ | 1.5–2.0 | - |
| Phagocytic activity of blood granulocytes | 55.0–95.0 | - |
| Parameter | Norm | Measure |
|---|---|---|
| Immunoglobulin class G (IgG) | 7.0–16.0 | g/L |
| Immunoglobulin class A (IgA) | 0.7–4.0 | g/L |
| Immunoglobulin class M (IgM) | 0.4–2.3 | g/L |
| Immunoglobulin class E (IgE) | 1.31–165.3 | IU/mL |
| Interferon alpha (IFN-α) | 640.0–1280.0 | IU/mL |
| Interferon gamma (IFN-γ) | 128.0–256.0 | IU/mL |
| Parameters | Baseline Examination, (n = 17) | Day 7, Subgroup A (n = 14) | Day 7 Subgroup B (n = 3) |
|---|---|---|---|
| Leukocytes (109/L) | 6.9 [6.3–7.6] | 7.3 [6.2–8.4] | 6.2 [3.9–8.5] |
| Lymphocytes (109/L) | 2.2 [2.1–2.4] | 2.4 [2.1–2.6] *,** | 1.8 [1.7–1.9] *,** |
| T-lymphocytes (109/L) | 1.6 [1.4–1.7] | 1.7 [1.5–1.9] | 1.2 [0.7–1.7] |
| Leukocytes/T-lymphocytes (109/L) | 4.5 [4.0–5.0] | 4.3 [3.6–5.0] | 5.1 [4.2–5.6] |
| CD3/4+, (109/L) | 0.9 [0.9–1.0] | 1 [0.9–1.1] | 0.7 [0.4–1.1] |
| CD3/8+, (109/L) | 0.6 [0.5–0.7] | 0.6 [0.5–0.8] | 0.4 [0.2–0.6] |
| CD19+, (109/L) | 0.3 [0.2–0.3] | 0.3 [0.2–0.4] | 0.2 [0–0.4] |
| CD16/56+, (109/L) | 0.4 [0.3–0.4] | 0.4 [0.3–0.4] | 0.3 [0.2–0.6] |
| CD3/16/56+, (109/L) | 6.9 [6.3–7.6] | 7.3 [6.2–8.4] | 6.2 [3.9–8.5] |
| Parameter | Baseline Examination (n = 17) | Day 7, Subgroup A (n = 14) | Day 7 Second Subgroup B (n = 3) |
|---|---|---|---|
| Stab neutrophils (%) | 53.8 [50.5–57.2] | 53.4 [47.4–59.3] | 56.3 [46.3–66.2] |
| Segmented neutrophils (%) | 2.4 [1.8–3.1] | 2.1 [1.2–2.9] | 4.0 [1.6–5.6] |
| Eosinophils (%) | 0.7 [0.6–0.9] | 0.7 [0.5–0.9] | 0.6 [0.2–1.1] |
| Basophils (%) | 9.7 [8.8–10.6] | 9.7 [8.2–11.1] | 9.7 [1.6–17.8] |
| Monocytes (%) | 32.8 [30.1–35.4] | 33 [28.7–37.3] | 32.1 [23.1–41.0] |
| Lymphocytes (%) | 71.2 [68.8–73.6] | 71.6 [68.5–74.7] | 68.1 [41.9–94.3] |
| CD3+, (%) | 42.1 [39.6–44.7] | 42.0 [38.4–45.5] | 40.8 [19.6–61.8] |
| CD3/4+, (%) | 26.2 [24–28.5] | 26.8 [23.1–30.4] | 23.4 [13.8–33.1] |
| CD3/8+, (%) | 1.7 [1.52–1.9] | 1.7 [1.3–2.0] | 1.7 [0.96–2.53] |
| Index CD4/CD8+ (%) | 12.0 [10.6–13.4] | 13.2 [10.6–15.8] | 11.3 [3.7–18.9] |
| CD19+ (%) | 16.1 [13.4–18.7] | 14.5 [11.0–18.0] | 19.0 [10.0–33.8] |
| CD16/56+ (%) | 52.0 [43.6–60.5] | 50.5 [41.9–59.1] | 58.8 [37.9–85.3] |
| CD3/16/56+ (%) | 53.8 [50.5–57.2] | 53.4 [47.4–59.3] | 56.3 [46.3–66.2] |
| Phagocytic activity of blood granulocytes (%) | 2.4 [1.8–3.1] | 2.1 [1.2–2.9] | 4.0 [1.6–5.6] |
| Parameter | Baseline Examination (n = 17) | Day 7, Subgroup A (n = 14) | Day 7 Subgroup B (n = 3) |
|---|---|---|---|
| IgG (g/L) | 11.8 [11.0–12.6] | 11.4 [10.2–12.6] | 15.2 [9.9–17.5] |
| IgA (g/L) | 2.2 [2.0–2.4] | 2.3 [1.9–2.6] | 2.1 [1.5–2.4] |
| IgM (g/L) | 1.0 [0.92–1.2] | 1.0 [0.87–1.15] | 1.0 [0.6–1.5] |
| IgE (IU/mL) | 69.3 [44.0–91.0] | 35.2 [17.5–52.8] | 36.2 [7.9–50.6] |
| Parameters | Baseline Examination (n = 32) | Day 7, Subgroup C (n = 14) | Day 7 Subgroup D (n = 18) |
|---|---|---|---|
| Leukocytes (109/L) | 7.0 [6.4–7.7] | 8.2 [7.6–8.9] *,** | 6.7 [6.1–7.5]* |
| Lymphocytes (109/L) | 2.3 [2.1–2.6] | 3.0 [2.8–3.3] *,** | 2 [1.7–2.2] *,** |
| T-lymphocytes (109/L) | 1.7 [1.6–1.9] | 2.2 [2–2.4] * | 1.4 [1.2–1.5] *,** |
| Leukocytes/T-lymphocytes (109/L) | 4.3 [3.8–4.6] | 3.9 [3.5–4.3] * | 5.1 [4.6–5.5] *,** |
| CD3/4+, (109/L) | 1.0 [0.9–1.1] | 1.2 [1.1–1.3] *,** | 0.8 [0.7–0.8] *,** |
| CD3/8+, (109/L) | 0.6 [0.5–0.7] | 0.9 [0.8–0.9] *,** | 0.5 [0.5–0.6] * |
| CD19+, (109/L) | 0.3 [0.3–0.4] | 0.4 [0.3–0.5] * | 0.3 [0.29–0.34] * |
| CD3/16/56+, (109/L) | 0.3 [0.2–0.3] | 0.3 [0.3–0.4] * | 0.2 [0.2–0.3] * |
| Parameter | Baseline Examination (n = 32) | Day 7, Subgroup C (n = 14) | Day 7 Subgroup D (n = 18) |
|---|---|---|---|
| Stab neutrophils (%) | 53.5 [50.4–56.6] | 51.4 [47.4–55.3] * | 57.4 [55.2–61.6] * |
| Segmented neutrophils (%) | 2.4 [1.7–3.2] | 3.0 [2.1–3.9] | 2.8 [1.8–3.6] |
| Eosinophils (%) | 0.2 [0–0.3] | 0.3 [0.2–0.5] | 0.2 [0–0.3] |
| Basophils (%) | 8.8 [8.2–9.5] | 8.5 [7.9–9.1] | 8.4 [7.7–9.1] |
| Monocytes (%) | 34.4 [31.1–37.6] | 35.7 [33.3–42.3] * | 28.9 [26.4–31] *,** |
| Lymphocytes (%) | 73.3 [70.5–76.1] | 73.6 [71.8–75.4] | 71.0 [69–72.9] |
| CD3+, (%) | 42.7 [40.7–44.6] | 41.5 [39.5–43.5] * | 38.9 [37–40.7] *,** |
| CD3/4+, (%) | 26.4 [25.0–27.8] | 27.9 [26.3–29.5] | 27.2 [25.4–28.9] |
| CD3/8+, (%) | 1.7 [1.5–1.8] | 1.6 [1.4–1.8] | 1.5 [1.4–1.7] |
| Index CD4/CD8+ (%) | 14.0 [12.6–15.4] | 14.2 [12.8–15.5] * | 17.0 [15.6–18.5] *,** |
| CD19+ (%) | 10.3 [8.5–12.1] | 10.4 [8.7–12.1] | 10.8 [9.4–12.2] |
| CD16/56+ (%) | 66.3 [62.0–70.4] | 63.7 [59.8–67.7] | 64.4 [59.9–68.8] |
| CD3/16/56+ (%) | 53.5 [50.4–56.6] | 51.4 [47.4–55.3] * | 57.4 [55.2–61.6] * |
| Phagocytic activity of blood granulocytes (%) | 2.4 [1.7–3.2] | 3.0 [2.1–3.9] | 2.8 [1.8–3.6] |
| Parameter | Baseline Examination (n = 17) | Day 7, Group C (n = 14) | Day 7 Group D (n = 3) |
|---|---|---|---|
| IgG (g/L) | 11.0 [10.3–11.7] | 11.1 [10.4–11.7] | 10.2 [9.5–10.8] |
| IgA (g/L) | 1.5 [1.2–1.7] | 1.4 [1.2–1.6] | 1.2 [0.9–1.5] |
| IgM (g/L) | 1.0 [0.9–1.12] | 0.91 [0.79–1] | 0.8 [0.7–1] |
| IgE (IU/mL) | 69.3 [44.0–91.0] | 77.0 [47.7–96.4] * | 36.1 [24.8–47.3] * |
| IFN-α (IU/mL) | 753.0 [585.0–921.0] | 582.0 [471.0–693.0] | 567.0 [447.0–714.0] |
| IFN-γ (IU/mL) | 111.3 [95.9–128.1] | 122.0 [103.6–139.3] * | 86.1 [76.3–95.2] *,** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malsagova, K.A.; Astrelina, T.A.; Balakin, E.I.; Kobzeva, I.V.; Adoeva, E.Y.; Yurku, K.A.; Suchkova, Y.B.; Stepanov, A.A.; Izotov, A.A.; Butkova, T.V.; et al. Influence of Sports Training in Foothills on the Professional Athlete’s Immunity. Sports 2023, 11, 30. https://doi.org/10.3390/sports11020030
Malsagova KA, Astrelina TA, Balakin EI, Kobzeva IV, Adoeva EY, Yurku KA, Suchkova YB, Stepanov AA, Izotov AA, Butkova TV, et al. Influence of Sports Training in Foothills on the Professional Athlete’s Immunity. Sports. 2023; 11(2):30. https://doi.org/10.3390/sports11020030
Chicago/Turabian StyleMalsagova, Kristina A., Tatiana A. Astrelina, Evgenii I. Balakin, Irina V. Kobzeva, Elena Ya. Adoeva, Kseniya A. Yurku, Yuliya B. Suchkova, Alexander A. Stepanov, Alexander A. Izotov, Tatyana V. Butkova, and et al. 2023. "Influence of Sports Training in Foothills on the Professional Athlete’s Immunity" Sports 11, no. 2: 30. https://doi.org/10.3390/sports11020030