
Polycomb Repressive Complex 2 in Eukaryotes—An Evolutionary Perspective


Abstract
:1. Introduction
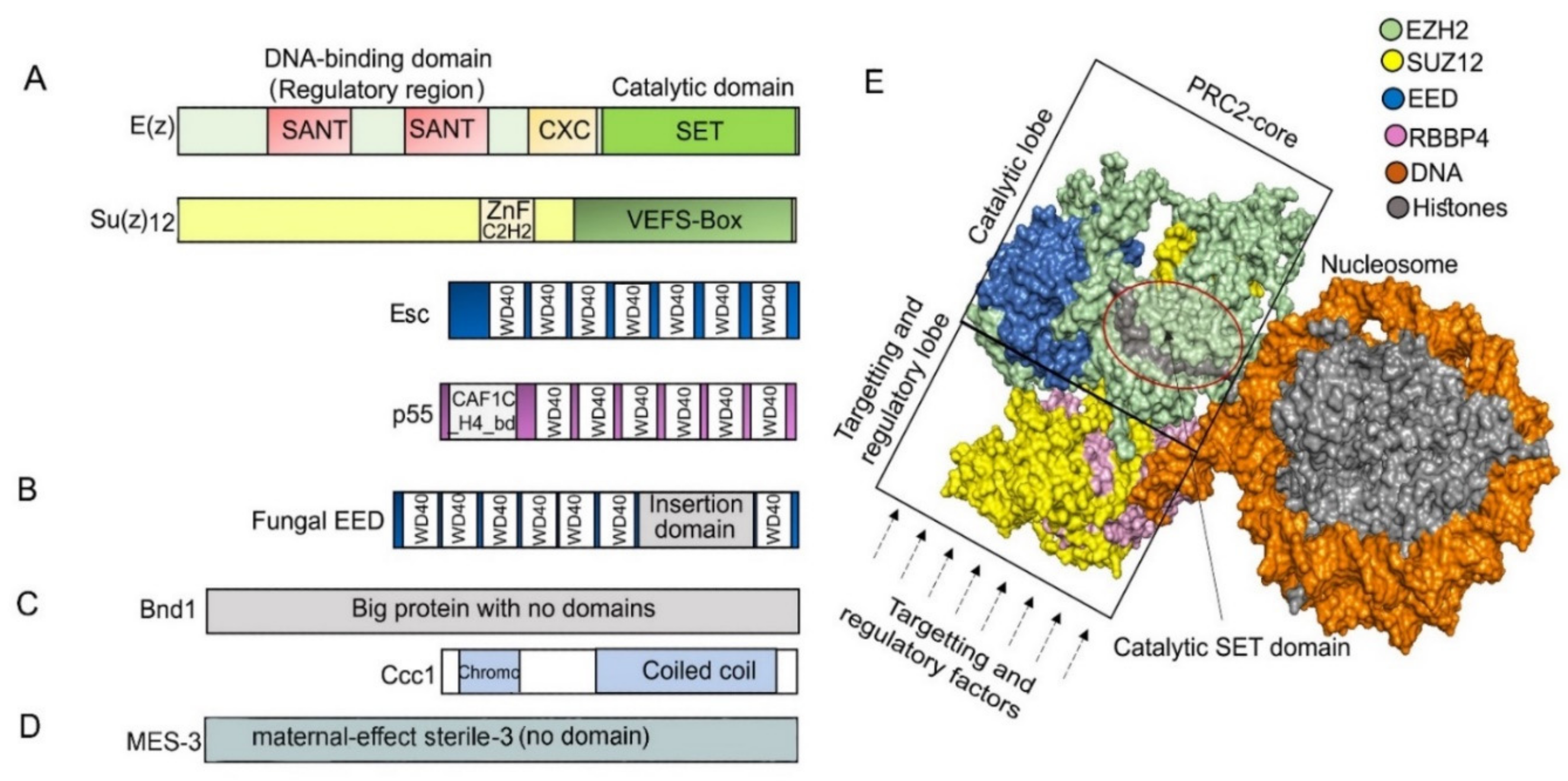
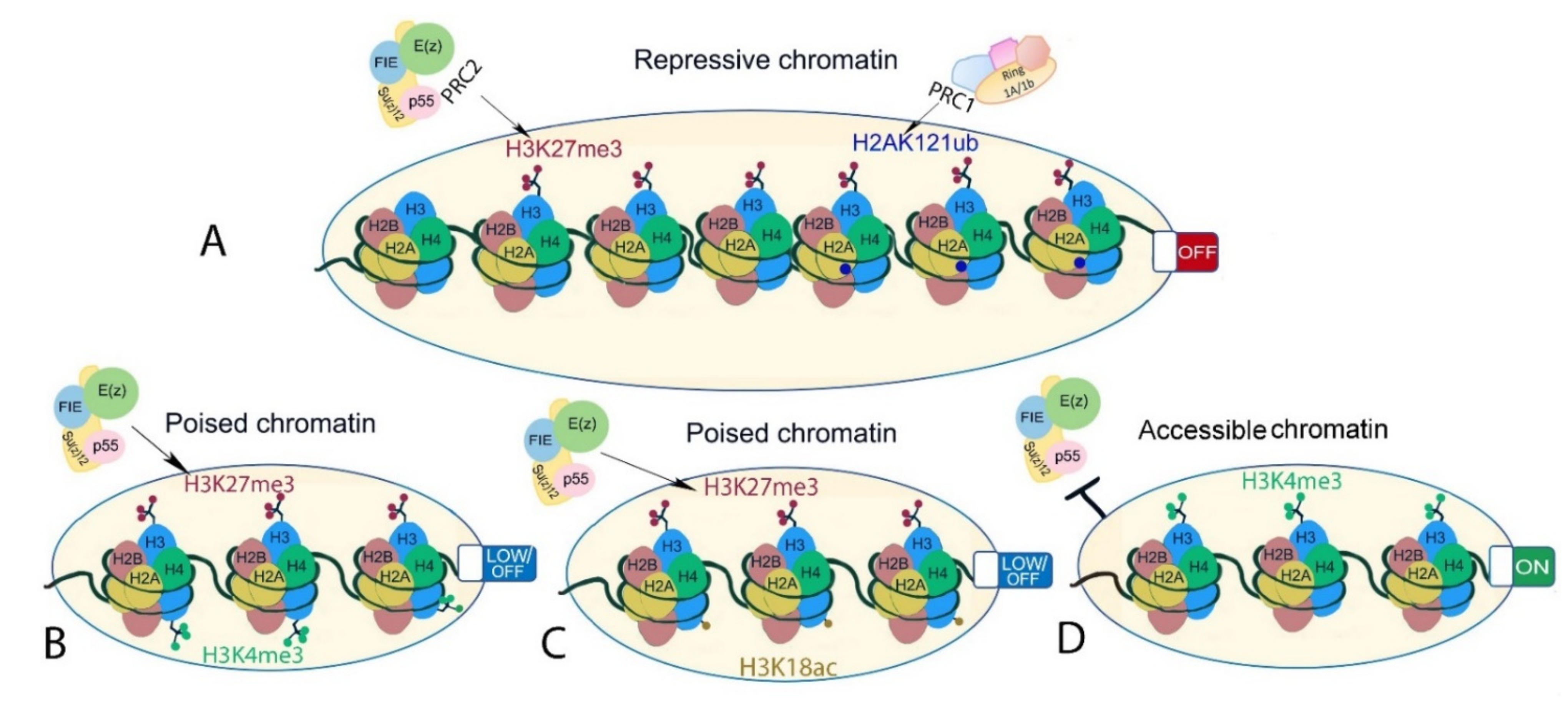
2. Features of PRC2 Core Composition and Function Are Conserved in Animal and Plant Models
3. PRC2 Is Conserved throughout Evolution in Unicellular and Multicellular Eukaryotes
3.1. PRC2 in Stramenopiles, Alveolates and Rhizaria (SAR)
3.2. PRC2 in Ophithokonts
3.3. PRC2 in Cryptophytes, Red Algae, and the Green Lineage
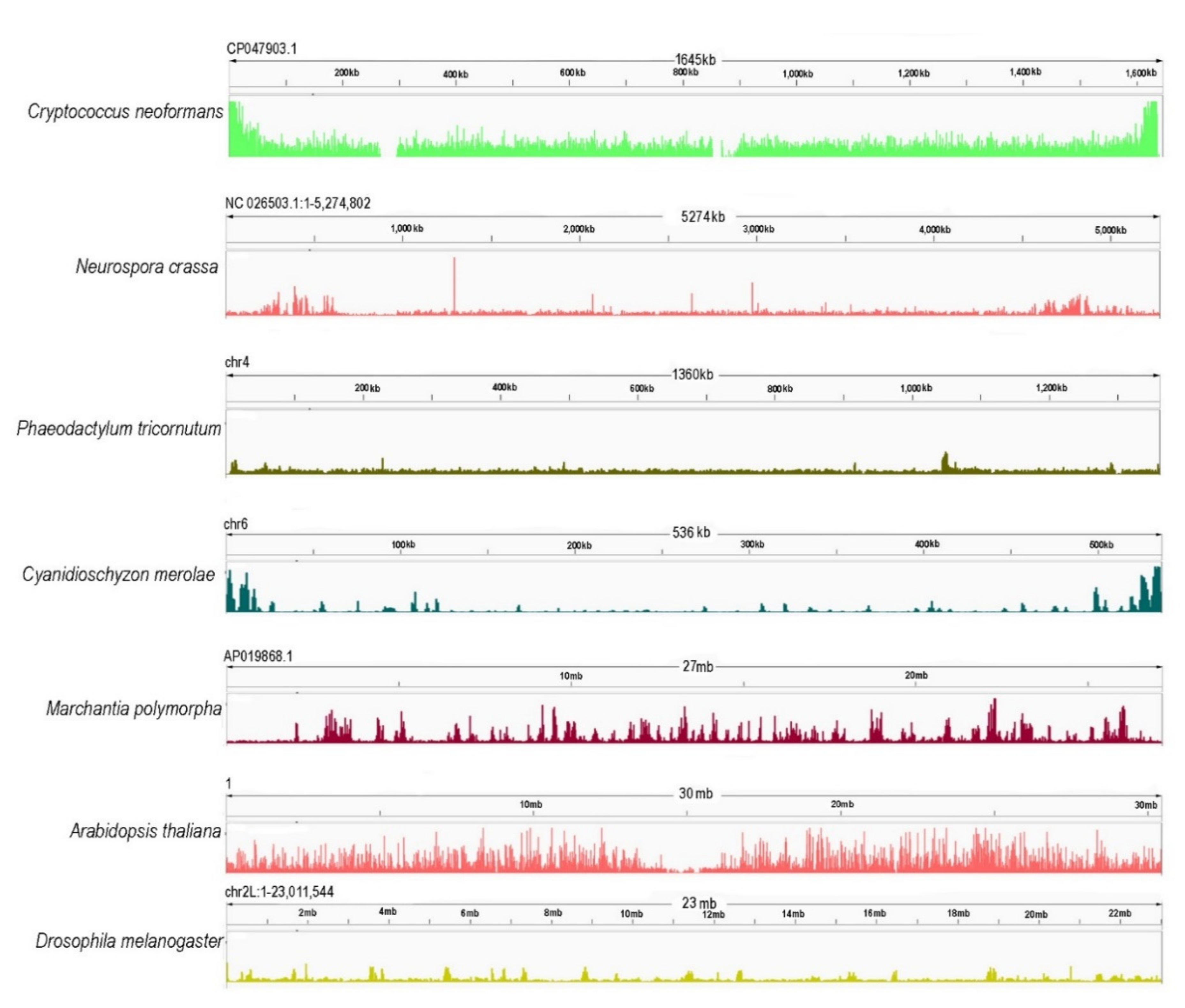
4. PRC2 Targeting to Repeats and Regions of Constitutive Heterochromatin
5. Emerging Patterns and Questions in PRC2 Evolution
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prezioso, C.; Orlando, V. Polycomb Proteins in Mammalian Cell Differentiation and Plasticity. FEBS Lett. 2011, 585, 2067–2077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aranda, S.; Mas, G.; Di Croce, L. Regulation of Gene Transcription by Polycomb Proteins. Sci. Adv. 2015, 1, e1500737. [Google Scholar] [CrossRef] [Green Version]
- Mozgova, I.; Hennig, L. The Polycomb Group Protein Regulatory Network. Annu. Rev. Plant Biol. 2015. [Google Scholar] [CrossRef]
- Yu, J.R.; Lee, C.H.; Oksuz, O.; Stafford, J.M.; Reinberg, D. PRC2 Is High Maintenance. Genes Dev. 2019, 33, 903–935. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Lin, Y.; Li, Y.; Wang, G. Plants Dynamics of H3K27me3 Modification on Plant Adaptation to Environmental Cues. Plants 2021, 10, 1165. [Google Scholar] [CrossRef]
- Lee, H.-G.; Kahn, T.G.; Simcox, A.; Schwartz, Y.B.; Pirrotta, V. Genome-Wide Activities of Polycomb Complexes Control Pervasive Transcription. Genome Res. 2015, 25, 1170–1181. [Google Scholar] [CrossRef] [Green Version]
- Bratzel, F.; López-Torrejón, G.; Koch, M.; Del Pozo, J.C.; Calonje, M. Keeping Cell Identity in Arabidopsis Requires PRC1 RING-Finger Homologs That Catalyze H2A Monoubiquitination. Curr. Biol. 2010, 20, 1853–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, L.; Erdjument-Bromage, H.; Vidal, M.; Tempst, P.; Jones, R.S.; Zhang, Y. Role of Histone H2A Ubiquitination in Polycomb Silencing. Nature 2004, 431, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Wiles, E.T.; Selker, E.U. H3K27 Methylation: A Promiscuous Repressive Chromatin Mark. Curr. Opin. Genet. Dev. 2017, 43, 31–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chittock, E.C.; Latwiel, S.; Miller, T.C.R.; Müller, C.W. Molecular Architecture of Polycomb Repressive Complexes. Biochem. Soc. Trans. 2017, 45, 193–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddington, C.J.; Fellner, M.; Burgess, A.E.; Mace, P.D. Molecular Regulation of the Polycomb Repressive-Deubiquitinase. Int. J. Mol. Sci. 2020, 21, 7837. [Google Scholar] [CrossRef] [PubMed]
- Derkacheva, M.; Liu, S.; Figueiredo, D.D.; Gentry, M.; Mozgova, I.; Nanni, P.; Tang, M.; Mannervik, M.; Köhler, C.; Hennig, L. H2A Deubiquitinases UBP12/13 Are Part of the Arabidopsis Polycomb Group Protein System. Nat. Plants 2016, 2, 16126. [Google Scholar] [CrossRef]
- Kralemann, L.E.M.; Liu, S.; Trejo-Arellano, M.S.; Muñoz-Viana, R.; Köhler, C.; Hennig, L. Removal of H2Aub1 by Ubiquitin-Specific Proteases 12 and 13 Is Required for Stable Polycomb-Mediated Gene Repression in Arabidopsis. Genome Biol. 2020, 21, 144. [Google Scholar] [CrossRef]
- Klymenko, T.; Papp, B.; Fischle, W.; Köcher, T.; Schelder, M.; Fritsch, C.; Wild, B.; Wilm, M.; Müller, J. A Polycomb Group Protein Complex with Sequence-Specific DNA-Binding and Selective Methyl-Lysine-Binding Activities. Genes Dev. 2006, 20, 1110–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuettengruber, B.; Bourbon, H.M.; Di Croce, L.; Cavalli, G. Genome Regulation by Polycomb and Trithorax: 70 Years and Counting. Cell 2017, 171, 34–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart-Morgan, K.R.; Petryk, N.; Groth, A. Chromatin Replication and Epigenetic Cell Memory. Nat. Cell Biol. 2020, 22, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Probst, A.V.; Desvoyes, B.; Gutierrez, C. Similar yet Critically Different: The Distribution, Dynamics and Function of Histone Variants. J. Exp. Bot. 2020, 71, 5191–5204. [Google Scholar] [CrossRef] [PubMed]
- Hugues, A.; Jacobs, C.S.; Roudier, F. Mitotic Inheritance of PRC2-Mediated Silencing: Mechanistic Insights and Developmental Perspectives. Front. Plant Sci. 2020, 11, 262. [Google Scholar] [CrossRef] [Green Version]
- Pu, L.; Sung, Z.R. PcG and TrxG in Plants—Friends or Foes. Trends Genet. 2015, 31, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Ordonez-Rubiano, S.C.; Dhiman, A.; Jiao, G.; Strohmier, B.P.; Krusemark, C.J.; Dykhuizen, E.C. Polycomb Group Proteins in Cancer: Multifaceted Functions and Strategies for Modulation. NAR Cancer 2021, 3, zcab039. [Google Scholar] [CrossRef] [PubMed]
- Dockerill, M.; Gregson, C.; O’ Donovan, D.H. Targeting PRC2 for the Treatment of Cancer: An Updated Patent Review (2016–2020). Expert Opin. Ther. Pat. 2021, 31, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Piunti, A.; Shilatifard, A. The Roles of Polycomb Repressive Complexes in Mammalian Development and Cancer. Nat. Rev. Mol. Cell Biol. 2021, 22, 326–345. [Google Scholar] [CrossRef]
- Lewis, P.H. New Mutants Report. Drosoph. Inf. Serv. 1947, 21, 69. [Google Scholar]
- Sanchez-Pulido, L.; Devos, D.; Sung, Z.R.; Calonje, M. RAWUL: A New Ubiquitin-like Domain in PRC1 Ring Finger Proteins That Unveils Putative Plant and Worm PRC1 Orthologs. BMC Genom. 2008, 9, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Molitor, A.; Liu, C.; Shen, W.H. The Arabidopsis PRC1-like Ring-Finger Proteins Are Necessary for Repression of Embryonic Traits during Vegetative Growth. Cell Res. 2010, 20, 1332–1344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Romero-Campero, F.J.; Gómez-Zambrano, Á.; Turck, F.; Calonje, M. H2A Monoubiquitination in Arabidopsis Thaliana Is Generally Independent of LHP1 and PRC2 Activity. Genome Biol. 2017, 18, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berke, L.; Snel, B. The Plant Polycomb Repressive Complex 1 (PRC1) Existed in the Ancestor of Seed Plants and Has a Complex Duplication History. BMC Evol. Biol. 2015, 15, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.; Huang, Y.; Ruan, Y.; Shen, W.-H. The Evolutionary Landscape of PRC1 Core Components in Green Lineage. Planta 2016, 243, 825–846. [Google Scholar] [CrossRef] [PubMed]
- Hennig, L.; Derkacheva, M. Diversity of Polycomb Group Complexes in Plants: Same Rules, Different Players? Trends Genet. 2009, 25, 414–423. [Google Scholar] [CrossRef]
- Calonje, M. Prc1 Marks the Difference in Plant Pcg Repression. Mol. Plant 2014, 7, 459–471. [Google Scholar] [CrossRef] [Green Version]
- Shaver, S.; Casas-Mollano, J.A.; Cerny, R.L.; Cerutti, H. Origin of the Polycomb Repressive Complex 2 and Gene Silencing by an E(z) Homolog in the Unicellular Alga Chlamydomonas. Epigenetics 2010, 5, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Sowpati, D.T.; Ramamoorthy, S.; Mishra, R.K. Expansion of the Polycomb System and Evolution of Complexity. Mech. Dev. 2015, 138, 97–112. [Google Scholar] [CrossRef]
- Förderer, A.; Zhou, Y.; Turck, F. The Age of Multiplexity: Recruitment and Interactions of Polycomb Complexes in Plants. Curr. Opin. Plant Biol. 2016, 29, 169–178. [Google Scholar] [CrossRef] [PubMed]
- van Mierlo, G.; Veenstra, G.J.C.; Vermeulen, M.; Marks, H. The Complexity of PRC2 Subcomplexes. Trends Cell Biol. 2019, 29, 660–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, S.A.; Damle, M.; Kim, J.; Kingston, R.E. Full Methylation of H3K27 by PRC2 Is Dispensable for Initial Embryoid Body Formation but Required to Maintain Differentiated Cell Identity. Development 2021, 148, dev196329. [Google Scholar] [CrossRef] [PubMed]
- Mozgová, I.; Muñoz-Viana, R.; Hennig, L. PRC2 Represses Hormone-Induced Somatic Embryogenesis in Vegetative Tissue of Arabidopsis Thaliana. PLoS Genet. 2017, 13, e1006562. [Google Scholar] [CrossRef] [PubMed]
- Turck, F.; Roudier, F.; Farrona, S.; Martin-Magniette, M.L.; Guillaume, E.; Buisine, N.; Gagnot, S.; Martienssen, R.A.; Coupland, G.; Colot, V. Arabidopsis TFL2/LHP1 Specifically Associates with Genes Marked by Trimethylation of Histone H3 Lysine 27. PLoS Genet. 2007, 3, e86. [Google Scholar] [CrossRef]
- Zhang, X.; Clarenz, O.; Cokus, S.; Bernatavichute, Y.V.; Pellegrini, M.; Goodrich, J.; Jacobsen, S.E. Whole-Genome Analysis of Histone H3 Lysine 27 Trimethylation in Arabidopsis. PLoS Biol. 2007, 5, e129. [Google Scholar] [CrossRef] [Green Version]
- Margueron, R.; Trojer, P.; Reinberg, D. The Key to Development: Interpreting the Histone Code? Curr. Opin. Genet. Dev. 2005, 15, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Steffen, P.A.; Ringrose, L. What Are Memories Made of? How Polycomb and Trithorax Proteins Mediate Epigenetic Memory. Nat. Rev. Mol. Cell Biol. 2014, 15, 340–356. [Google Scholar] [CrossRef]
- Kang, S.J.; Chun, T. Structural Heterogeneity of the Mammalian Polycomb Repressor Complex in Immune Regulation. Exp. Mol. Med. 2020, 52, 1004–1015. [Google Scholar] [CrossRef]
- Martin, C.J.; Moorehead, R.A. Polycomb Repressor Complex 2 Function in Breast Cancer (Review). Int. J. Oncol. 2020, 57, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, L.; Fan, Z. The Role and Mechanisms of Polycomb Repressive Complex 2 on the Regulation of Osteogenic and Neurogenic Differentiation of Stem Cells. Cell Prolif. 2021, 54, e13032. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhao, S.; Wang, G.G. Polycomb Gene Silencing Mechanisms: PRC2 Chromatin Targeting, H3K27me3 “Readout”, and Phase Separation-Based Compaction. Trends Genet. 2021, 37, 547–565. [Google Scholar] [CrossRef] [PubMed]
- Bieluszewski, T.; Xiao, J.; Yang, Y.; Wagner, D. PRC2 Activity, Recruitment, and Silencing: A Comparative Perspective. Trends Plant Sci. 2021, 26, 1186–1198. [Google Scholar] [CrossRef] [PubMed]
- Glancy, E.; Ciferri, C.; Bracken, A.P. Structural Basis for PRC2 Engagement with Chromatin. Curr. Opin. Struct. Biol. 2021, 67, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Shu, J.; Chen, C.; Li, C.; Cui, Y. The Complexity of PRC2 Catalysts CLF and SWN in Plants. Biochem. Soc. Trans. 2020, 48, 2779–2789. [Google Scholar] [CrossRef] [PubMed]
- Hinsch, V.; Adkins, S.; Manuela, D.; Xu, M. Post-Embryonic Phase Transitions Mediated by Polycomb Repressive Complexes in Plants. Int. J. Mol. Sci. 2021, 22, 7533. [Google Scholar] [CrossRef] [PubMed]
- Déléris, A.; Berger, F.; Duharcourt, S. Role of Polycomb in the Control of Transposable Elements. Trends Genet. 2021, 37, 882–889. [Google Scholar] [CrossRef]
- Müller, J.; Hart, C.M.; Francis, N.J.; Vargas, M.L.; Sengupta, A.; Wild, B.; Miller, E.L.; O’Connor, M.B.; Kingston, R.E.; Simon, J.A. Histone Methyltransferase Activity of a Drosophila Polycomb Group Repressor Complex. Cell 2002, 111, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Li, G. Post-Translational Modifications of PRC2: Signals Directing Its Activity. Epigenet. Chromatin 2020, 13, 47. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, M.; Wild, B.; Müller, J. Nucleosome Binding and Histone Methyltransferase Activity of Drosophila PRC2. EMBO Rep. 2005, 6, 348–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Paucek, R.D.; Gooding, A.R.; Brown, Z.Z.; Eva, J.; Muir, T.W.; Cech, T.R. Molecular Analysis of PRC2 Recruitment to DNA in Chromatin and Its Inhibition by RNA. Nat. Struct. Mol. Biol. 2017, 24, 1028–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Goodrich, K.J.; Gooding, A.R.; Youmans, D.T.; Cech, T.R.; Wang, X.; Goodrich, K.J.; Gooding, A.R.; Naeem, H.; Archer, S.; et al. Targeting of Polycomb Repressive Complex 2 to RNA by Short Repeats of Consecutive Guanines Article Targeting of Polycomb Repressive Complex 2 to RNA by Short Repeats of Consecutive Guanines. Mol. Cell 2017, 65, 1056–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasinath, V.; Poepsel, S.; Nogales, E. Recent Structural Insights into PRC2 Regulation and Substrate Binding. Biochemistry 2019, 58, 346–354. [Google Scholar] [CrossRef]
- Holoch, D.; Margueron, R. Mechanisms Regulating PRC2 Recruitment and Enzymatic Activity. Trends Biochem. Sci. 2017, 42, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Ketel, C.S.; Andersen, E.F.; Vargas, M.L.; Suh, J.; Strome, S.; Simon, J.A. Subunit Contributions to Histone Methyltransferase Activities of Fly and Worm Polycomb Group Complexes. Mol. Cell. Biol. 2005, 25, 6857–6868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, R.; Zhang, Y. SUZ12 Is Required for Both the Histone Methyltransferase Activity and the Silencing Function of the EED-EZH2 Complex. Mol. Cell 2004, 15, 57–67. [Google Scholar] [CrossRef]
- Pasini, D.; Bracken, A.P.; Jensen, M.R.; Denchi, E.L.; Helin, K. Suz12 Is Essential for Mouse Development and for EZH2 Histone Methyltransferase Activity. EMBO J. 2004, 23, 4061–4071. [Google Scholar] [CrossRef] [Green Version]
- Kasinath, V.; Faini, M.; Poepsel, S.; Reif, D.; Feng, X.A.; Stjepanovic, G.; Aebersold, R.; Nogales, E. Structures of Human PRC2 with Its Cofactors AEBP2 and JARID2. Science 2018, 359, 940–944. [Google Scholar] [CrossRef] [Green Version]
- Mosquna, A.; Katz, A.; Decker, E.L.; Rensing, S.A.; Reski, R.; Ohad, N. Regulation of Stem Cell Maintenance by the Polycomb Protein FIE Has Been Conserved during Land Plant Evolution. Development 2009, 136, 2433–2444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okano, Y.; Aono, N.; Hiwatashi, Y.; Murata, T.; Nishiyama, T.; Ishikawa, T.; Kubo, M.; Hasebe, M. A Polycomb Repressive Complex 2 Gene Regulates Apogamy and Gives Evolutionary Insights into Early Land Plant Evolution. Proc. Natl. Acad. Sci. USA 2009, 106, 16321–16326. [Google Scholar] [CrossRef] [Green Version]
- Mikulski, P.; Komarynets, O.; Fachinelli, F.; Weber, A.P.M.; Schubert, D. Characterization of the Polycomb-Group Mark H3K27me3 in Unicellular Algae. Front. Plant Sci. 2017, 8, 607. [Google Scholar] [CrossRef] [PubMed]
- Dumesic, P.A.; Homer, C.M.; Moresco, J.J.; Pack, L.R.; Shanle, E.K.; Coyle, S.M.; Strahl, B.D.; Fujimori, D.G.; Yates, J.R.; Madhani, H.D. Product Binding Enforces the Genomic Specificity of a Yeast Polycomb Repressive Complex. Cell 2015, 160, 204–218. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Rastogi, A.; Deton Cabanillas, A.F.; Ait Mohamed, O.; Cantrel, C.; Lombard, B.; Murik, O.; Genovesio, A.; Bowler, C.; Bouyer, D.; et al. Genome Wide Natural Variation of H3K27me3 Selectively Marks Genes Predicted to Be Important for Cell Differentiation in Phaeodactylum Tricornutum. New Phytol. 2021, 229, 3208–3220. [Google Scholar] [CrossRef] [PubMed]
- Gall Trošelj, K.; Novak Kujundzic, R.; Ugarkovic, D. Polycomb Repressive Complex’s Evolutionary Conserved Function: The Role of EZH2 Status and Cellular Background. Clin. Epigenet. 2016, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Deevy, O.; Bracken, A.P. PRC2 Functions in Development and Congenital Disorders. Development 2019, 146, dev181354. [Google Scholar] [CrossRef] [Green Version]
- Lewis, Z.A. Polycomb Group Systems in Fungi: New Models for Understanding Polycomb Repressive Complex 2. Trends Genet. 2017, 33, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Derkacheva, M.; Hennig, L. Variations on a Theme: Polycomb Group Proteins in Plants. J. Exp. Bot. 2014, 65, 2769–2784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smits, A.H.; Jansen, P.W.T.C.; Poser, I.; Hyman, A.A.; Vermeulen, M. Stoichiometry of Chromatin-Associated Protein Complexes Revealed by Label-Free Quantitative Mass Spectrometry-Based Proteomics. Nucleic Acids Res. 2013, 41, e28. [Google Scholar] [CrossRef]
- Laugesen, A.; Højfeldt, J.W.; Helin, K. Molecular Mechanisms Directing PRC2 Recruitment and H3K27 Methylation. Mol. Cell 2019, 74, 8–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Su, T.; Wang, C.; Dong, L.; Liu, S.; Zhu, Y.; Hao, K.; Xia, Y.; Jiang, Q.; Qin, J. Rbbp4 Suppresses Premature Differentiation of Embryonic Stem Cells. Stem Cell Rep. 2021, 16, 566–581. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Min, J. Structure and Function of WD40 Domain Proteins. Protein Cell 2011, 2, 202–214. [Google Scholar] [CrossRef] [PubMed]
- Hauri, S.; Comoglio, F.; Seimiya, M.; Gerstung, M.; Glatter, T.; Hansen, K.; Aebersold, R.; Paro, R.; Gstaiger, M.; Beisel, C. A High-Density Map for Navigating the Human Polycomb Complexome. Cell Rep. 2016, 17, 583–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margueron, R.; Reinberg, D. The Polycomb Complex PRC2 and Its Mark in Life. Nature 2011, 469, 343–349. [Google Scholar] [CrossRef] [Green Version]
- Giner-Laguarda, N.; Vidal, M. Functions of Polycomb Proteins on Active Targets. Epigenomes 2020, 4, 17. [Google Scholar] [CrossRef]
- Kasinath, V.; Beck, C.; Sauer, P.; Poepsel, S.; Kosmatka, J.; Faini, M.; Toso, D.; Aebersold, R.; Nogales, E. JARID2 and AEBP2 Regulate PRC2 in the Presence of H2AK119ub1 and Other Histone Modifications. Science 2021, 371, eabc3393. [Google Scholar] [CrossRef]
- Han, Z.; Xing, X.; Hu, M.; Zhang, Y.; Liu, P.; Chai, J. Structural Basis of EZH2 Recognition by EED. Structure 2007, 15, 1306–1315. [Google Scholar] [CrossRef] [Green Version]
- Jiao, L.; Liu, X. Structural Basis of Histone H3K27 Trimethylation by an Active Polycomb Repressive Complex 2. Science 2015, 350, aac4383. [Google Scholar] [CrossRef] [Green Version]
- Bratkowski, M.; Yang, X.; Liu, X. Polycomb Repressive Complex 2 in an Autoinhibited State. J. Biol. Chem. 2017, 292, 13323–13332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uckelmann, M.; Davidovich, C. Not Just a Writer: PRC2 as a Chromatin Reader. Biochem. Soc. Trans. 2021, 49, 1159–1170. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, X.X.; Zhuang, Y.W.; Jiang, Y.; Melcher, K.; Xu, H.E. Structure of the PRC2 Complex and Application to Drug Discovery. Acta Pharmacol. Sin. 2017, 38, 963–976. [Google Scholar] [CrossRef]
- Moritz, L.E.; Trievel, R.C. Structure, Mechanism, and Regulation of Polycomb-Repressive Complex 2. J. Biol. Chem. 2018, 293, 13805–13814. [Google Scholar] [CrossRef] [Green Version]
- Chammas, P.; Mocavini, I.; Di Croce, L. Engaging Chromatin: PRC2 Structure Meets Function. Br. J. Cancer 2020, 122, 315–328. [Google Scholar] [CrossRef] [PubMed]
- The PyMOL Molecular Graphics System; Version 2.4.1; Schrödinger, LLC.: New York, NY, USA, 2020.
- Kasinath, V.; Nogales, E.; Beck, C.; Sauer, P.; Poepsel, S.; Kosmatka, J.; Faini, M.; Toso, D.; Aebersold, R. PRC2-AEBP2-JARID2 Bound to H2AK119ub1 Nucleosome. Available online: https://www.wwpdb.org/pdb?id=pdb_00006wkr (accessed on 20 November 2021). [CrossRef]
- Li, L.; Zhang, H.; Zhang, M.; Zhao, M.; Feng, L.; Luo, X.; Gao, Z.; Huang, Y.; Ardayfio, O.; Zhang, J.H.; et al. Discovery and Molecular Basis of a Diverse Set of Polycomb Repressive Complex 2 Inhibitors Recognition by EED. PLoS ONE 2017, 12, e0169855. [Google Scholar] [CrossRef] [PubMed]
- Blackledge, N.P.; Klose, R.J. The Molecular Principles of Gene Regulation by Polycomb Repressive Complexes. Nat. Rev. Mol. Cell Biol. 2021, 22, 815–833. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, D.H.; Liu, B.Y.; Shen, W.H.; Ruan, Y. Conservation and Diversification of Polycomb Repressive Complex 2 (PRC2) Proteins in the Green Lineage. Brief. Funct. Genom. 2017, 16, 106–119. [Google Scholar] [CrossRef]
- Chen, L.J.; Diao, Z.Y.; Specht, C.; Sung, Z.R. Molecular Evolution of VEF-Domain-Containing PcG Genes in Plants. Mol. Plant 2009, 2, 738–754. [Google Scholar] [CrossRef] [Green Version]
- Luo, M.; Platten, D.; Chaudhury, A.; Peacock, W.J.; Dennis, E.S. Expression, Imprinting, and Evolution of Rice Homologs of the Polycomb Group Genes. Mol. Plant 2009, 2, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Batista, R.A.; Köhler, C.; Hennig, L. Polycomb Repressive Complex 2-Mediated Histone Modification H3K27me3 Is Associated with Embryogenic Potential in Norway Spruce. J. Exp. Bot. 2020, 71, 6366–6378. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, C.; Zhao, Y.; Zhou, S.; Wang, W.; Zhou, D.X. The Rice Enhancer of Zeste [E(z)] Genes SDG711 and SDG718 Are Respectively Involved in Long Day and Short Day Signaling to Mediate the Accurate Photoperiod Control of Flowering Time. Front. Plant Sci. 2014, 5, 591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.; Ma, X.; Feng, Y.; Tian, Q.; Wang, Y.; Xu, N.; Tang, J.; Wang, G. Updating and Interaction of Polycomb Repressive Complex 2 Components in Maize (Zea Mays). Planta 2019, 250, 573–588. [Google Scholar] [CrossRef]
- Cheng, X.; Pan, M.; Zhiguo, E.; Zhou, Y.; Niu, B.; Chen, C. The Maternally Expressed Polycomb Group Gene OsEMF2a Is Essential for Endosperm Cellularization and Imprinting in Rice. Plant Commun. 2020, 2, 100092. [Google Scholar] [CrossRef] [PubMed]
- Conrad, L.J.; Khanday, I.; Johnson, C.; Guiderdoni, E.; An, G.; Vijayraghavan, U.; Sundaresan, V. The Polycomb Group Gene EMF2B Is Essential for Maintenance of Floral Meristem Determinacy in Rice. Plant J. 2014, 80, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Strejčková, B.; Čegan, R.; Pecinka, A.; Milec, Z.; Šafář, J. Identification of Polycomb Repressive Complex 1 and 2 Core Components in Hexaploid Bread Wheat. BMC Plant Biol. 2020, 20, 175. [Google Scholar] [CrossRef]
- Higgins, J.A.; Bailey, P.C.; Laurie, D.A. Comparative Genomics of Flowering Time Pathways Using Brachypodium Distachyon as a Model for the Temperate Grasses. PLoS ONE 2010, 5, e10065. [Google Scholar] [CrossRef]
- Lomax, A.; Woods, D.P.; Dong, Y.; Bouché, F.; Rong, Y.; Mayer, K.S.; Zhong, X.; Amasino, R.M. An Ortholog of CURLY LEAF/ENHANCER OF ZESTE like-1 Is Required for Proper Flowering in Brachypodium Distachyon. Plant J. 2018, 93, 871–882. [Google Scholar] [CrossRef] [Green Version]
- Kapazoglou, A.; Tondelli, A.; Papaefthimiou, D.; Ampatzidou, H.; Francia, E.; Stanca, M.A.; Bladenopoulos, K.; Tsaftaris, A.S. Epigenetic Chromatin Modifiers in Barley: IV. The Study of Barley Polycomb Group (PcG) Genes during Seed Development and in Response to External ABA. BMC Plant Biol. 2010, 10, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonosaki, K.; Kinoshita, T. Possible Roles for Polycomb Repressive Complex 2 in Cereal Endosperm. Front. Plant Sci. 2015, 6, 144. [Google Scholar] [CrossRef] [Green Version]
- Derkacheva, M.; Steinbach, Y.; Wildhaber, T.; Mozgová, I.; Mahrez, W.; Nanni, P.; Bischof, S.; Gruissem, W.; Hennig, L. Arabidopsis MSI1 Connects LHP1 to PRC2 Complexes. EMBO J. 2013, 32, 2073–2085. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Deton Cabanillas, A.F.; Veluchamy, A.; Bowler, C.; Vieira, F.R.J.; Tirichine, L. Probing the Diversity of Polycomb and Trithorax Proteins in Cultured and Environmentally Sampled Microalgae. Front. Mar. Sci. 2020, 7, 189. [Google Scholar] [CrossRef]
- Bourdareau, S.; Tirichine, L.; Lombard, B.; Loew, D.; Scornet, D.; Wu, Y.; Coelho, S.M.; Cock, J.M. Histone Modifications during the Life Cycle of the Brown Alga Ectocarpus. Genome Biol. 2021, 22, 12. [Google Scholar] [CrossRef]
- Rzeszutek, I.; Maurer-Alcalá, X.X.; Nowacki, M. Programmed Genome Rearrangements in Ciliates. Cell. Mol. Life Sci. 2020, 77, 4615–4629. [Google Scholar] [CrossRef]
- Frapporti, A.; Miró Pina, C.; Arnaiz, O.; Holoch, D.; Kawaguchi, T.; Humbert, A.; Eleftheriou, E.; Lombard, B.; Loew, D.; Sperling, L.; et al. The Polycomb Protein Ezl1 Mediates H3K9 and H3K27 Methylation to Repress Transposable Elements in Paramecium. Nat. Commun. 2019, 10, 2710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Zhao, X.; Mao, F.; Basrur, V.; Ueberheide, B.; Chait, B.T.; David Allis, C.; Taverna, S.D.; Gao, S.; Wang, W.; et al. A Polycomb Repressive Complex Is Required for RNAi-Mediated Heterochromatin Formation and Dynamic Distribution of Nuclear Bodies. Nucleic Acids Res. 2021, 49, 5407–5425. [Google Scholar] [CrossRef]
- Nabeel-Shah, S.; Garg, J.; Saettone, A.; Ashraf, K.; Lee, H.; Wahab, S.; Ahmed, N.; Fine, J.; Derynck, J.; Pu, S.; et al. Functional Characterization of RebL1 Highlights the Evolutionary Conservation of Oncogenic Activities of the RBBP4/7 Orthologue in Tetrahymena Thermophila. Nucleic Acids Res. 2021, 49, 6196–6212. [Google Scholar] [CrossRef]
- Ridenour, J.B.; Möller, M.; Freitag, M. Polycomb Repression without Bristles: Facultative Heterochromatin and Genome Stability in Fungi. Genes 2020, 11, 638. [Google Scholar] [CrossRef]
- Connolly, L.R.; Smith, K.M.; Freitag, M. The Fusarium Graminearum Histone H3 K27 Methyltransferase KMT6 Regulates Development and Expression of Secondary Metabolite Gene Clusters. PLoS Genet. 2013, 9, e1003916. [Google Scholar] [CrossRef] [Green Version]
- Jamieson, K.; Rountree, M.R.; Lewis, Z.A.; Stajich, J.E.; Selker, E.U. Regional Control of Histone H3 Lysine 27 Methylation in Neurospora. Proc. Natl. Acad. Sci. USA 2013, 110, 6027–6032. [Google Scholar] [CrossRef] [Green Version]
- McNaught, K.J.; Wiles, E.T.; Selker, E.U. Identification of a PRC2 Accessory Subunit Required for Subtelomeric H3K27 Methylation in Neurospora Crassa. Mol. Cell. Biol. 2020, 40, e00003-20. [Google Scholar] [CrossRef]
- Sebé-Pedrós, A.; Ballaré, C.; Parra-Acero, H.; Chiva, C.; Tena, J.J.; Sabidó, E.; Gómez-Skarmeta, J.L.; Di Croce, L.; Ruiz-Trillo, I. The Dynamic Regulatory Genome of Capsaspora and the Origin of Animal Multicellularity. Cell 2016, 165, 1224–1237. [Google Scholar] [CrossRef] [Green Version]
- Gaiti, F.; Jindrich, K.; Fernandez-Valverde, S.L.; Roper, K.E.; Degnan, B.M.; Tanurdžić, M. Landscape of Histone Modifications in a Sponge Reveals the Origin of Animal Cis-Regulatory Complexity. eLife 2017, 6, e22194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genikhovich, G.; Kürn, U.; Hemmrich, G.; Bosch, T.C.G. Discovery of Genes Expressed in Hydra Embryogenesis. Dev. Biol. 2006, 289, 466–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, A.; Gungi, A.; Reddy, P.C.; Galande, S. Epigenetic Regulation in Hydra: Conserved and Divergent Roles. Front. Cell Dev. Biol. 2021, 9, 663208. [Google Scholar] [CrossRef] [PubMed]
- PREDICTED: Polycomb Protein Suz12-like [Hydra Vulgaris]. Available online: Https://Www.Ncbi.Nlm.Nih.Gov/Protein/XP_012556196.1?Report=genbank&log$=prottop&blast_rank=1&RID=R2KAB9PA013 (accessed on 21 October 2021).
- PREDICTED: Histone-Binding Protein RBBP7 [Hydra Vulgaris]. Available online: Https://Www.Ncbi.Nlm.Nih.Gov/Protein/XP_012554465.1?Report=genbank&log$=prottop&blast_rank=1&RID=R2KUGECN013 (accessed on 21 October 2021).
- Herz, H.-M.; Mohan, M.; Garrett, A.S.; Miller, C.; Casto, D.; Zhang, Y.; Seidel, C.; Haug, J.S.; Florens, L.; Washburn, M.P.; et al. Polycomb Repressive Complex 2-Dependent and -Independent Functions of Jarid2 in Transcriptional Regulation in Drosophila. Mol. Cell. Biol. 2012, 32, 1683–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolhuis, B.; Muijrers, I.; de Wit, E.; Teunissen, H.; Talhout, W.; van Steensel, B.; van Lohuizen, M. Genome-Wide Profiling of PRC1 and PRC2 Polycomb Chromatin Binding in Drosophila Melanogaster. Nat. Genet. 2006, 38, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Siebold, A.P.; Banerjee, R.; Tie, F.; Kiss, D.L.; Moskowitz, J.; Harte, P.J. Polycomb Repressive Complex 2 and Trithorax Modulate Drosophila Longevity and Stress Resistance. Proc. Natl. Acad. Sci. USA 2010, 107, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bender, L.B.; Cao, R.; Zhang, Y.; Strome, S. The MES-2/MES-3/MES-6 Complex and Regulation of Histone H3 Methylation in C. Elegans. Curr. Biol. 2004, 14, 1639–1643. [Google Scholar] [CrossRef] [Green Version]
- Gaydos, L.J.; Wang, W.; Strome, S. H3K27me and PRC2 Transmit a Memory of Repression across Generations and during Development. Science 2014, 345, 1515–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooun, A.; Gajiwala, K.S.; Deng, Y.L.; Liu, W.; Bolaños, B.; Bingham, P.; He, Y.A.; Diehl, W.; Grable, N.; Kung, P.P.; et al. Polycomb Repressive Complex 2 Structure with Inhibitor Reveals a Mechanism of Activation and Drug Resistance. Nat. Commun. 2016, 7, 11384. [Google Scholar] [CrossRef]
- PREDICTED: LOW QUALITY PROTEIN: Polycomb Protein SUZ12 [Anolis Carolinensis]. Available online: Https://Www.Ncbi.Nlm.Nih.Gov/Protein/XP_008112590.1?Report=genbank&log$=prottop&blast_rank=1&RID=R2PMHJ83016 (accessed on 21 October 2021).
- PREDICTED: Polycomb Protein EED [Anolis Carolinensis]. Available online: Https://Www.Ncbi.Nlm.Nih.Gov/Protein/XP_003219406.1?Report=genbank&log$=prottop&blast_rank=1&RID=R2PDKH5B013 (accessed on 21 October 2021).
- PREDICTED: Histone-Binding Protein RBBP4 Isoform X2 [Anolis Carolinensis]. Available online: Https://Www.Ncbi.Nlm.Nih.Gov/Protein/XP_003230003.3?Report=genbank&log$=prottop&blast_rank=1&RID=R2P2UE3N013 (accessed on 21 October 2021).
- Grossniklaus, U.; Paro, R. Transcriptional Silencing by Polycomb Group Proteins Cellular Memory. Cold Spring Harb. Perspect. Biol. 2014, 6, a019331. [Google Scholar] [CrossRef] [Green Version]
- He, A.; Ma, Q.; Cao, J.; von Gise, A.; Zhou, P.; Xie, H.; Zhang, B.; Hsing, M.; Christodoulou, D.C.; Cahan, P.; et al. Polycomb Repressive Complex 2 Regulates Normal Development of the Mouse Heart. Circ. Res. 2012, 110, 406–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennig, L.; Taranto, P.; Walser, M.; Schönrock, N.; Gruissem, W. Arabidopsis MSI1 Is Required for Epigenetic Maintenance of Reproductive Development. Development 2003, 130, 2555–2565. [Google Scholar] [CrossRef] [Green Version]
- Köhler, C.; Hennig, L.; Bouveret, R.; Gheyselinck, J.; Grossniklaus, U.; Gruissem, W. Arabidopsis MSI1 Is a Component of the MEA/FIE Polycomb Group Complex and Required for Seed Development. EMBO J. 2003, 22, 4804–4814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennig, L.; Bouveret, R.; Gruissem, W. MSI1-like Proteins: An Escort Service for Chromatin Assembly and Remodeling Complexes. Trends Cell Biol. 2005, 15, 295–302. [Google Scholar] [CrossRef]
- Doyle, M.R.; Amasino, R.M. A Single Amino Acid Change in the Enhancer of Zeste Ortholog CURLY LEAF Results in Vernalization-Independent, Rapid Flowering in Arabidopsis. Plant Physiol. 2009, 151, 1688–1697. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Hu, T.; Smith, M.R.; Poethig, R.S. Epigenetic Regulation of Vegetative Phase Change in Arabidopsis. Plant Cell 2016, 28, 28–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schubert, D.; Primavesi, L.; Bishopp, A.; Roberts, G.; Doonan, J.; Jenuwein, T.; Goodrich, J. Silencing by Plant Polycomb-Group Genes Requires Dispersed Trimethylation of Histone H3 at Lysine 27. EMBO J. 2006, 25, 4638–4649. [Google Scholar] [CrossRef] [PubMed]
- Chanvivattana, Y.; Bishopp, A.; Schubert, D.; Stock, C.; Moon, Y.H.; Sung, Z.R.; Goodrich, J. Interaction of Polycomb-Group Proteins Controlling Flowering in Arabidopsis. Development 2004, 131, 5263–5276. [Google Scholar] [CrossRef] [Green Version]
- Aichinger, E.; Villar, C.B.R.; Farrona, S.; Reyes, J.C.; Hennig, L.; Köhler, C. CHD3 Proteins and Polycomb Group Proteins Antagonistically Determine Cell Identity in Arabidopsis. PLoS Genet. 2009, 5, e1000605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aichinger, E.; Villar, C.B.R.; Di Mambro, R.; Sabatini, S.; Köhler, C. The CHD3 Chromatin Remodeler PICKLE and Polycomb Group Proteins Antagonistically Regulate Meristem Activity in the Arabidopsis Root. Plant Cell 2011, 23, 1047–1060. [Google Scholar] [CrossRef] [Green Version]
- Simonini, S.; Bemer, M.; Bencivenga, S.; Gagliardini, V.; Pires, N.D.; Desvoyes, B.; van der Graaff, E.; Gutierrez, C.; Grossniklaus, U. The Polycomb Group Protein MEDEA Controls Cell Proliferation and Embryonic Patterning in Arabidopsis. Dev. Cell 2021, 56, 1945–1960.e7. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.H.; Chen, L.; Pan, R.L.; Chang, H.S.; Zhu, T.; Maffeo, D.M.; Sung, Z.R. Erratum: EMF Genes Maintain Vegetative Development by Repressing the Flower Program in Arabidopsis. Plant Cell 2003, 15, 681–693. [Google Scholar] [CrossRef] [Green Version]
- De Lucia, F.; Crevillen, P.; Jones, A.M.E.; Greb, T.; Dean, C. A PHD-Polycomb Repressive Complex 2 Triggers the Epigenetic Silencing of FLC during Vernalization. Proc. Natl. Acad. Sci. USA 2008, 105, 16831–16836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Wang, D.; Zhang, H.; Skaggs, M.I.; Lloyd, A.; Ran, D.; An, L.; Schumaker, K.S.; Drews, G.N.; Yadegari, R. FERTILIZATION-INDEPENDENT SEED-Polycomb Repressive Complex 2 Plays a Dual Role in Regulating Type i MADS-Box Genes in Early Endosperm Development. Plant Physiol. 2018, 177, 285–299. [Google Scholar] [CrossRef] [Green Version]
- Bloomer, R.H.; Hutchison, C.E.; Bäurle, I.; Walker, J.; Fang, X.; Perera, P.; Velanis, C.N.; Gümüs, S.; Spanos, C.; Rappsilber, J.; et al. The Arabidopsis Epigenetic Regulator ICU11 as an Accessory Protein of Polycomb Repressive Complex 2. Proc. Natl. Acad. Sci. USA 2020, 117, 16660. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Y.; Krause, K.; Yang, T.; Dongus, J.A.; Zhang, Y.; Turck, F. Telobox Motifs Recruit CLF/SWN-PRC2 for H3K27me3 Deposition via TRB Factors in Arabidopsis. Nat. Genet. 2018, 50, 638–644. [Google Scholar] [CrossRef]
- Hohenstatt, M.L.; Mikulski, P.; Komarynets, O.; Klose, C.; Kycia, I.; Jeltsch, A.; Farrona, S.; Schubert, D. PWWP-DOMAIN INTERACTOR OF POLYCOMBS1 Interacts with Polycomb-Group Proteins and Histones and Regulates Arabidopsis Flowering and Development. Plant Cell 2018, 30, 117–133. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Song, X.; Zhang, L.; Yu, Y.; Liang, Z.; Lei, Y.; Ruan, J.; Tan, B.; Liu, J.; Li, C. The Transcriptional Repressors VAL1 and VAL2 Recruit PRC2 for Genome-Wide Polycomb Silencing in Arabidopsis. Nucleic Acids Res. 2021, 49, 98–113. [Google Scholar] [CrossRef]
- Zhang, P.; Zhu, C.; Geng, Y.; Wang, Y.; Yang, Y.; Liu, Q.; Guo, W.; Chachar, S.; Riaz, A.; Yan, S.; et al. Rice and Arabidopsis Homologs of Yeast CHROMOSOME TRANSMISSION FIDELITY PROTEIN 4 Commonly Interact with Polycomb Complexes but Exert Divergent Regulatory Functions. Plant Cell 2021, 33, 1417–1429. [Google Scholar] [CrossRef]
- Velanis, C.N.; Perera, P.; Thomson, B.; de Leau, E.; Liang, S.C.; Hartwig, B.; Förderer, A.; Thornton, H.; Arede, P.; Chen, J.; et al. The Domesticated Transposase ALP2 Mediates Formation of a Novel Polycomb Protein Complex by Direct Interaction with MSI1, a Core Subunit of Polycomb Repressive Complex 2 (PRC2). PLoS Genet. 2020, 16, e1008681. [Google Scholar] [CrossRef]
- Zhou, Y.; Tergemina, E.; Cui, H.; Förderer, A.; Hartwig, B.; Velikkakam James, G.; Schneeberger, K.; Turck, F. Ctf4-Related Protein Recruits LHP1-PRC2 to Maintain H3K27me3 Levels in Dividing Cells in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2017, 114, 4833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, Y.; Feng, S.; LeBlanc, C.A.; Bernatavichute, Y.V.; Stroud, H.; Cokus, S.; Johnson, L.M.; Pellegrini, M.; Jacobsen, S.E.; Michaels, S.D. ATXR5 and ATXR6 Are H3K27 Monomethyltransferases Required for Chromatin Structure and Gene Silencing. Nat. Struct. Mol. Biol. 2009, 16, 763–768. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, Y.B.; Kahn, T.G.; Nix, D.A.; Li, X.-Y.; Bourgon, R.; Biggin, M.; Pirrotta, V. Genome-Wide Analysis of Polycomb Targets in Drosophila Melanogaster. Nat. Genet. 2006, 38, 700–705. [Google Scholar] [CrossRef]
- Crispatzu, G.; Rehimi, R.; Pachano, T.; Bleckwehl, T.; Cruz-Molina, S.; Xiao, C.; Mahabir, E.; Bazzi, H.; Rada-Iglesias, A. The Chromatin, Topological and Regulatory Properties of Pluripotency-Associated Poised Enhancers Are Conserved in Vivo. Nat. Commun. 2021, 12, 4344. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.L.; Beckedorff, F.; Zhang, Y.; Garcia-Huidobro, J.; Jiang, H.; Colaprico, A.; Bilbao, D.; Figueroa, M.E.; LaCava, J.; Shiekhattar, R.; et al. Polycomb Complexes Associate with Enhancers and Promote Oncogenic Transcriptional Programs in Cancer through Multiple Mechanisms. Nat. Commun. 2018, 9, 3377. [Google Scholar] [CrossRef] [Green Version]
- Roudier, F.; Ahmed, I.; Bérard, C.; Sarazin, A.; Mary-Huard, T.; Cortijo, S.; Bouyer, D.; Caillieux, E.; Duvernois-Berthet, E.; Al-Shikhley, L.; et al. Integrative Epigenomic Mapping Defines Four Main Chromatin States in Arabidopsis. EMBO J. 2011, 30, 1928–1938. [Google Scholar] [CrossRef] [Green Version]
- Sequeira-Mendes, J.; Aragüez, I.; Peiró, R.; Mendez-Giraldez, R.; Zhang, X.; Jacobsen, S.E.; Bastolla, U.; Gutierrez, C. The Functional Topography of the Arabidopsis Genome Is Organized in a Reduced Number of Linear Motifs of Chromatin States. Plant Cell 2014, 26, 2351–2366. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.; Nützmann, H.W.; Macdonald, J.T.; Moore, B.; Field, B.; Berriri, S.; Trick, M.; Rosser, S.J.; Kumar, S.V.; Freemont, P.S.; et al. Delineation of Metabolic Gene Clusters in Plant Genomes by Chromatin Signatures. Nucleic Acids Res. 2016, 44, 2255–2265. [Google Scholar] [CrossRef] [PubMed]
- Lafos, M.; Kroll, P.; Hohenstatt, M.L.; Thorpe, F.L.; Clarenz, O.; Schubert, D. Dynamic Regulation of H3K27 Trimethylation during Arabidopsis Differentiation. PLoS Genet. 2011, 7, e1002040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellegarde, F.; Herbert, L.; Séré, D.; Caillieux, E.; Boucherez, J.; Fizames, C.; Roudier, F.; Gojon, A.; Martin, A. Polycomb Repressive Complex 2 Attenuates the Very High Expression of the Arabidopsis Gene NRT2.1. Sci. Rep. 2018, 8, 7905. [Google Scholar] [CrossRef] [Green Version]
- Park, E.Y.; Tsuyuki, K.M.; Hu, F.; Lee, J.; Jeong, J. PRC2-Mediated H3K27me3 Contributes to Transcriptional Regulation of FIT-Dependent Iron Deficiency Response. Front. Plant Sci. 2019, 10, 627. [Google Scholar] [CrossRef]
- Sani, E.; Herzyk, P.; Perrella, G.; Colot, V.; Amtmann, A. Hyperosmotic Priming of Arabidopsis Seedlings Establishes a Long-Term Somatic Memory Accompanied by Specific Changes of the Epigenome. Genome Biol. 2013, 14, R59. [Google Scholar] [CrossRef] [Green Version]
- Chica, C.; Louis, A.; Roest Crollius, H.; Colot, V.; Roudier, F. Comparative Epigenomics in the Brassicaceae Reveals Two Evolutionarily Conserved Modes of PRC2-Mediated Gene Regulation. Genome Biol. 2017, 18, 207. [Google Scholar] [CrossRef] [Green Version]
- Lämke, J.; Bäurle, I. Epigenetic and Chromatin-Based Mechanisms in Environmental Stress Adaptation and Stress Memory in Plants. Genome Biol. 2017, 18, 124. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-M.; Sasaki, T.; Ueda, M.; Sako, K.; Seki, M. Chromatin Changes in Response to Drought, Salinity, Heat, and Cold Stresses in Plants. Front. Plant Sci. 2015, 6, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, Y.; Sawikowska, A.; Neumann, M.; Posé, D.; Capovilla, G.; Langenecker, T.; Neher, R.A.; Krajewski, P.; Schmid, M. Temporal Dynamics of Gene Expression and Histone Marks at the Arabidopsis Shoot Meristem during Flowering. Nat. Commun. 2017, 8, 207. [Google Scholar] [CrossRef]
- Veluchamy, A.; Rastogi, A.; Lin, X.; Lombard, B.; Murik, O.; Thomas, Y.; Dingli, F.; Rivarola, M.; Ott, S.; Liu, X.; et al. An Integrative Analysis of Post-Translational Histone Modifications in the Marine Diatom Phaeodactylum Tricornutum. Genome Biol. 2015, 16, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, S.A.; Tanizawa, Y.; Galik, B.; Wang, N.; Ito, T.; Mochizuki, T.; Akimcheva, S.; Bowman, J.L.; Cognat, V.; Maréchal-Drouard, L.; et al. Chromatin Organization in Early Land Plants Reveals an Ancestral Association between H3K27me3, Transposons, and Constitutive Heterochromatin. Curr. Biol. 2020, 30, 573–588.e7. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Romero, J.; Jiang, H.; Santos-González, J.; Köhler, C. Parental Epigenetic Asymmetry of PRC2-Mediated Histone Modifications in the Arabidopsis Endosperm. EMBO J. 2016, 35, 1298–1311. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Wang, H.; Cai, Y.; Wang, H.; Niu, K.; Wu, X.; Ma, H.; Yang, Y.; Tong, W.; Liu, F.; et al. Epigenetic Drift of H3K27me3 in Aging Links Glycolysis to Healthy Longevity in Drosophila. Elife 2018, 7, e35368. [Google Scholar] [CrossRef]
- Trim Galore. Available online: http://www.Bioinformatics.Babraham.Ac.Uk/Projects/Trim_galore/ (accessed on 7 January 2022).
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome Project Data Processing Subgroup the Sequence Alignment/Map Format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, J.T.; Thorvaldsdóttir, H.; Wenger, A.M.; Zehir, A.; Mesirov, J.P. Variant Review with the Integrative Genomics Viewer. Cancer Res. 2017, 77, e31–e34. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, F.; Ryan, D.P.; Grüning, B.; Bhardwaj, V.; Kilpert, F.; Richter, A.S.; Heyne, S.; Dündar, F.; Manke, T. DeepTools2: A next Generation Web Server for Deep-Sequencing Data Analysis. Nucleic Acids Res. 2016, 44, W160–W165. [Google Scholar] [CrossRef]
- Bredesen, B.A.; Rehmsmeier, M. DNA Sequence Models of Genome-Wide Drosophila Melanogaster Polycomb Binding Sites Improve Generalization to Independent Polycomb Response Elements. Nucleic Acids Res. 2019, 47, 7781–7797. [Google Scholar] [CrossRef] [PubMed]
- Erokhin, M.; Gorbenko, F.; Lomaev, D.; Mazina, M.Y.; Mikhailova, A.; Garaev, A.K.; Parshikov, A.; Vorobyeva, N.E.; Georgiev, P.; Schedl, P.; et al. Boundaries Potentiate Polycomb Response Element-Mediated Silencing. BMC Biol. 2021, 19, 113. [Google Scholar] [CrossRef] [PubMed]
- Kassis, J.A.; Brown, J.L. Polycomb Group Response Elements in Drosophila and Vertebrates. Adv. Genet. 2013, 81, 83–118. [Google Scholar] [CrossRef] [Green Version]
- Schorderet, P.; Lonfat, N.; Darbellay, F.; Tschopp, P.; Gitto, S.; Soshnikova, N.; Duboule, D. A Genetic Approach to the Recruitment of PRC2 at the HoxD Locus. PLoS Genet. 2013, 9, e1003951. [Google Scholar] [CrossRef] [Green Version]
- Ray, P.; De, S.; Mitra, A.; Bezstarosti, K.; Demmers, J.A.A.; Pfeifer, K.; Kassis, J.A. Combgap Contributes to Recruitment of Polycomb Group Proteins in Drosophila. Proc. Natl. Acad. Sci. USA 2016, 113, 3826–3831. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Kirk, B.; Zeng, J.; Ma, J.; Wang, Q. Three Classes of Response Elements for Human PRC2 and MLL1/2-Trithorax Complexes. Nucleic Acids Res. 2018, 46, 8848–8864. [Google Scholar] [CrossRef] [Green Version]
- Lynch, M.D.; Smith, A.J.H.; De Gobbi, M.; Flenley, M.; Hughes, J.R.; Vernimmen, D.; Ayyub, H.; Sharpe, J.A.; Sloane-Stanley, J.A.; Sutherland, L.; et al. An Interspecies Analysis Reveals a Key Role for Unmethylated CpG Dinucleotides in Vertebrate Polycomb Complex Recruitment. EMBO J. 2012, 31, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Jiao, L.; Liu, X.; Yang, X.; Liu, X. A Dimeric Structural Scaffold for PRC2-PCL Targeting to CpG Island Chromatin. Mol. Cell 2020, 77, 1265–1278. [Google Scholar] [CrossRef] [PubMed]
- Mendenhall, E.M.; Koche, R.P.; Truong, T.; Zhou, V.W.; Issac, B.; Chi, A.S.; Ku, M.; Bernstein, B.E. GC-Rich Sequence Elements Recruit PRC2 in Mammalian ES Cells. PLoS Genet. 2010, 6, e1001244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodha, M.; Marco, C.F.; Timmermans, M.C.P. The ASYMMETRIC LEAVES Complex Maintains Repression of KNOX Homeobox Genes via Direct Recruitment of Polycomb-Repressive Complex2. Genes Dev. 2013, 27, 596–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, N.; Dubreucq, B.; Roudier, F.; Dubos, C.; Lepiniec, L. Transcriptional Regulation of Arabidopsis LEAFY COTYLEDON2 Involves RLE, a Cis-Element That Regulates Trimethylation of Histone H3 at Lysine-27. Plant Cell 2011, 23, 4065–4078. [Google Scholar] [CrossRef] [Green Version]
- Xiao, J.; Jin, R.; Yu, X.; Shen, M.; Wagner, J.D.; Pai, A.; Song, C.; Zhuang, M.; Klasfeld, S.; He, C.; et al. Cis and Trans Determinants of Epigenetic Silencing by Polycomb Repressive Complex 2 in Arabidopsis. Nat. Genet. 2017, 49, 1546–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, Y.; Zou, M.; Sun, X.; He, B.; Xu, X.; Liu, Y.; Zhang, L.; Chi, W. BASIC PENTACYSTEINE Proteins Repress Abscisic Acid INSENSITIVE 4 Expression via Direct Recruitment of the Polycomb-Repressive Complex 2 in Arabidopsis Root Development. Plant Cell Physiol. 2017, 58, 607–621. [Google Scholar] [CrossRef] [Green Version]
- Fouracre, J.P.; He, J.; Chen, V.J.; Sidoli, S.; Scott Poethig, R. VAL Genes Regulate Vegetative Phase Change via MiR156-Dependent and Independent Mechanisms. PLoS Genet. 2021, 17, e1009626. [Google Scholar] [CrossRef]
- Brockdorff, N. Noncoding RNA and Polycomb Recruitment. RNA 2013, 19, 429–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidovich, C.; Cech, T.R. The Recruitment of Chromatin Modifiers by Long Noncoding RNAs: Lessons from PRC2. Rna 2015, 21, 2007–2022. [Google Scholar] [CrossRef] [Green Version]
- Skourti-Stathaki, K.; Torlai Triglia, E.; Warburton, M.; Voigt, P.; Bird, A.; Pombo, A. R-Loops Enhance Polycomb Repression at a Subset of Developmental Regulator Genes. Mol. Cell 2019, 73, 930–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allison, D.F.; Wang, G.G. R-Loops: Formation, Function, and Relevance to Cell Stress. Cell Stress 2019, 3, 38–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alecki, C.; Chiwara, V.; Sanz, L.A.; Grau, D.; Arias Pérez, O.; Boulier, E.L.; Armache, K.J.; Chédin, F.; Francis, N.J. RNA-DNA Strand Exchange by the Drosophila Polycomb Complex PRC2. Nat. Commun. 2020, 11, 1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.B.; Chen, H.V.; Acharya, D.; Rando, O.J.; Fazzio, T.G. R Loops Regulate Promoter-Proximal Chromatin Architecture and Cellular Differentiation. Nat. Struct. Mol. Biol. 2015, 22, 999–1007. [Google Scholar] [CrossRef]
- Tian, Y.; Zheng, H.; Zhang, F.; Wang, S.; Ji, X.; Xu, C.; He, Y.; Ding, Y. PRC2 Recruitment and H3K27me3 Deposition at FLC Require FCA Binding of COOLAIR. Sci. Adv. 2019, 5, eaau7246. [Google Scholar] [CrossRef] [Green Version]
- Csorba, T.; Questa, J.I.; Sun, Q.; Dean, C. Antisense COOLAIR Mediates the Coordinated Switching of Chromatin States at FLC during Vernalization. Proc. Natl. Acad. Sci. USA 2014, 111, 16160–16165. [Google Scholar] [CrossRef] [Green Version]
- Ariel, F.; Jegu, T.; Latrasse, D.; Romero-Barrios, N.; Christ, A.; Benhamed, M.; Crespi, M. Noncoding Transcription by Alternative RNA Polymerases Dynamically Regulates an Auxin-Driven Chromatin Loop. Mol. Cell 2014, 55, 383–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.; Rothbart, S.B.; Lu, R.; Xu, B.; Chen, W.-Y.; Tripathy, A.; Rockowitz, S.; Zheng, D.; Patel, D.J.; Allis, C.D.; et al. An H3K36 Methylation-Engaging Tudor Motif of Polycomb-like Proteins Mediates PRC2 Complex Targeting. Mol. Cell 2013, 49, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Schmitges, F.W.; Prusty, A.B.; Faty, M.; Stützer, A.; Lingaraju, G.M.; Aiwazian, J.; Sack, R.; Hess, D.; Li, L.; Zhou, S.; et al. Histone Methylation by PRC2 Is Inhibited by Active Chromatin Marks. Mol. Cell 2011, 42, 330–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, W.; Xu, M.; Huang, C.; Liu, N.; Chen, S.; Zhu, B. H3K36 Methylation Antagonizes PRC2-Mediated H3K27 Methylation. J. Biol. Chem. 2011, 286, 7983–7989. [Google Scholar] [CrossRef] [Green Version]
- Finogenova, K.; Bonnet, J.; Poepsel, S.; Schäfer, I.B.; Finkl, K.; Schmid, K.; Litz, C.; Strauss, M.; Benda, C.; Müller, J. Structural Basis for PRC2 Decoding of Active Histone Methylation Marks H3K36me2/3. eLife 2020, 9, e61964. [Google Scholar] [CrossRef] [PubMed]
- Van Kruijsbergen, I.; Hontelez, S.; Veenstra, G.J.C. Recruiting Polycomb to Chromatin. Int. J. Biochem. Cell Biol. 2015, 67, 177–187. [Google Scholar] [CrossRef] [Green Version]
- Ballaré, C.; Lange, M.; Lapinaite, A.; Martin, G.M.; Morey, L.; Pascual, G.; Liefke, R.; Simon, B.; Shi, Y.; Gozani, O.; et al. Phf19 Links Methylated Lys36 of Histone H3 to Regulation of Polycomb Activity. Nat. Struct. Mol. Biol. 2012, 19, 1257–1265. [Google Scholar] [CrossRef] [Green Version]
- Blanco, E.; González-Ramírez, M.; Alcaine-Colet, A.; Aranda, S.; Croce, L.D. The Bivalent Genome: Characterization, Structure, and Regulation. Trends Genet. 2020, 36, 118–131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Guan, P.; Zhao, L.; Ma, M.; Xie, L.; Li, Y.; Zheng, R.; Ouyang, W.; Wang, S.; Li, H.; et al. Asymmetric Epigenome Maps of Subgenomes Reveal Imbalanced Transcription and Distinct Evolutionary Trends in Brassica Napus. Mol. Plant 2021, 14, 604–619. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Kong, D.; Jin, B.; Smolke, C.D.; Rhee, S.Y. A Novel Bivalent Chromatin Associates with Rapid Induction of Camalexin Biosynthesis Genes in Response to a Pathogen Signal in Arabidopsis. elife 2021, 10, e69508. [Google Scholar] [CrossRef] [PubMed]
- Hansen, K.H.; Bracken, A.P.; Pasini, D.; Dietrich, N.; Gehani, S.S.; Monrad, A.; Rappsilber, J.; Lerdrup, M.; Helin, K. A Model for Transmission of the H3K27me3 Epigenetic Mark. Nat. Cell Biol. 2008, 10, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Margueron, R.; Justin, N.; Ohno, K.; Sharpe, M.L.; Son, J.; Iii, W.J.D.; Voigt, P.; Martin, S.; Taylor, W.R.; Marco, V.D.; et al. Role of the Polycomb Protein EED in the Propagation of Repressive Histone Marks. Nature 2009, 461, 762–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Exner, V.; Aichinger, E.; Shu, H.; Wildhaber, T.; Alfarano, P.; Caflisch, A.; Gruissem, W.; Köhler, C.; Hennig, L. The Chromodomain of LIKE HETEROCHROMATIN PROTEIN 1 Is Essential for H3K27me3 Binding and Function during Arabidopsis Development. PLoS ONE 2009, 4, e0005335. [Google Scholar] [CrossRef] [Green Version]
- Cooper, S.; Dienstbier, M.; Hassan, R.; Schermelleh, L.; Sharif, J.; Blackledge, N.P.; De Marco, V.; Elderkin, S.; Koseki, H.; Klose, R.; et al. Targeting Polycomb to Pericentric Heterochromatin in Embryonic Stem Cells Reveals a Role for H2AK119u1 in PRC2 Recruitment. Cell Rep. 2014, 7, 1456–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrinić, P.; Szczurek, A.T.; Klose, R.J. PRC1 Drives Polycomb-Mediated Gene Repression by Controlling Transcription Initiation and Burst Frequency. Nat. Struct. Mol. Biol. 2021, 28, 811–824. [Google Scholar] [CrossRef]
- Blackledge, N.P.; Rose, N.R.; Klose, R.J. Targeting Polycomb Systems to Regulate Gene Expression: Modifications to a Complex Story. Nat. Rev. Mol. Cell Biol. 2015, 16, 643–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbour, H.; Daou, S.; Hendzel, M.; Affar, E.B. Polycomb Group-Mediated Histone H2A Monoubiquitination in Epigenome Regulation and Nuclear Processes. Nat. Commun. 2020, 11, 5947. [Google Scholar] [CrossRef]
- Yin, X.; Romero-Campero, F.J.; de Los Reyes, P.; Yan, P.; Yang, J.; Tian, G.; Yang, X.; Mo, X.; Zhao, S.; Calonje, M.; et al. H2AK121ub in Arabidopsis Associates with a Less Accessible Chromatin State at Transcriptional Regulation Hotspots. Nat. Commun. 2021, 12, 315. [Google Scholar] [CrossRef] [PubMed]
- Freitag, M. Histone Methylation by SET Domain Proteins in Fungi. Annu. Rev. Microbiol. 2017, 71, 413–439. [Google Scholar] [CrossRef]
- Köhler, C.; Villar, C.B.R. Programming of Gene Expression by Polycomb Group Proteins. Trends Cell Biol. 2008, 18, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Trojer, P.; Reinberg, D. Histone Lysine Demethylases and Their Impact on Epigenetics. Cell 2006, 125, 213–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharaf, A.; Vijayanathan, M.; Oborník, M.; Mozgová, I. Phylogenetic Profiling Suggests Early Origin of the Core Subunits of Polycomb Repressive Complex 2 (PRC2). 2021. Available online: https://www.biorxiv.org/content/10.1101/2021.07.16.452543v1.abstract (accessed on 28 December 2021).
- Margulis, L.; Chapman, M.; Guerrero, R.; Hall, J. The Last Eukaryotic Common Ancestor (LECA): Acquisition of Cytoskeletal Motility from Aerotolerant Spirochetes in the Proterozoic Eon. Proc. Natl. Acad. Sci. USA 2006, 103, 13080. [Google Scholar] [CrossRef] [Green Version]
- O’Malley, M.A.; Leger, M.M.; Wideman, J.G.; Ruiz-Trillo, I. Concepts of the Last Eukaryotic Common Ancestor. Nat. Ecol. Evol. 2019, 3, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Spillane, C.; Schmid, K.J.; Laoueillé-Duprat, S.; Pien, S.; Escobar-Restrepo, J.M.; Baroux, C.; Gagliardini, V.; Page, D.R.; Wolfe, K.H.; Grossniklaus, U. Positive Darwinian Selection at the Imprinted MEDEA Locus in Plants. Nature 2007, 448, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Liu, S.L.; Adams, K.L. Concerted Divergence after Gene Duplication in Polycomb Repressive Complexes. Plant Physiol. 2017, 174, 1192–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.T.Y. Architectural Organization of Dinoflagellate Liquid Crystalline Chromosomes. Microorganisms 2019, 7, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marinov, G.K.; Lynch, M. Diversity and Divergence of Dinoflagellate Histone Proteins. G3: Genes Genomes Genet. 2016, 6, 397–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gornik, S.G.; Ford, K.L.; Mulhern, T.D.; Bacic, A.; McFadden, G.I.; Waller, R.F. Loss of Nucleosomal DNA Condensation Coincides with Appearance of a Novel Nuclear Protein in Dinoflagellates. Curr. Biol. 2012, 22, 2303–2312. [Google Scholar] [CrossRef] [Green Version]
- Pan, B.; Chen, X.; Hou, L.; Zhang, Q.; Qu, Z.; Warren, A.; Miao, M. Comparative Genomics Analysis of Ciliates Provides Insights on the Evolutionary History Within “Nassophorea–Synhymenia–Phyllopharyngea” Assemblage. Front. Microbiol. 2019, 10, 2819. [Google Scholar] [CrossRef] [Green Version]
- Chalker, D.L.; Meyer, E.; Mochizuki, K. Epigenetics of Ciliates. Cold Spring Harb. Perspect. Biol. 2013, 5, a017764. [Google Scholar] [CrossRef] [Green Version]
- Taverna, S.D.; Coyne, R.S.; Allis, C.D. Methylation of Histone H3 at Lysine 9 Targets Programmed DNA Elimination in Tetrahymena. Cell 2002, 110, 701–711. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Mochizuki, K.; Gorovsky, M.A. Histone H3 Lysine 9 Methylation Is Required for DNA Elimination in Developing Macronuclei in Tetrahymena. Proc. Natl. Acad. Sci. USA 2004, 101, 1679–1684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Taverna, S.D.; Muratore, T.L.; Shabanowitz, J.; Hunt, D.F.; Allis, C.D. RNAi-Dependent H3K27 Methylation Is Required for Heterochromatin Formation and DNA Elimination in Tetrahymena. Genes Dev. 2007, 21, 1530–1545. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Xiong, J.; Mao, F.; Sheng, Y.; Chen, X.; Feng, L.; Dui, W.; Yang, W.; Kapusta, A.; Feschotte, C.; et al. RNAi-Dependent Polycomb Repression Controls Transposable Elements in Tetrahymena. Genes Dev. 2019, 33, 348–364. [Google Scholar] [CrossRef] [Green Version]
- Cao, R.; Wang, L.; Wang, H.; Xia, L.; Erdjument-Bromage, H.; Tempst, P.; Jones, R.S.; Zhang, Y. Role of Histone H3 Lysine 27 Methylation in Polycomb-Group Silencing. Science 2002, 298, 1039–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czermin, B.; Melfi, R.; McCabe, D.; Seitz, V.; Imhof, A.; Pirrotta, V. Drosophila Enhancer of Zeste/ESC Complexes Have a Histone H3 Methyltransferase Activity That Marks Chromosomal Polycomb Sites. Cell 2002, 111, 185–196. [Google Scholar] [CrossRef] [Green Version]
- Kuzmichev, A.; Nishioka, K.; Erdjument-Bromage, H.; Tempst, P.; Reinberg, D. Histone Methyltransferase Activity Associated with a Human Multiprotein Complex Containing the Enhancer of Zeste Protein. Genes Dev. 2002, 16, 2893–2905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erlendson, A.A.; Friedman, S.; Freitag, M. A Matter of Scale and Dimensions: Chromatin of Chromosome Landmarks in the Fungi. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.M.; Kothe, G.O.; Matsen, C.B.; Khlafallah, T.K.; Adhvaryu, K.K.; Hemphill, M.; Freitag, M.; Motamedi, M.R.; Selker, E.U. The Fungus Neurospora Crassa Displays Telomeric Silencing Mediated by Multiple Sirtuins and by Methylation of Histone H3 Lysine 9. Epigenet. Chromatin 2008, 1, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bratkowski, M.; Yang, X.; Liu, X. An Evolutionarily Conserved Structural Platform for PRC2 Inhibition by a Class of Ezh2 Inhibitors. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Studt, L.; Rösler, S.M.; Burkhardt, I.; Arndt, B.; Freitag, M.; Humpf, H.U.; Dickschat, J.S.; Tudzynski, B. Knock-down of the Methyltransferase Kmt6 Relieves H3K27me3 and Results in Induction of Cryptic and Otherwise Silent Secondary Metabolite Gene Clusters in Fusarium Fujikuroi. Environ. Microbiol. 2016, 18, 4037–4054. [Google Scholar] [CrossRef] [Green Version]
- Carlier, F.; Li, M.; Maroc, L.; Debuchy, R.; Souaid, C.; Noordermeer, D.; Grognet, P.; Malagnac, F. Loss of EZH2-like or SU(VAR)3–9-like Proteins Causes Simultaneous Perturbations in H3K27 and H3K9 Tri-Methylation and Associated Developmental Defects in the Fungus Podospora Anserina. Epigenet. Chromatin 2021, 14, 22. [Google Scholar] [CrossRef]
- Ferraro, A.R.; Ameri, A.J.; Lu, Z.; Kamei, M.; Schmitz, R.J.; Lewis, Z.A. Chromatin Accessibility Profiling in Neurospora Crassa Reveals Molecular Features Associated with Accessible and Inaccessible Chromatin. BMC Genom. 2021, 22, 459. [Google Scholar] [CrossRef]
- Jiao, L.; Liu, X. Structural Analysis of an Active Fungal PRC2. Nucleus 2016, 7, 284–291. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E.A.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon Queenslandica Genome and the Evolution of Animal Complexity. Nature 2010, 466, 720–726. [Google Scholar] [CrossRef]
- Müller, W.E.G.; Wiens, M.; Adell, T.; Gamulin, V.; Schröder, H.C.; Müller, I.M. Bauplan of Urmetazoa: Basis for Genetic Complexity of Metazoa. Int. Rev. Cytol. 2004, 235, 53–92. [Google Scholar] [CrossRef]
- Verheul, T.C.J.; van Hijfte, L.; Perenthaler, E.; Barakat, T.S. The Why of YY1: Mechanisms of Transcriptional Regulation by Yin Yang 1. Front. Cell Dev. Biol. 2020, 8, 1034. [Google Scholar] [CrossRef]
- Vandamme, J.; Sidoli, S.; Mariani, L.; Friis, C.; Christensen, J.; Helin, K.; Jensen, O.N.; Salcini, A.E. H3K23me2 Is a New Heterochromatic Mark in Caenorhabditis Elegans. Nucleic Acids Res. 2015, 43, 9694–9710. [Google Scholar] [CrossRef] [Green Version]
- Ahringer, J.; Gasser, S.M. Repressive Chromatin in Caenorhabditis Elegans: Establishment, Composition, and Function. Genetics 2018, 208, 491–511. [Google Scholar] [CrossRef] [Green Version]
- Holdeman, R.; Nehrt, S.; Strome, S. MES-2, a Maternal Protein Essential for Viability of the Germline in Caenorhabditis Elegans, Is Homologous to a Drosophila Polycomb Group Protein. Development 1998, 125, 2457–2467. [Google Scholar] [CrossRef]
- Xu, L.; Paulsen, J.; Yoo, Y.; Goodwin, E.B.; Strome, S. Caenorhabditis Elegans MES-3 Is a Target of GLD-1 and Functions Epigenetically in Germline Development. Genetics 2001, 159, 1007–1017. [Google Scholar] [CrossRef]
- Guillermo, A.R.R.; Chocian, K.; Gavriilidis, G.; Vandamme, J.; Salcini, A.E.; Mellor, J.; Woollard, A. H3K27 Modifiers Regulate Lifespan in C. Elegans in a Context-Dependent Manner. BMC Biol. 2021, 19, 59. [Google Scholar] [CrossRef]
- Matsuzaki, M.; Misumi, O.; Shin-I, T.; Maruyama, S.; Takahara, M.; Miyagishima, S.Y.; Mori, T.; Nishida, K.; Yagisawa, F.; Nishida, K.; et al. Genome Sequence of the Ultrasmall Unicellular Red Alga Cyanidioschyzon Merolae 10D. Nature 2004, 428, 653–657. [Google Scholar] [CrossRef]
- Schubert, D. Evolution of Polycomb-Group Function in the Green Lineage. F1000Research 2019, 8, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, J.; Wagner, D. Polycomb Repression in the Regulation of Growth and Development in Arabidopsis. Curr. Opin. Plant Biol. 2015, 23, 15–24. [Google Scholar] [CrossRef]
- Mozgova, I.; Köhler, C.; Hennig, L. Keeping the Gate Closed: Functions of the Polycomb Repressive Complex PRC2 in Development. Plant J. 2015, 83, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Lv, Y.; Zhao, C.; Wang, X. Knowing When to Silence: Roles of Polycomb-Group Proteins in SAM Maintenance, Root Development, and Developmental Phase Transition. Int. J. Mol. Sci. 2020, 21, 5871. [Google Scholar] [CrossRef]
- Tonosaki, K.; Ono, A.; Kunisada, M.; Nishino, M.; Nagata, H.; Sakamoto, S.; Kijima, S.T.; Furuumi, H.; Nonomura, K.I.; Sato, Y.; et al. Mutation of the Imprinted Gene OsEMF2a Induces Autonomous Endosperm Development and Delayed Cellularization in Rice. Plant Cell 2021, 33, 85–103. [Google Scholar] [CrossRef]
- Springer, N.M.; Danilevskaya, O.N.; Hermon, P.; Helentjaris, T.G.; Phillips, R.L.; Kaeppler, H.F.; Kaeppler, S.M. Sequence Relationships, Conserved Domains, and Expression Patterns for Maize Homologs of the Polycomb Group Genes E(z), Esc, and E(Pc). Plant Physiol. 2002, 128, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Horst, N.A.; Katz, A.; Pereman, I.; Decker, E.L.; Ohad, N.; Reski, R. A Single Homeobox Gene Triggers Phase Transition, Embryogenesis and Asexual Reproduction. Nat. Plants 2016, 2, 15209. [Google Scholar] [CrossRef] [PubMed]
- Widiez, T.; Symeonidi, A.; Luo, C.; Lam, E.; Lawton, M.; Rensing, S.A. The Chromatin Landscape of the Moss Physcomitrella Patens and Its Dynamics during Development and Drought Stress. Plant J. 2014, 79, 67–81. [Google Scholar] [CrossRef]
- Goodrich, J.; Puangsomlee, P.; Martin, M.; Long, D.; Meyerowitz, E.M.; Coupland, G. A Polycomb-Group Gene Regulates Homeotic Gene Expression in Arabidopsis. Nature 1997, 386, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Grossniklaus, U.; Vielle-Calzada, J.-P.; Hoeppner, M.A.; Gagliano, W.B. Maternal Control of Embryogenesis by MEDEA, a Polycomb Group Gene in Arabidopsis. Science 1998, 280, 446–450. [Google Scholar] [CrossRef]
- Danilevskaya, O.N.; Hermon, P.; Hantke, S.; Muszynski, M.G.; Kollipara, K.; Ananiev, E.V. Duplicated Fie Genes in Maize: Expression Pattern and Imprinting Suggest Distinct Functions. Plant Cell 2003, 15, 425–438. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, C.; Cheng, J.; Liu, J.; Zhang, L.; He, C.; Shen, W.H.; Jin, H.; Xu, L.; Zhang, Y. Arabidopsis Flower and Embryo Developmental Genes Are Repressed in Seedlings by Different Combinations of Polycomb Group Proteins in Association with Distinct Sets of Cis-Regulatory Elements. PLoS Genet. 2016, 12, e1005771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereman, I.; Mosquna, A.; Katz, A.; Wiedemann, G.; Lang, D.; Decker, E.L.; Tamada, Y.; Ishikawa, T.; Nishiyama, T.; Hasebe, M.; et al. The Polycomb Group Protein CLF Emerges as a Specific Tri-Methylase of H3K27 Regulating Gene Expression and Development in Physcomitrella Patens. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2016, 1859, 860–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, D.J.; Tedds, H.M.; Labandera, A.M.; Bailey, M.; White, M.D.; Hartman, S.; Sprigg, C.; Mogg, S.L.; Osborne, R.; Dambire, C.; et al. Oxygen-Dependent Proteolysis Regulates the Stability of Angiosperm Polycomb Repressive Complex 2 Subunit VERNALIZATION 2. Nat. Commun. 2018, 9, 5438. [Google Scholar] [CrossRef]
- Katz, A.; Oliva, M.; Mosquna, A.; Hakim, O.; Ohad, N. FIE and CURLY LEAF Polycomb Proteins Interact in the Regulation of Homeobox Gene Expression during Sporophyte Development. Plant J. 2004, 37, 707–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, G.; Zhu, X.; Elling, A.A.; Chen, L.; Wang, X.; Guo, L.; Liang, M.; He, H.; Zhang, H.; Chen, F.; et al. Global Epigenetic and Transcriptional Trends among Two Rice Subspecies and Their Reciprocal Hybrids. Plant Cell 2010, 22, 17–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saripalli, G.; Singh, K.; Gautam, T.; Kumar, S.; Raghuvanshi, S.; Prasad, P.; Jain, N.; Sharma, P.K.; Balyan, H.S.; Gupta, P.K. Genome-Wide Analysis of H3K4me3 and H3K27me3 Modifications Due to Lr28 for Leaf Rust Resistance in Bread Wheat (Triticum Aestivum). Plant Mol. Biol. 2020, 104, 113–136. [Google Scholar] [CrossRef] [PubMed]
- Makarevitch, I.; Eichten, S.R.; Briskine, R.; Waters, A.J.; Danilevskaya, O.N.; Meeley, R.B.; Myers, C.L.; Vaughn, M.W.; Springer, N.M. Genomic Distribution of Maize Facultative Heterochromatin Marked by Trimethylation of H3K27. Plant Cell 2013, 25, 780–793. [Google Scholar] [CrossRef] [Green Version]
- Baker, K.; Dhillon, T.; Colas, I.; Cook, N.; Milne, I.; Milne, L.; Bayer, M.; Flavell, A.J. Chromatin State Analysis of the Barley Epigenome Reveals a Higher-Order Structure Defined by H3K27me1 and H3K27me3 Abundance. Plant J. 2015, 84, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Payá-Milans, M.; Poza-Viejo, L.; Martín-Uriz, P.S.; Lara-Astiaso, D.; Wilkinson, M.D.; Crevillén, P. Genome-Wide Analysis of the H3K27me3 Epigenome and Transcriptome in Brassica Rapa. GigaScience 2019, 8, giz147. [Google Scholar] [CrossRef] [Green Version]
- Huan, Q.; Mao, Z.; Chong, K.; Zhang, J. Global Analysis of H3K4me3/H3K27me3 in Brachypodium Distachyon Reveals VRN3 as Critical Epigenetic Regulation Point in Vernalization and Provides Insights into Epigenetic Memory. New Phytol. 2018, 219, 1373–1387. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Elling, A.A.; Li, X.; Li, N.; Peng, Z.; He, G.; Sun, H.; Qi, Y.; Liu, X.S.; Deng, X.W. Genome-Wide and Organ-Specific Landscapes of Epigenetic Modifications and Their Relationships to MRNA and Small RNA Transcriptomes in Maize. Plant Cell 2009, 21, 1053–1069. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, J.; Jovtchev, G.; Schubert, I. The Chromosomal Distribution of Histone Methylation Marks in Gymnosperms Differs from That of Angiosperms. Chromosome Res. 2008, 16, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Trejo-arellano, M.S.; Qiu, Y.; Eklund, D.M.; Köhler, C.; Hennig, L. H2A Ubiquitination is Essential for Polycomb Repressive Complex 1-Mediated Gene Regulation in Marchantia polymorpha. Genome Biol. 2021, 22, 253. [Google Scholar] [CrossRef]
- Lang, D.; Ullrich, K.K.; Murat, F.; Fuchs, J.; Jenkins, J.; Haas, F.B.; Piednoel, M.; Gundlach, H.; Van Bel, M.; Meyberg, R.; et al. The Physcomitrella Patens Chromosome-Scale Assembly Reveals Moss Genome Structure and Evolution. Plant J. 2018, 93, 515–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterborg, J.H.; Robertson, A.J.; Tatar, D.L.; Borza, C.M.; Davie, J.R. Histones of Chlamydomonas Reinhardtii (Synthesis, Acetylation, and Methylation). Plant Physiol. 1995, 109, 393–407. [Google Scholar] [CrossRef] [Green Version]
- Ngan, C.Y.; Wong, C.-H.; Choi, C.; Yoshinaga, Y.; Louie, K.; Jia, J.; Chen, C.; Bowen, B.; Cheng, H.; Leonelli, L.; et al. Lineage-Specific Chromatin Signatures Reveal a Regulator of Lipid Metabolism in Microalgae. Nat. Plants 2015, 1, 15107. [Google Scholar] [CrossRef] [PubMed]
- Kiyosue, T.; Ohad, N.; Yadegari, R.; Hannon, M.; Dinneny, J.; Wells, D.; Katz, A.; Margossian, L.; Harada, J.J.; Goldberg, R.B.; et al. Control of Fertilization-Independent Endosperm Development by the MEDEA Polycomb Gene in Arabidopsis. Proc. Natl. Acad. Sci. USA 1999, 96, 4186. [Google Scholar] [CrossRef] [Green Version]
- Ohad, N.; Yadegari, R.; Margossian, L.; Hannon, M.; Michaeli, D.; Harada, J.J.; Goldberg, R.B.; Fischer, R.L. Mutations in FIE, a WD Polycomb Group Gene, Allow Endosperm Development without Fertilization. Plant Cell 1999, 11, 407–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, M.; Bilodeau, P.; Koltunow, A.; Dennis, E.S.; Peacock, W.J.; Chaudhury, A.M. Genes Controlling Fertilization-Independent Seed Development in Arabidopsis Thaliana. Proc. Natl. Acad. Sci. USA 1999, 96, 296–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringrose, L.; Ehret, H.; Paro, R. Distinct Contributions of Histone H3 Lysine 9 and 27 Methylation to Locus-Specific Stability of Polycomb Complexes. Mol. Cell 2004, 16, 641–653. [Google Scholar] [CrossRef]
- Kim, J.; Kim, H. Recruitment and Biological Consequences of Histone Modification of H3K27me3 and H3K9me3. ILAR J. 2012, 53, 232–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zervudacki, J.; Yu, A.; Amesefe, D.; Wang, J.; Drouaud, J.; Navarro, L.; Deleris, A. Transcriptional Control and Exploitation of an Immune-Responsive Family of Plant Retrotransposons. EMBO J. 2018, 37, e98482. [Google Scholar] [CrossRef]
- Peters, A.H.F.M.; Kubicek, S.; Mechtler, K.; O’Sullivan, R.J.; Derijck, A.A.H.A.; Perez-Burgos, L.; Kohlmaier, A.; Opravil, S.; Tachibana, M.; Shinkai, Y.; et al. Partitioning and Plasticity of Repressive Histone Methylation States in Mammalian Chromatin. Mol. Cell 2003, 12, 1577–1589. [Google Scholar] [CrossRef]
- Saksouk, N.; Barth, T.K.; Ziegler-Birling, C.; Olova, N.; Nowak, A.; Rey, E.; Mateos-Langerak, J.; Urbach, S.; Reik, W.; Torres-Padilla, M.-E.; et al. Redundant Mechanisms to Form Silent Chromatin at Pericentromeric Regions Rely on BEND3 and DNA Methylation. Mol. Cell 2014, 56, 580–594. [Google Scholar] [CrossRef] [Green Version]
- Walter, M.; Teissandier, A.; Pérez-Palacios, R.; Bourc’his, D. An Epigenetic Switch Ensures Transposon Repression upon Dynamic Loss of DNA Methylation in Embryonic Stem Cells. eLife 2016, 5, e11418. [Google Scholar] [CrossRef]
- Deleris, A.; Stroud, H.; Bernatavichute, Y.; Johnson, E.; Klein, G.; Schubert, D.; Jacobsen, S.E. Loss of the DNA Methyltransferase MET1 Induces H3K9 Hypermethylation at PcG Target Genes and Redistribution of H3K27 Trimethylation to Transposons in Arabidopsis Thaliana. PLoS Genet. 2012, 8, e1003062. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, O.; Probst, A.V.; Paszkowski, J. Distinct Regulation of Histone H3 Methylation at Lysines 27 and 9 by CpG Methylation in Arabidopsis. EMBO J. 2005, 24, 2783–2791. [Google Scholar] [CrossRef] [Green Version]
- Weinhofer, I.; Hehenberger, E.; Roszak, P.; Hennig, L.; Köhler, C. H3K27me3 Profiling of the Endosperm Implies Exclusion of Polycomb Group Protein Targeting by DNA Methylation. PLoS Genet. 2010, 6, e1001152. [Google Scholar] [CrossRef]
- Moreno-Romero, J.; Del Toro-De León, G.; Yadav, V.K.; Santos-González, J.; Köhler, C. Epigenetic Signatures Associated with Imprinted Paternally Expressed Genes in the Arabidopsis Endosperm. Genome Biol. 2019, 20, 41. [Google Scholar] [CrossRef] [Green Version]
- Rougée, M.; Quadrana, L.; Zervudacki, J.; Hure, V.; Colot, V.; Navarro, L.; Deleris, A. Polycomb Mutant Partially Suppresses DNA Hypomethylation-Associated Phenotypes in Arabidopsis. Life Sci. Alliance 2021, 4, e202000848. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, K.; Wiles, E.T.; McNaught, K.J.; Sidoli, S.; Leggett, N.; Shao, Y.; Garcia, B.A.; Selker, E.U. Loss of HP1 Causes Depletion of H3K27me3 from Facultative Heterochromatin and Gain of H3K27me2 at Constitutive Heterochromatin. Genome Res. 2016, 26, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Basenko, E.Y.; Sasaki, T.; Ji, L.; Prybol, C.J.; Burckhardt, R.M.; Schmitz, R.J.; Lewis, Z.A. Genome-Wide Redistribution of H3K27me3 Is Linked to Genotoxic Stress and Defective Growth. Proc. Natl. Acad. Sci. USA 2015, 112, E6339–E6348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möller, M.; Schotanus, K.; Soyer, J.L.; Haueisen, J.; Happ, K.; Stralucke, M.; Happel, P.; Smith, K.M.; Connolly, L.R.; Freitag, M.; et al. Destabilization of Chromosome Structure by Histone H3 Lysine 27 Methylation. PLoS Genet. 2019, 15, e1008093. [Google Scholar] [CrossRef] [Green Version]
- Leeb, M.; Pasini, D.; Novatchkova, M.; Jaritz, M.; Helin, K.; Wutz, A. Polycomb Complexes Act Redundantly to Repress Genomic Repeats and Genes. Genes Dev. 2010, 24, 265–276. [Google Scholar] [CrossRef] [Green Version]
- Klocko, A.D.; Ormsby, T.; Galazka, J.M.; Leggett, N.A.; Uesaka, M.; Honda, S.; Freitag, M.; Selker, E.U. Normal Chromosome Conformation Depends on Subtelomeric Facultative Heterochromatin in Neurospora Crassa. Proc. Natl. Acad. Sci. USA 2016, 113, 15048–15053. [Google Scholar] [CrossRef] [Green Version]
- Montero, J.J.; López-Silanes, I.; Megías, D.; Fraga, F.M.; Castells-García, Á.; Blasco, M.A. TERRA Recruitment of Polycomb to Telomeres Is Essential for Histone Trymethylation Marks at Telomeric Heterochromatin. Nat. Commun. 2018, 9, 1548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majerová, E.; Mandáková, T.; Vu, G.T.H.; Fajkus, J.; Lysak, M.A.; Fojtová, M. Chromatin Features of Plant Telomeric Sequences at Terminal vs. Internal Positions. Front. Plant Sci. 2014, 5, 593. [Google Scholar] [CrossRef] [Green Version]
- Adamusová, K.; Khosravi, S.; Fujimoto, S.; Houben, A.; Matsunaga, S.; Fajkus, J.; Fojtová, M. Two Combinatorial Patterns of Telomere Histone Marks in Plants with Canonical and Non-Canonical Telomere Repeats. Plant J. 2020, 102, 678–687. [Google Scholar] [CrossRef] [PubMed]
- Bettin, N.; Oss Pegorar, C.; Cusanelli, E. The Emerging Roles of TERRA in Telomere Maintenance and Genome Stability. Cells 2019, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Vrbsky, J.; Akimcheva, S.; Watson, J.M.; Turner, T.L.; Daxinger, L.; Vyskot, B.; Aufsatz, W.; Riha, K. SiRNA–Mediated Methylation of Arabidopsis Telomeres. PLoS Genet. 2010, 6, e1000986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lange, T. Shelterin: The Protein Complex That Shapes and Safeguards Human Telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [Green Version]
- Marión, R.M.; Montero, J.J.; de Silanes, I.L.; Graña-Castro, O.; Martínez, P.; Schoeftner, S.; Palacios-Fábrega, J.A.; Blasco, M.A. TERRA Regulate the Transcriptional Landscape of Pluripotent Cells through TRF1-Dependent Recruitment of PRC2. eLife 2019, 8, e44656. [Google Scholar] [CrossRef] [PubMed]
- Procházková Schrumpfová, P.; Vychodilová, I.; Dvořáčková, M.; Majerská, J.; Dokládal, L.; Schořová, Š.; Fajkus, J. Telomere Repeat Binding Proteins Are Functional Components of Arabidopsis Telomeres and Interact with Telomerase. Plant J. 2014, 77, 770–781. [Google Scholar] [CrossRef] [Green Version]
- Procházková Schrumpfová, P.; Fojtová, M.; Fajkus, J. Telomeres in Plants and Humans: Not So Different, Not So Similar. Cells 2019, 8, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Hartwig, B.; James, G.V.; Schneeberger, K.; Turck, F. Complementary Activities of TELOMERE REPEAT BINDING Proteins and Polycomb Group Complexes in Transcriptional Regulation of Target Genes. Plant Cell 2016, 28, 87–101. [Google Scholar] [CrossRef]
- Jamieson, K.; McNaught, K.J.; Ormsby, T.; Leggett, N.A.; Honda, S.; Selker, E.U. Telomere Repeats Induce Domains of H3K27 Methylation in Neurospora. eLife 2018, 7, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Teano, G.; Concia, L.; Carron, L.; Wolff, L.; Adamusová, K.; Fojtová, M.; Bourge, M.; Kramdi, A.; Colot, V.; Grossniklaus, U.; et al. Histone H1 Protects Telomeric Repeats from H3K27me3 Invasion in Arabidopsis. bioRxiv 2021. [Google Scholar] [CrossRef]
- Marasca, F.; Bodega, B.; Orlando, V. How Polycomb-Mediated Cell Memory Deals With a Changing Environment. BioEssays 2018, 40, 1700137. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Supergroups | Eukaryotic Group/Kingdom | Phylum or Class | Species | E(z) | Su(z)12 | Esc | p55 | References |
|---|---|---|---|---|---|---|---|---|
| Archaeplastida | Eukaryota | Rhodophyta | Cyanidioschyzon merolae | ✓ | ✓ | ✓ | ✓ (2) | [63] |
| Viridiplantae | Chlorophyta | Ostreococcus lucimarinus | ✓ | ✓ | ✓ | ✓(2) | [89] | |
| Chlamydomonas reinhardtii | ✓ | ✓ | ✓(2) | ✓(2) | [63,89] | |||
| Volvox carteri | ✓(2) | ? | ✓ | ✓(2) | [89] | |||
| Viridiplantae—Embryophyta | Bryophyta-Bryopsida | Physcomitrium patens | ✓ | ✓(3) | ✓ | ✓(2) | [61,62,89,90] | |
| Lycophyte-Lycopodiopsida | Selaginella moellendorffii | ✓(2) | ✓ | ✓ | ✓(3) | [89,90,91] | ||
| Gymnosperm | Picea abies | ✓ | ? | ? | ? | [92] | ||
| Angiosperms-Monocot | Oryza sativa | ✓(2) | ✓(2) | ✓(2) | ✓(2) | [91,93,94,95,96] | ||
| Triticum aestivum | ✓(9) | ✓(8) | ✓ (7) | ✓(6) | [97] | |||
| Zea mays | ✓(3) | ✓(2) | ✓(2) | ✓(5) | [94] | |||
| Brachypodium distachyon | ✓(2) | ✓(2) | ✓(3) | ✓(4) | [89,98,99] | |||
| Hordeum vulgare | ✓(3) | ✓(3) | ✓ | ✓(2) | [94,100,101] | |||
| Sorghum bicolor | ✓(2) | ✓(3) | ✓(2) | ✓(2) | [94] | |||
| Angiosperms-Eudicot | Arabidopsis thaliana | ✓(3) | ✓(3) | ✓ | ✓(5) | [102] | ||
| Chromalveolata | SAR—Stramenopiles | Bacillariophyceae (diatoms) | Phaeodactylum tricornutum | ✓ | ✓ | ✓ | ✓ | [89,103] |
| Ochrophyta-Phaeophyceae | Ectocarpus | - | - | - | - | [104] | ||
| SAR—Alveolata | Cilliophora/cilliates | Paramecium tetraurelia | ✓(2) | - | - | ✓ | [31,105,106] | |
| Tetrahymena thermophila | ✓ | ✓ | ✓ | ✓ | [107,108] | |||
| Opisthokonta | Fungi | Basidiomycota | Cryptococcus neoformans | ✓ | - | ✓ | ✓ | [64,109] |
| Ascomycota | Fusarium graminearum | ✓ | ✓ | ✓ | ✓ | [109,110] | ||
| Chaetomium thermophilum | ✓ | ✓ | ✓ | ✓ | [79] | |||
| Neurospora crassa | ✓ | ✓ | ✓ | ✓ | [109,111,112] | |||
| Saccharomyces cerevisiae | - | - | - | - | [31,75] | |||
| Filasterea—single-celled eukaryote | Capsaspora | Capsaspora owczarzaki | - | - | - | - | [113] | |
| Animalia/animals | Porifera | Amphimedon queenslandica | ✓(4) | ✓ | ✓(2) | ✓ | [114] | |
| Cnidaria/Hydrozoa | Hydra vulgaris | ✓ | ✓ | ✓ | ✓ | [115,116,117,118] | ||
| Insecta/insects | Drosophila melanogaster | ✓ | ✓ | ✓ | ✓ | [119,120,121] | ||
| Nematoda/nematodes | Caenorhabditis elegans | ✓ | - | ✓ | - | [122,123] | ||
| Reptilia/reptiles | Anolis carolinensis | ✓ | ✓ | ✓ | ✓ | [124,125,126,127] | ||
| Mammalia/mammals | Homo sapiens | ✓(2) | ✓ | ✓ | ✓ | [56,60] | ||
| Mus musculus | ✓(2) | ✓ | ✓ | ✓ | [128,129] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vijayanathan, M.; Trejo-Arellano, M.G.; Mozgová, I. Polycomb Repressive Complex 2 in Eukaryotes—An Evolutionary Perspective. Epigenomes 2022, 6, 3. https://doi.org/10.3390/epigenomes6010003
Vijayanathan M, Trejo-Arellano MG, Mozgová I. Polycomb Repressive Complex 2 in Eukaryotes—An Evolutionary Perspective. Epigenomes. 2022; 6(1):3. https://doi.org/10.3390/epigenomes6010003
Chicago/Turabian StyleVijayanathan, Mallika, María Guadalupe Trejo-Arellano, and Iva Mozgová. 2022. "Polycomb Repressive Complex 2 in Eukaryotes—An Evolutionary Perspective" Epigenomes 6, no. 1: 3. https://doi.org/10.3390/epigenomes6010003