
Sex-Specific miRNA Differences in Liquid Biopsies from Subjects with Solid Tumors and Healthy Controls
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
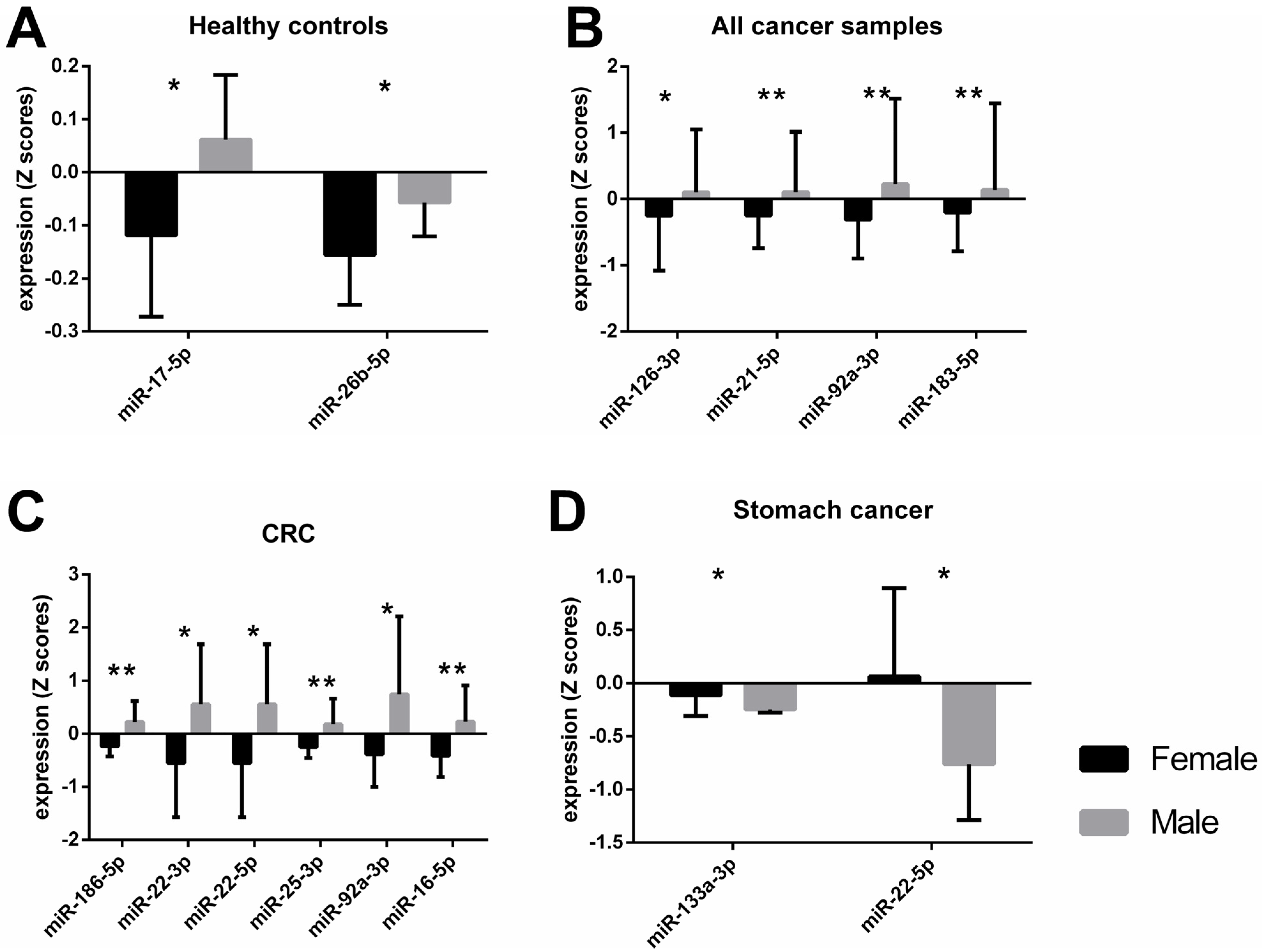
2.1. Differences in miRNA Expression between Female and Male Participants
2.2. DNA Methylation Levels Results
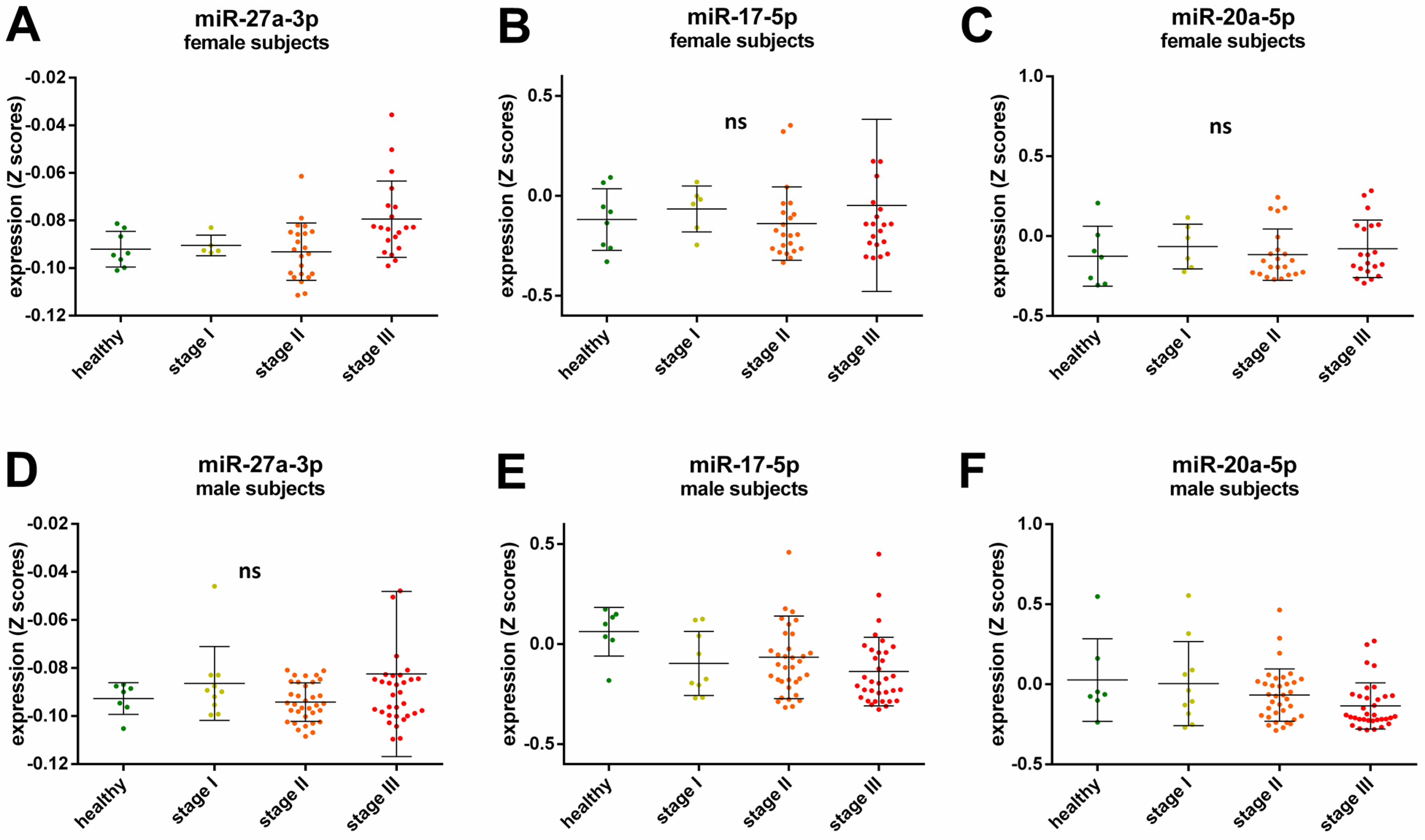
2.3. Sex-Specific miRNA Expression Related to Cancer Stage
3. Discussion
4. Materials and Methods
4.1. Study Population and Sample Processing
4.2. Cell-Free DNA Extraction and Methylation Analysis
4.3. Circulating miRNA Extraction and Analysis
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Migliore, L.; Nicolì, V.; Stoccoro, A. Gender Specific Differences in Disease Susceptibility: The Role of Epigenetics. Biomedicines 2021, 9, 652. [Google Scholar] [CrossRef] [PubMed]
- Vallabhajosyula, S.; Verghese, D.; Desai, V.K.; Sundaragiri, P.R.; Miller, V.M. Sex Differences in Acute Cardiovascular Care: A Review and Needs Assessment. Cardiovasc. Res. 2022, 118, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Walli-Attaei, M.; Joseph, P.; Rosengren, A.; Chow, C.K.; Rangarajan, S.; Lear, S.A.; AlHabib, K.F.; Davletov, K.; Dans, A.; Lanas, F.; et al. Variations between Women and Men in Risk Factors, Treatments, Cardiovascular Disease Incidence, and Death in 27 High-Income, Middle-Income, and Low-Income Countries (PURE): A Prospective Cohort Study. Lancet 2020, 396, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Morgan, C.P.; Bale, T.L. Sex Differences in MicroRNA Regulation of Gene Expression: No Smoke, Just MiRs. Biol. Sex Differ. 2012, 3, 22. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Eghbali, M. Influence of Sex Differences on MicroRNA Gene Regulation in Disease. Biol. Sex Differ. 2014, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Liu, Y.; Goldin, L.R.; Lyu, C.; Kong, X.; Zhang, Y.; Caporaso, N.E.; Xiang, S.; Gao, Y. Sex-Related DNA Methylation Differences in B Cell Chronic Lymphocytic Leukemia. Biol. Sex Differ. 2019, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kelly, T.K.; Jones, P.A. Epigenetics in Cancer. Carcinogenesis 2010, 31, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, S.; Van Emburgh, B.O.; Robertson, K.D. DNA Methylation in Development and Human Disease. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2008, 647, 30–38. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Suriya Muthukumaran, N.; Velusamy, P.; Akino Mercy, C.S.; Langford, D.; Natarajaseenivasan, K.; Shanmughapriya, S. MicroRNAs as Regulators of Cancer Cell Energy Metabolism. JPM 2022, 12, 1329. [Google Scholar] [CrossRef]
- Peng, Y.; Croce, C.M. The Role of MicroRNAs in Human Cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [Green Version]
- Olive, V.; Sabio, E.; Bennett, M.J.; De Jong, C.S.; Biton, A.; McGann, J.C.; Greaney, S.K.; Sodir, N.M.; Zhou, A.Y.; Balakrishnan, A.; et al. A Component of the Mir-17-92 Polycistronic Oncomir Promotes Oncogene-Dependent Apoptosis. eLife 2013, 2, e00822. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Zhang, X.; Sun, S.; Wu, D. MiR-92a Contributes to Cell Proliferation, Apoptosis and Doxorubicin Chemosensitivity in Gastric Carcinoma Cells. Oncol. Rep. 2019, 42, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Vecchiarelli-Federico, L.M.; Li, Y.-J.; Egan, S.E.; Spaner, D.; Hough, M.R.; Ben-David, Y. The MiR-17-92 Cluster Expands Multipotent Hematopoietic Progenitors Whereas Imbalanced Expression of Its Individual Oncogenic MiRNAs Promotes Leukemia in Mice. Blood 2012, 119, 4486–4498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Huang, S.; Ding, J.; Zhao, Y.; Liang, L.; Liu, T.; Zhan, R.; He, X. Multiple MicroRNAs Modulate P21Cip1/Waf1 Expression by Directly Targeting Its 3′ Untranslated Region. Oncogene 2010, 29, 2302–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.-X.; Yin, S.; Ma, L.; Wheeler, A.; Chen, Y.; Zhang, Y.; Liu, B.; Xiong, J.; Zhang, W.; Hu, J.; et al. 5-Hydroxymethylcytosine Signatures in Cell-Free DNA Provide Information about Tumor Types and Stages. Cell Res. 2017, 27, 1231–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Casanova, A.; Costa-Fraga, N.; Bao-Caamano, A.; López-López, R.; Muinelo-Romay, L.; Diaz-Lagares, A. Epigenetic Landscape of Liquid Biopsy in Colorectal Cancer. Front. Cell Dev. Biol. 2021, 9, 622459. [Google Scholar] [CrossRef]
- Russano, M.; Napolitano, A.; Ribelli, G.; Iuliani, M.; Simonetti, S.; Citarella, F.; Pantano, F.; Dell’Aquila, E.; Anesi, C.; Silvestris, N.; et al. Liquid Biopsy and Tumor Heterogeneity in Metastatic Solid Tumors: The Potentiality of Blood Samples. J. Exp. Clin. Cancer Res. 2020, 39, 95. [Google Scholar] [CrossRef]
- Lu, Y.; Chan, Y.-T.; Tan, H.-Y.; Li, S.; Wang, N.; Feng, Y. Epigenetic Regulation in Human Cancer: The Potential Role of Epi-Drug in Cancer Therapy. Mol. Cancer 2020, 19, 79. [Google Scholar] [CrossRef]
- Rubin, J.B.; Lagas, J.S.; Broestl, L.; Sponagel, J.; Rockwell, N.; Rhee, G.; Rosen, S.F.; Chen, S.; Klein, R.S.; Imoukhuede, P.; et al. Sex Differences in Cancer Mechanisms. Biol. Sex Differ. 2020, 11, 17. [Google Scholar] [CrossRef]
- Tomeva, E.; Switzeny, O.J.; Heitzinger, C.; Hippe, B.; Haslberger, A.G. Comprehensive Approach to Distinguish Patients with Solid Tumors from Healthy Controls by Combining Androgen Receptor Mutation p.H875Y with Cell-Free DNA Methylation and Circulating MiRNAs. Cancers 2022, 14, 462. [Google Scholar] [CrossRef]
- Zhang, X.; Li, Z.; Xuan, Z.; Xu, P.; Wang, W.; Chen, Z.; Wang, S.; Sun, G.; Xu, J.; Xu, Z. Novel Role of MiR-133a-3p in Repressing Gastric Cancer Growth and Metastasis via Blocking Autophagy-Mediated Glutaminolysis. J. Exp. Clin. Cancer Res. 2018, 37, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaude, E.; Frezza, C. Tissue-Specific and Convergent Metabolic Transformation of Cancer Correlates with Metastatic Potential and Patient Survival. Nat. Commun. 2016, 7, 13041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Sun, Q.; Wang, X. Transcriptional Landscape of Human Cancers. Oncotarget 2017, 8, 34534–34551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Ning, S.; Ghandi, M.; Kryukov, G.V.; Gopal, S.; Deik, A.; Souza, A.; Pierce, K.; Keskula, P.; Hernandez, D.; et al. The Landscape of Cancer Cell Line Metabolism. Nat. Med. 2019, 25, 850–860. [Google Scholar] [CrossRef]
- Wang, W.; Li, F.; Zhang, Y.; Tu, Y.; Yang, Q.; Gao, X. Reduced Expression of MiR-22 in Gastric Cancer Is Related to Clinicopathologic Characteristics or Patient Prognosis. Diagn. Pathol. 2013, 8, 102. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Jiang, Y.; Zhang, H.; Greenlee, A.R.; Yu, R.; Yang, Q. MiR-22 Functions as a Micro-Oncogene in Transformed Human Bronchial Epithelial Cells Induced by Anti-Benzo[a]Pyrene-7,8-Diol-9,10-Epoxide. Toxicol. Vitr. 2010, 24, 1168–1175. [Google Scholar] [CrossRef]
- Poliseno, L.; Salmena, L.; Riccardi, L.; Fornari, A.; Song, M.S.; Hobbs, R.M.; Sportoletti, P.; Varmeh, S.; Egia, A.; Fedele, G.; et al. Identification of the MiR-106b ~ 25 MicroRNA Cluster as a Proto-Oncogenic PTEN -Targeting Intron That Cooperates with Its Host Gene MCM7 in Transformation. Sci. Signal. 2010, 3, ra29. [Google Scholar] [CrossRef] [Green Version]
- Tong, D.; Zhang, J.; Wang, X.; Li, Q.; Liu, L.; Lu, A.; Guo, B.; Yang, J.; Ni, L.; Qin, H.; et al. MiR-22, Regulated by MeCP2, Suppresses Gastric Cancer Cell Proliferation by Inducing a Deficiency in Endogenous S-Adenosylmethionine. Oncogenesis 2020, 9, 99. [Google Scholar] [CrossRef]
- Sadre-Marandi, F.; Dahdoul, T.; Reed, M.C.; Nijhout, H.F. Sex Differences in Hepatic One-Carbon Metabolism. BMC Syst. Biol. 2018, 12, 89. [Google Scholar] [CrossRef]
- Bhat-Nakshatri, P.; Wang, G.; Collins, N.R.; Thomson, M.J.; Geistlinger, T.R.; Carroll, J.S.; Brown, M.; Hammond, S.; Srour, E.F.; Liu, Y.; et al. Estradiol-Regulated MicroRNAs Control Estradiol Response in Breast Cancer Cells. Nucleic Acids Res. 2009, 37, 4850–4861. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, S.; Hvid, T.; Kelly, M.; Lindegaard, B.; Dethlefsen, C.; Winding, K.; Mathur, N.; Scheele, C.; Pedersen, B.K.; Laye, M.J. Muscle Specific MiRNAs Are Induced by Testosterone and Independently Upregulated by Age. Front. Physiol. 2014, 4, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, Y.; Tian, Q.; Guan, L.; Niu, S. The Dual Role of MiR-186 in Cancers: Oncomir Battling with Tumor Suppressor MiRNA. Front. Oncol. 2020, 10, 233. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Xia, L.; Zhou, Z.; Zuo, Z.; Xu, C.; Song, H.; Cai, J. MiR-186-5p Upregulation Inhibits Proliferation, Metastasis and Epithelial-to-Mesenchymal Transition of Colorectal Cancer Cell by Targeting ZEB1. Arch. Biochem. Biophys. 2018, 640, 53–60. [Google Scholar] [CrossRef]
- Islam, F.; Gopalan, V.; Vider, J.; Wahab, R.; Ebrahimi, F.; Lu, C.; Kasem, K.; Lam, A.K.Y. MicroRNA-186-5p Overexpression Modulates Colon Cancer Growth by Repressing the Expression of the FAM134B Tumour Inhibitor. Exp. Cell Res. 2017, 357, 260–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Wang, X.; Guo, Q.; Zhu, S.; Li, P.; Zhang, S.; Min, L. M2 Macrophage Derived Extracellular Vesicle-Mediated Transfer of MiR-186-5p Promotes Colon Cancer Progression by Targeting DLC1. Int. J. Biol. Sci. 2022, 18, 1663–1676. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.-L.; Lu, L.-L.; Dong, L.-L.; Liu, Y.; Bian, X.-Y.; Lian, B.-F.; Xie, L.; Wen, D.; Gao, D.-M.; Ke, A.-W.; et al. MiR-17-5p and MiR-20a-5p Suppress Postoperative Metastasis of Hepatocellular Carcinoma via Blocking HGF/ERBB3-NF-ΚB Positive Feedback Loop. Theranostics 2020, 10, 3668–3683. [Google Scholar] [CrossRef]
- Su, C.; Huang, D.; Liu, J.; Liu, W.; Cao, Y. MiR-27a-3p Regulates Proliferation and Apoptosis of Colon Cancer Cells by Potentially Targeting BTG1. Oncol. Lett. 2019, 18, 2825–2834. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Zhang, Q.; Ma, X.; Wang, J.; Liang, T. MiRNA and MRNA Expression Analysis Reveals Potential Sex-Biased MiRNA Expression. Sci. Rep. 2017, 7, 39812. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, O.; Pers, Y.-M.; Ferreira, R.; Sénéchal, A.; Jorgensen, C.; Apparailly, F.; Duroux-Richard, I. X-Linked MiRNAs Associated with Gender Differences in Rheumatoid Arthritis. Int. J. Mol. Sci. 2016, 17, 1852. [Google Scholar] [CrossRef]
- Li, Z.; Liu, J.-Y.; Zhang, J.-T. 14-3-3sigma, the Double-Edged Sword of Human Cancers. Am. J. Transl. Res. 2009, 1, 326–340. [Google Scholar] [PubMed]
- Singmann, P.; Shem-Tov, D.; Wahl, S.; Grallert, H.; Fiorito, G.; Shin, S.-Y.; Schramm, K.; Wolf, P.; Kunze, S.; Baran, Y.; et al. Characterization of Whole-Genome Autosomal Differences of DNA Methylation between Men and Women. Epigenetics Chromatin 2015, 8, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mestdagh, P.; Van Vlierberghe, P.; De Weer, A.; Muth, D.; Westermann, F.; Speleman, F.; Vandesompele, J. A Novel and Universal Method for MicroRNA RT-QPCR Data Normalization. Genome Biol. 2009, 10, R64. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Groups | n (f/m) | miRNA | Mean Difference Female–Male (SED) | t-Test p-Value | Hedges’ g Effect Size | Adjusted p-Value | Partial Eta Squared η2 |
|---|---|---|---|---|---|---|---|
| Healthy c | 15 (8/7) | miR-17-5p | −0.180 (0.072) | 0.027 | 1.291 | 0.027 * | 0.371 |
| miR-26b-5p | −0.098 (0.042) | 0.036 | 1.208 | 0.011 * | 0.46 | ||
| All cancer a,c | 128 (48/80) | miR-126-3p | −0.352 (0.165) | 0.035 | 0.389 | 0.017 * | 0.046 |
| miR-21-5p | −0.353 (0.124) | 0.005 | 0.452 | 0.006 ** | 0.059 | ||
| miR-92a-3p | −0.537 (0.167) | 0.002 | 0.497 | 0.01 * | 0.053 | ||
| miR-183-5p | −0.346 (0.168) | 0.041 | 0.318 | 0.015 * | 0.046 | ||
| miR-16-5p | −0.269 (0.111) | 0.017 | 0.392 | 0.075 | 0.025 | ||
| CRC b,c | 28 (14/14) | miR-133a-3p | −0.558 (0.249) | 0.043 | 0.847 | 0.076 | 0.116 |
| miR-186-5p | −0.463 (0.117) | 0.001 | 1.503 | 0.003 ** | 0.287 | ||
| miR-195-5p | −0.381 (0.126) | 0.007 | 1.142 | 0.085 | 0.11 | ||
| miR-21-5p | −0.562 (0.258) | 0.039 | 0.721 | 0.115 | 0.104 | ||
| miR-210-3p | −0.741 (0.293) | 0.018 | 0.956 | 0.061 | 0.145 | ||
| miR-22-3p | −0.984 (0.421) | 0.027 | 0.884 | 0.039 * | 0.173 | ||
| miR-22-5p | −1.105 (0.407) | 0.012 | 1.025 | 0.038 * | 0.174 | ||
| miR-25-3p | −0.431 (0.141) | 0.007 | 1.159 | 0.003 ** | 0.324 | ||
| miR-34a-5p | −0.496 (0.199) | 0.026 | 0.941 | 0.411 | 0.026 | ||
| miR-92a-3p | −1.132 (0.423) | 0.013 | 1.013 | 0.007 ** | 0.278 | ||
| miR-16-5p | −0.644 (0.211) | 0.005 | 1.155 | 0.021 * | 0.21 | ||
| Stomach cancer b,c | 23 (15/8) | miR-133a-3p | 0.136 (0.052) | 0.019 | 0.842 | 0.037 * | 0.19 |
| miR-22-5p | 0.823 (0.326) | 0.02 | 1.106 | 0.042 * | 0.21 |
| Subjects | n | Age Years Mean (SD) | BMI kg/m2 Mean (SD) | Cancer Stage n | ||
|---|---|---|---|---|---|---|
| I | II | III | ||||
| Female | 56 | 64.0 (14.5) | 24.1 (3.4) b | 6 | 22 | 20 |
| Healthy | 8 | 49.4 (16.7) | 25.6 (4.2) | 0 | 0 | 0 |
| Bladder | 3 | 81.0 (10.8) | 23.9 (2.7) | 3 | 0 | 0 |
| Brain | 3 | 61.3 (17.5) | 25.9 (1.5) | 1 | 2 | 0 |
| CRC | 14 | 69.1 (11.1) | 23.7 (2.9) | 1 | 11 | 2 |
| Lung | 4 | 54.5 (3.0) | 24.5 (2.4) | 0 | 1 | 3 |
| Stomach | 15 | 70.9 (11.2) | 23.0 (4.0) | 0 | 5 | 10 |
| Pancreas | 6 | 62.3 (4.5) a | 25.4 (2.1) | 0 | 2 | 4 |
| Liver | 3 | 47.3 (13.4) | 22.7 (5.8) | 1 | 1 | 1 |
| Male | 87 | 65.3 (9.1) | 25.5 (3.5) b | 10 | 35 | 35 |
| Healthy | 7 | 60.6 (11.5) | 25.3 (1.1) | 0 | 0 | 0 |
| Bladder | 17 | 68.9 (9.1) | 26.7 (2.9) | 6 | 6 | 5 |
| Brain | 6 | 64.2 (9.4) | 24.9 (4.1) | 1 | 2 | 3 |
| CRC | 14 | 64.1 (7.7) | 25.6 (2.8) | 2 | 8 | 4 |
| Lung | 25 | 63.9 (9.4) | 29.9 (4.3) | 1 | 11 | 13 |
| Stomach | 8 | 66.5 (6.8) | 22.9 (4.9) | 0 | 3 | 5 |
| Pancreas | 6 | 71.8 (8.9) a | 23.4 (2.5) | 0 | 3 | 3 |
| Liver | 4 | 61.3 (5.7) | 26.3 (0.4) | 0 | 2 | 2 |
| Total | 143 | 64.8 (11.5) | 24.9 (3.6) | 16 | 57 | 55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomeva, E.; Krammer, U.D.B.; Switzeny, O.J.; Haslberger, A.G.; Hippe, B. Sex-Specific miRNA Differences in Liquid Biopsies from Subjects with Solid Tumors and Healthy Controls. Epigenomes 2023, 7, 2. https://doi.org/10.3390/epigenomes7010002
Tomeva E, Krammer UDB, Switzeny OJ, Haslberger AG, Hippe B. Sex-Specific miRNA Differences in Liquid Biopsies from Subjects with Solid Tumors and Healthy Controls. Epigenomes. 2023; 7(1):2. https://doi.org/10.3390/epigenomes7010002
Chicago/Turabian StyleTomeva, Elena, Ulrike D. B. Krammer, Olivier J. Switzeny, Alexander G. Haslberger, and Berit Hippe. 2023. "Sex-Specific miRNA Differences in Liquid Biopsies from Subjects with Solid Tumors and Healthy Controls" Epigenomes 7, no. 1: 2. https://doi.org/10.3390/epigenomes7010002