

Effects of Training Sets Sequence on Swimming Performance, Training Load and Physiological Responses

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preliminary Tests
2.2. Main Tests
2.3. Measurements
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laursen, P.B. Training for Intense Exercise Performance: High-Intensity or High-Volume Training? Scand. J. Med. Sci. Sports 2010, 20, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hermosilla, F.; González-Rave, J.M.; Del Castillo, J.A.; Pyne, D.B. Periodization and Programming for Individual 400 m Medley Swimmers. Int. J. Environ. Res. Public Health 2021, 18, 6474. [Google Scholar] [CrossRef] [PubMed]
- Skorski, S.; Faude, O.; Urhausen, A.; Kindermann, W.; Meyer, T. Intensity Control in Swim Training by Means of the Individual Anaerobic Threshold. J. Strength Cond. Res. 2012, 26, 3304–3311. [Google Scholar] [CrossRef] [PubMed]
- Dekerle, J.; Brickley, G.; Alberty, M.; Pelayo, P. Characterising the Slope of the Distance-Time Relationship in Swimming. J. Sci. Med. Sport 2010, 13, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Dekerle, J.; Pelayo, P.; Clipet, B.; Depretz, S.; Lefevre, T.; Sidney, M. Critical Swimming Speed Does Not Represent the Speed at Maximal Lactate Steady State. Int. J. Sports Med. 2005, 26, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Kabasakalis, A.; Nikolaidis, S.; Tsalis, G.; Mougios, V. Low-Volume Sprint Interval Swimming Is Sufficient to Increase Blood Metabolic Biomarkers in Master Swimmers. Res. Q. Exerc. Sport 2020, 93, 318–324. [Google Scholar] [CrossRef]
- Kabasakalis, A.; Nikolaidis, S.; Tsalis, G.; Mougios, V. Response of Blood Biomarkers to Sprint Interval Swimming. Int. J. Sports Physiol. Perform. 2020, 15, 1442–1447. [Google Scholar] [CrossRef]
- Toubekis, A.; Adam, G.; Douda, H.; Antoniou, P.; Douroundos, I.; Tokmakidis, S. Repeated Sprint Swimming Performance after Low- or High-Intensity Active and Passive Recoveries. J. Strength Cond. Res. 2011, 25, 109–116. [Google Scholar] [CrossRef]
- Greenwood, J.D.; Moses, G.E.; Bernardino, F.M.; Gaesser, G.A.; Weltman, A. Intensity of Exercise Recovery, Blood Lactate Disappearance, and Subsequent Swimming Performance. J. Sports Sci. 2008, 26, 29–34. [Google Scholar] [CrossRef]
- Parolin, M.L.; Chesley, A.; Matsos, M.P.; Spriet, L.L.; Jones, N.L.; Heigenhauser, G.J.F. Regulation of Skeletal Muscle Glycogen Phosphorylase and PDH during Maximal Intermittent Exercise. Am. J. Physiol. 1999, 277, E890–E900. [Google Scholar] [CrossRef]
- Ktenidis, C.K.; Margaritelis, N.V.; Cherouveim, E.D.; Stergiopoulos, D.C.; Malliou, V.J.; Geladas, N.D.; Nikolaidis, M.G.; Paschalis, V. Priming Exercise Increases Wingate Cycling Peak Power Output. Eur. J. Sport Sci. 2021, 21, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.; Koppo, K.; Burnley, M. Effects of Prior Exercise on Metabolic and Gas Exchange Responses to Exercise. Sport. Med. 2003, 33, 949–971. [Google Scholar] [CrossRef]
- Vigh-Larsen, J.F.; Ørtenblad, N.; Spriet, L.L.; Overgaard, K.; Mohr, M. Muscle Glycogen Metabolism and High-Intensity Exercise Performance: A Narrative Review. Sport. Med. 2021, 51, 1855–1874. [Google Scholar] [CrossRef] [PubMed]
- Nikitakis, I.S.; Bogdanis, G.C.; Paradisis, G.P.; Toubekis, A.G. Physiological Responses and Swimming-Performance Changes Induced by Altering the Sequence of Training Sets. Int. J. Sports Physiol. Perform. 2023, 1, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Buchheit, M.; Laursen, P.B. High-Intensity Interval Training, Solutions to the Programming Puzzle: Part II: Anaerobic Energy, Neuromuscular Load and Practical Applications. Sport. Med. 2013, 43, 927–954. [Google Scholar] [CrossRef] [PubMed]
- Kabasakalis, A.; Tsalis, G.; Zafrana, E.; Loupos, D.; Mougios, V. Effects of Endurance and High-Intensity Swimming Exercise on the Redox Status of Adolescent Male and Female Swimmers. J. Sports Sci. 2014, 32, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Peyrebrune, M.C.; Nevill, M.E.; Donaldson, F.J.; Cosford, D.J. The Effects of Oral Creatine Supplementation on Performance in Single and Repeated Sprint Swimming. J. Sports Sci. 1998, 16, 271–279. [Google Scholar] [CrossRef]
- Lloria-Varella, J.; Koral, J.; Ravel, A.; Féasson, L.; Murias, J.M.; Busso, T. Neuromuscular and Autonomic Function Is Fully Recovered within 24 h Following a Sprint Interval Training Session. Eur. J. Appl. Physiol. 2023, 123, 2317–2329. [Google Scholar] [CrossRef]
- Brooks, G.A. The Science and Translation of Lactate Shuttle Theory. Cell Metab. 2018, 27, 757–785. [Google Scholar] [CrossRef]
- Bangsbo, J. Performance in Sports-With Specific Emphasis on the Effect of Intensified Training. Scand. J. Med. Sci. Sports 2015, 25, 88–99. [Google Scholar] [CrossRef]
- McKay, A.K.A.; Stellingwerff, T.; Smith, E.S.; Martin, D.T.; Mujika, I.; Goosey-Tolfrey, V.L.; Sheppard, J.; Burke, L.M. Defining Training and Performance Caliber: A Participant Classification Framework. Int. J. Sports Physiol. Perform. 2022, 17, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Zacca, R.; Azevedo, R.; Peterson Silveira, R.; Vilas-Boas, J.; Pyne, D.; Castro, F.; Fernandes, R. Comparison of Incremental Intermittent and Time Trial Testing in Age-Group Swimmers. J. Strength Cond. Res. 2019, 33, 801–810. [Google Scholar] [CrossRef] [PubMed]
- Nikitakis, I.S.; Toubekis, A.G. Lactate Threshold Evaluation in Swimmers: The Importance of Age and Method. Int. J. Sports Med. 2021, 42, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Libicz, S.; Roels, B.; Millet, G.P. VO2 Responses to Intermittent Swimming Sets at Velocity Associated with VO2max. Can J. Appl. Physiol. 2005, 30, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Barroso, R.; Salgueiro, D.F.; do Carmo, E.C.; Nakamura, F.Y. The Effects of Training Volume and Repetition Distance on Session Rating of Perceived Exertion and Internal Load in Swimmers. Int. J. Sports Physiol. Perform. 2015, 10, 848–852. [Google Scholar] [CrossRef] [PubMed]
- Manzi, V.; Iellamo, F.; Impellizzeri, F.; D’ottavio, S.; Castagna, C. Relation between Individualized Training Impulses and Performance in Distance Runners. Med. Sci. Sport. Exerc. 2009, 41, 2090–2096. [Google Scholar] [CrossRef]
- Hynynen, E.; Vesterinen, V.; Rusko, H.; Nummela, A. Effects of Moderate and Heavy Endurance Exercise on Nocturnal HRV. Int. J. Sports Med. 2010, 31, 428–432. [Google Scholar] [CrossRef]
- Buchheit, M. Monitoring Training Status with HR Measures: Do All Roads Lead to Rome? Front. Physiol. 2014, 5, 73. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences, 2nd ed.; Mahwah, N., Ed.; Lawrence Erlbaum Associates: New York, NY, USA, 1988; ISBN 0805802835. [Google Scholar]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A Flexible Statistical Power Analysis Program for the Social, Behavioral, and Biomedical Sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Toubekis, A.; Douda, H.; Tokmakidis, S. Influence of Different Rest Intervals during Active or Passive Recovery on Repeated Sprint Swimming Performance. Eur. J. Appl. Physiol. 2005, 93, 694–700. [Google Scholar] [CrossRef]
- Toubekis, A.; Smilios, I.; Bogdanis, G.; Mavridis, G.; Tokmakidis, S. Effect of Different Intensities of Active Recovery on Sprint Swimming Performance. Appl. Physiol. Nutr. Metab. 2006, 31, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Toubekis, A.; Peyrebrune, M.; Lakomy, H.; Nevill, M. Effects of Active and Passive Recovery on Performance during Repeated-Sprint Swimming. J. Sports Sci. 2008, 26, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Goulding, R.P.; Burnley, M.; Wüst, R.C.I. How Priming Exercise Affects Oxygen Uptake Kinetics: From Underpinning Mechanisms to Endurance Performance. Sport. Med. 2023, 53, 959–976. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.; Vilas-Boas, J.P.; Fernandes, R.J. VO2 Kinetics and Metabolic Contributions Whilst Swimming at 95, 100, and 105% of the Velocity at VO2max. Biomed Res. Int. 2014, 2014, 675363. [Google Scholar] [CrossRef]
- Peyrebrune, M.; Toubekis, A.; Lakomy, H.; Nevill, M. Estimating the Energy Contribution during Single and Repeated Sprint Swimming. Scand. J. Med. Sci. Sports 2014, 24, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Ceesay, S.M.; Prentice, A.M.; Day, K.C.; Murgatroyd, P.R.; Goldberg, G.R.; Scott, W.; Spurr, G.B. The Use of Heart Rate Monitoring in the Estimation of Energy Expenditure: A Validation Study Using Indirect Whole-Body Calorimetry. Br. J. Nutr. 1989, 61, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Impellizzeri, F.M.; Marcora, S.M.; Coutts, A.J. Internal and External Training Load: 15 Years On. Int. J. Sports Physiol. Perform. 2019, 14, 270–273. [Google Scholar] [CrossRef]
- Bourdon, P.C.; Cardinale, M.; Murray, A.; Gastin, P.; Kellmann, M.; Varley, M.C.; Gabbett, T.J.; Coutts, A.J.; Burgess, D.J.; Gregson, W.; et al. Monitoring Athlete Training Loads: Consensus Statement. Int. J. Sports Physiol. Perform. 2017, 12, S2-161–S2-170. [Google Scholar] [CrossRef]
- Mujika, I.; Halson, S.; Burke, L.M.; Balagué, G.; Farrow, D. An Integrated, Multifactorial Approach to Periodization for Optimal Performance in Individual and Team Sports. Int. J. Sports Physiol. Perform. 2018, 13, 538–561. [Google Scholar] [CrossRef]



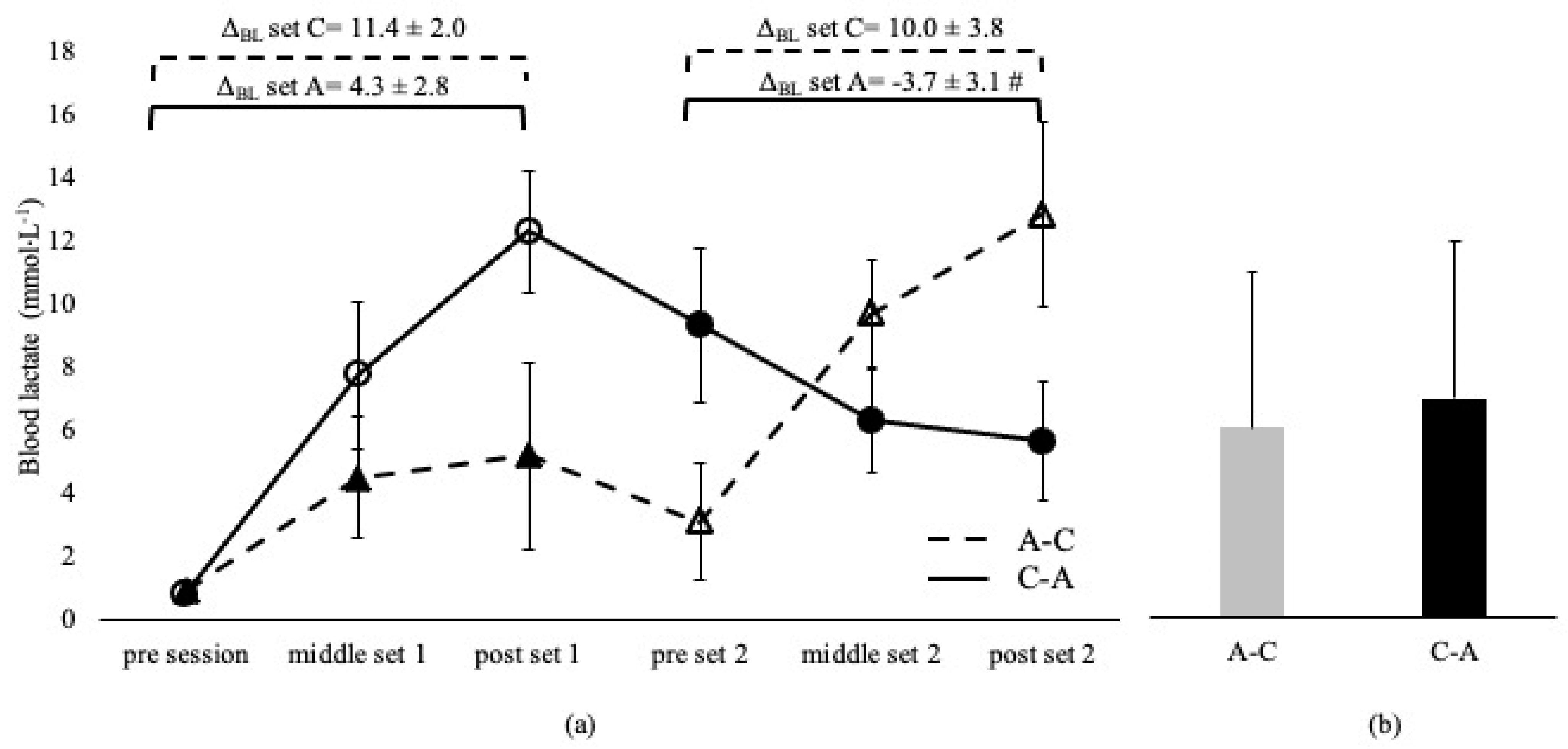{kind=link}
{kind=link}
| Session A-C | Session C-A | |||||
| set A | set C | set C | set A | ES set C | ES set A | |
| Speed (m·s−1) | 1.43 ± 0.08 | 1.71 ± 0.08 | 1.72 ± 0.06 | 1.42 ± 0.07 | 0.08 | 0.21 |
| HR (beats·min−1) | 173 ± 12 | 180 ± 6 | 177 ± 4 | 177 ± 6 | 0.60 | 0.63 |
| TRIMPi (a.u.) | 63.6 ± 40.4 | 18.0 ± 7.7 | 16.4 ± 6.2 | 82.0 ± 45.0 * | 0.23 | 0.43 |
| Session B-C | Session C-B | |||||
| set B | set C | set C | set B | ES set C | ES set B | |
| Speed (m·s−1) | 1.53 ± 0.06 | 1.71 ± 0.07 | 1.71 ± 0.06 | 1.53 ± 0.07 | 0.02 | 0.03 |
| HR (beats·min−1) | 173 ± 11 | 179 ± 7 | 176 ± 6 | 178 ± 6 * | 0.52 | 0.61 |
| TRIMPi (a.u.) | 34.2 ± 20.8 | 19.1 ± 9.1 | 16.4 ± 5.3 | 39.4 ± 14.9 | 0.37 | 0.29 |
| Session A-C | Session C-A | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre-Set A | Post-Set A | Δset A | Pre-Set C | Post-Set C | Δset C | Overall Session | Pre-Set C | Post-Set C | Δset C | Pre-Set A | Post-Set A | Δset A | Overall Session | ES | ||||
| pH | 7.42 ± 0.02 | 7.37 ± 0.05 | −0.05 ± 0.05 | 7.38 ± 0.04 | 7.23 ± 0.05 | −0.15 ± 0.07 | 7.35 ± 0.08 | 7.42 ± 0.01 | 7.20 ± 0.05 | −0.22 ± 0.05 | 7.25 ± 0.07 | 7.36 ± 0.03 | 0.11 ± 0.08 * | 7.31 ± 0.10 # | 0.45 | |||
| BE (mmol·L−1) | 1.8 ± 2.0 | −4.4 ± 4.7 | −6.3 ± 4.0 | −3.2 ± 3.9 | −14.6 ± 3.7 | −11.4 ± 5.2 | −5.1 ± 7.0 | 0.9 ± 1.4 | −15.6 ± 2.5 | −16.5 ± 2.3 * | −13.3 ± 3.9 | −5.7 ± 2.3 | 7.6 ± 4.0 * | −8.4 ± 7.1 # | 0.47 | |||
| HCO3 (mmol·L−1) | 26.1 ± 1.8 | 21.0 ± 3.9 | −5.1 ± 3.2 | 22.2 ± 3.3 | 12.9 ± 2.7 | −9.3 ± 4.0 | 20.5 ± 5.7 | 25.3 ± 1.3 | 12.5 ± 1.8 | −12.8 ± 1.7* | 13.9 ± 2.7 | 19.8 ± 1.9 | 5.9 ± 2.8 * | 17.9 ± 5.5 # | 0.48 | |||
| Ln RMSSD (ms) | 2.7 ± 0.3 | 2.0 ± 0.4 | - | - | 1.9 ± 0.4 | - | 2.2 ± 0.5 | 2.7 ± 0.2 | 1.7 ± 0.4 | - | - | 1.8 ± 0.4 | - | 2.1 ± 0.6 | 0.24 | |||
| RPE (a.u.) | 1.8 ± 1.7 | 6.1 ± 2.2 | 4.3 ± 1.7 | 4.5 ± 2.6 | 9.2 ± 1.1 | 4.7 ± 2.8 | 7.9 ± 0.9 | 2.5 ± 2.2* | 8.8 ± 1.5 | 6.9 ± 2.0 | 4.4 ± 2.3 * | 7.0 ± 2.6 | 2.7 ± 2.7 | 8.1 ± 1.9 | 0.12 | |||
| Session B-C | Session C-B | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre-Set B | Post-Set B | Δset B | Pre-Set C | Post-Set C | Δset C | Overall Session | Pre-Set C | Post-Set C | Δset C | Pre-Set B | Post-Set B | Δset B | Overall Session | ES | ||||
| pH | 7.40 ± 0.04 | 7.34 ± 0.05 | −0.06 ± 0.07 | 7.36 ± 0.04 | 7.24 ± 0.05 | −0.12 ± 0.06 | 7.34 ± 0.07 | 7.41 ± 0.02 | 7.22 ± 0.04 | −0.19 ± 0.04 * | 7.26 ± 0.04 | 7.32 ± 0.04 | 0.06 ± 0.06 * | 7.30 ± 0.08 # | 0.43 | |||
| BE (mmol·L−1) | 0.3 ± 1.2 | −7.2 ± 4.0 | −7.5 ± 3.9 | −5.1 ± 3.0 | −14.3 ± 2.8 | −9.3 ± 3.7 | −6.6 ± 6.0 | 0.5 ± 1.9 | −14.9 ± 2.0 | −15.4 ± 2.6 * | −12.9 ± 2.5 | −9.4 ± 2.7 | 3.5 ± 3.4 * | −9.2 ± 6.4 # | 0.42 | |||
| HCO3 (mmol·L−1) | 25.1 ± 0.8 | 18.6 ± 3.3 | −6.5 ± 3.2 | 20.3 ± 2.5 | 13.1 ± 2.1 | −7.2 ± 2.9 | 19.3 ± 4.9 | 25.1 ± 1.7 | 12.8 ± 1.4 | −12.3 ± 2.1 * | 14.2 ± 2.0 | 16.7 ± 2.1 | 2.5 ± 2.6 * | 17.2 ± 5.1 # | 0.41 | |||
| Ln RMSSD (ms) | 2.9 ± 0.3 | 2.1 ± 0.4 | - | - | 1.9 ± 0.3 | - | 2.3 ± 0.5 | 2.7 ± 0.2 | 1.7 ± 0.4 | - | - | 1.8 ± 0.4 * | - | 2.0 ± 0.6 | 0.50 | |||
| RPE (a.u.) | 2.4 ± 2.2 | 7.5 ± 2.2 | 5.0 ± 2.7 | 4.6 ± 1.6 | 9.4 ± 0.8 | 4.8 ± 1.9 | 7.5 ± 2.0 | 3.1 ± 2.4 * | 8.9 ± 1.4 | 5.8 ± 2.6 | 5.1 ± 1.9 * | 8.3 ± 1.6 | 3.3 ± 1.4 | 7.9 ± 1.1 | 0.30 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikitakis, I.S.; Bogdanis, G.C.; Paradisis, G.P.; Toubekis, A.G. Effects of Training Sets Sequence on Swimming Performance, Training Load and Physiological Responses. Sports 2023, 11, 240. https://doi.org/10.3390/sports11120240
Nikitakis IS, Bogdanis GC, Paradisis GP, Toubekis AG. Effects of Training Sets Sequence on Swimming Performance, Training Load and Physiological Responses. Sports. 2023; 11(12):240. https://doi.org/10.3390/sports11120240
Chicago/Turabian StyleNikitakis, Ioannis S., Gregory C. Bogdanis, Giorgos P. Paradisis, and Argyris G. Toubekis. 2023. "Effects of Training Sets Sequence on Swimming Performance, Training Load and Physiological Responses" Sports 11, no. 12: 240. https://doi.org/10.3390/sports11120240