
Use of the Proboscis Extension Response Assay to Evaluate the Mechanism of House Fly Behavioral Resistance to Imidacloprid

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. House Fly Strains
2.2. Proboscis Extension Response—Proboscis Contact
2.3. Proboscis Extension Response—Tarsal Contact
2.4. Statistical Analysis
3. Results
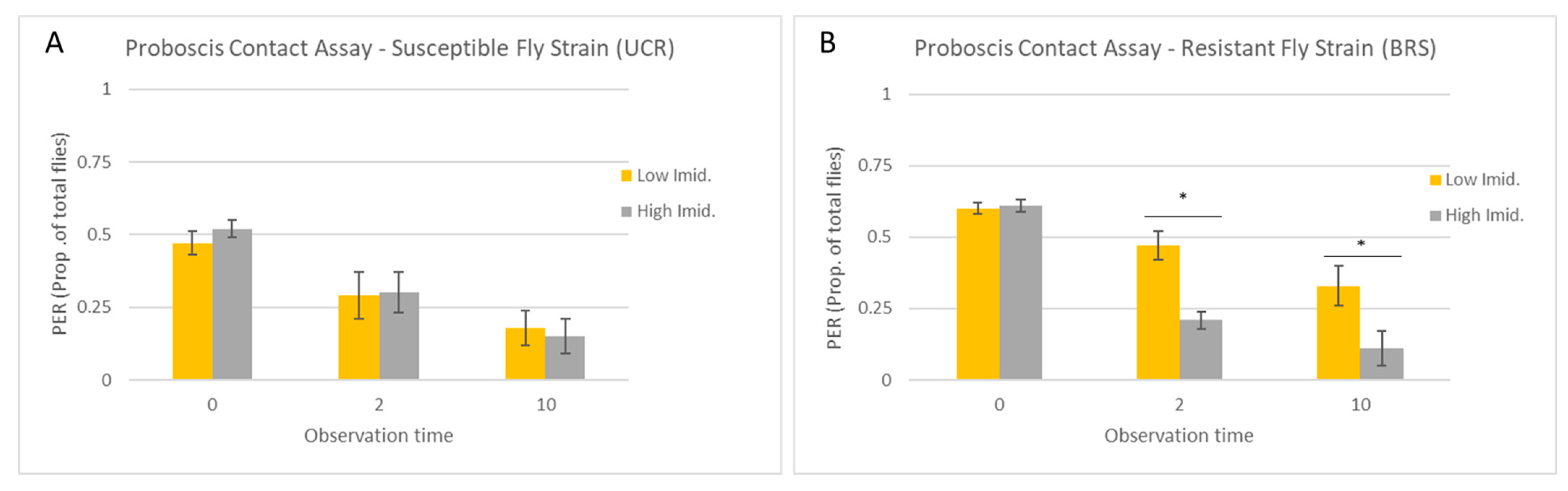
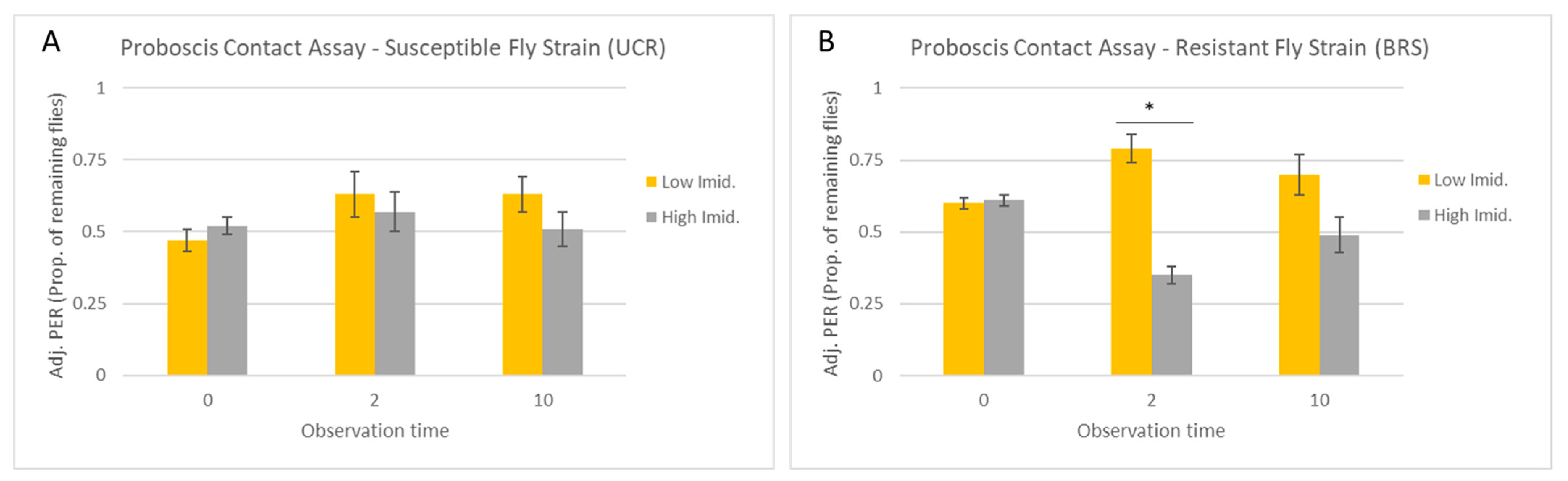
3.1. Proboscis Contact Assay
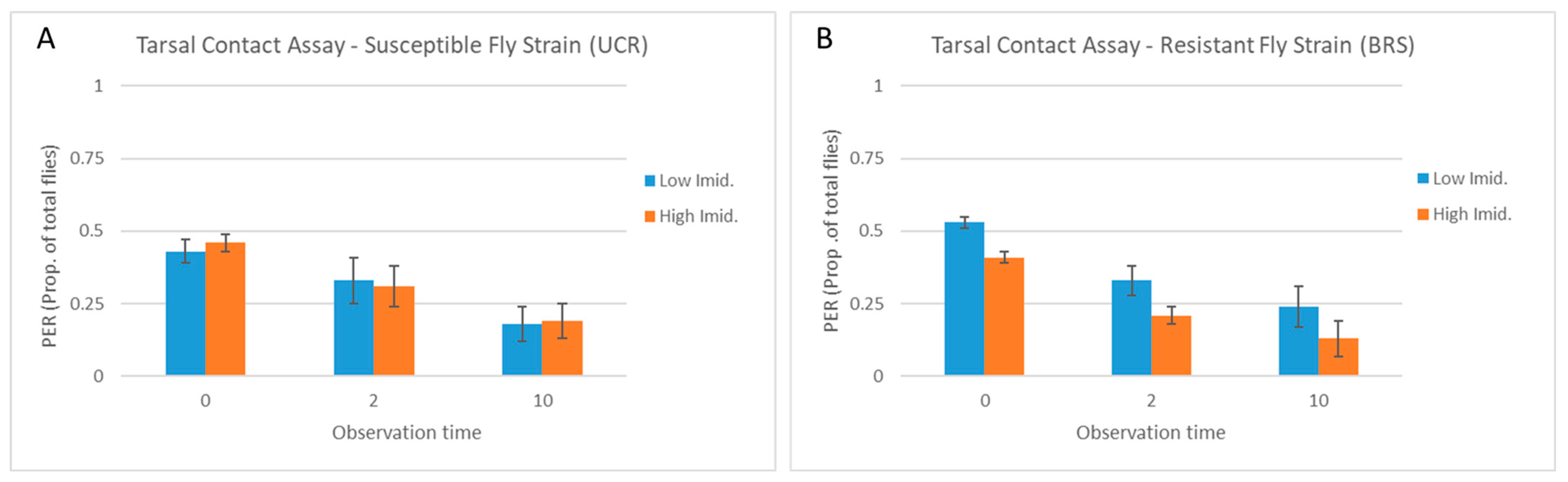
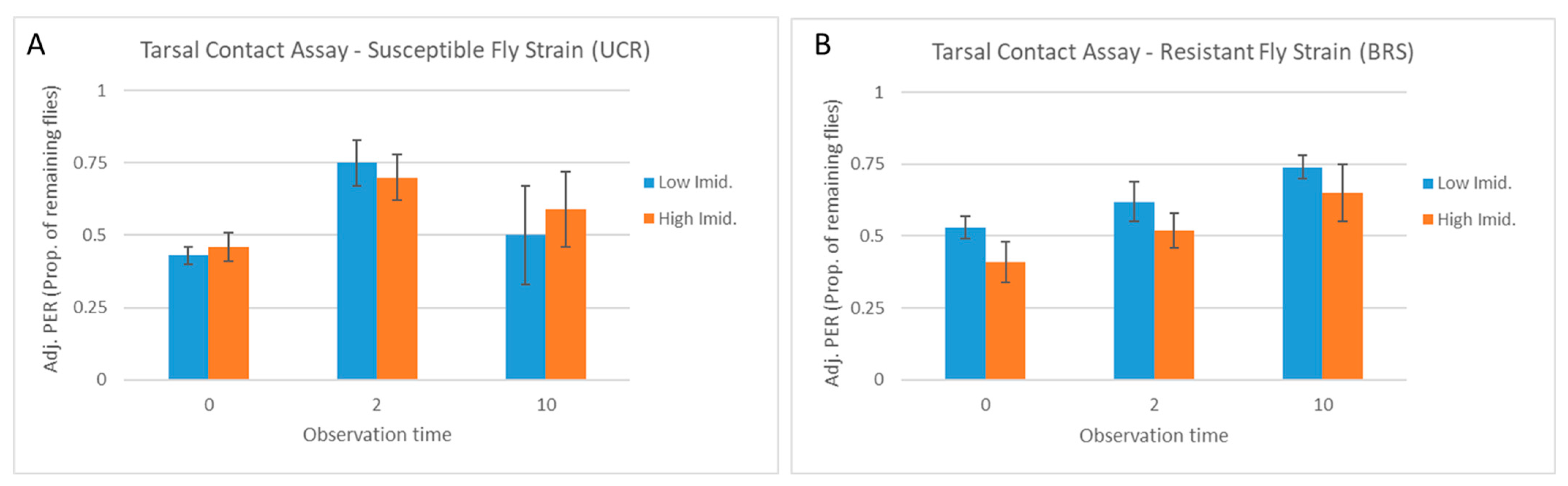
3.2. Tarsal Contact Assay
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Greenberg, B. Flies and Disease: I. Ecology, Classification, and Biotic Associations; Princeton University Press: Princeton, NJ, USA, 2019. [Google Scholar] [CrossRef]
- Nayduch, D.; Neupane, S.; Pickens, V.; Purvis, T.; Olds, C. House Flies Are Underappreciated Yet Important Reservoirs and Vectors of Microbial Threats to Animal and Human Health. Microorganisms 2023, 11, 583. [Google Scholar] [CrossRef]
- Geden, C.J.; Nayduch, D.; Scott, J.G.; Burgess, E.R.; Gerry, A.C.; Kaufman, P.E.; Thomson, J.; Pickens, V.; Machtinger, E.T. House Fly (Diptera: Muscidae): Biology, Pest Status, Current Management Prospects, and Research Needs. J. Integr. Pest Manag. 2021, 12, 39. [Google Scholar] [CrossRef]
- Butler, S.M.; Gerry, A.C.; Mullens, B.A. House Fly (Diptera: Muscidae) Activity near Baits Containing (Z)-9-Tricosene and Efficacy of Commercial Toxic Fly Baits on a Southern California Dairy. J. Econ. Entomol. 2007, 100, 1489–1495. [Google Scholar] [CrossRef]
- Gerry, A.C.; Zhang, D. Behavioral Resistance of House Flies, Musca domestica (Diptera: Muscidae) to Imidacloprid. US Army Med. Dep. J. 2009, 54–59. [Google Scholar]
- Tan, M.I.; Yusmalinar, S.; Ambarningr, T.B.; Ahmad, I. Monitoring Permethrin and Imidacloprid Resistance in Indonesian House Fly Musca domestica L. (Diptera: Muscidae). J. Entomol. 2015, 13, 40–47. [Google Scholar] [CrossRef]
- Hubbard, C.B.; Gerry, A.C. Selection, Reversion, and Characterization of House Fly (Diptera: Muscidae) Behavioral Resistance to the Insecticide Imidacloprid. J. Med. Entomol. 2020, 57, 1843–1851. [Google Scholar] [CrossRef]
- Masuhisa, T. Methods for the Linkage-Group Determination of Insecticide Resistance Factors in the Housefly. 防虫科学 1964, 29, 51–59. [Google Scholar]
- Hubbard, C.B.; Gerry, A.C. Genetic Evaluation and Characterization of Behavioral Resistance to Imidacloprid in the House Fly. Pestic. Biochem. Physiol. 2021, 171, 104741. [Google Scholar] [CrossRef]
- Chen, Y.-C.D.; Menon, V.; Joseph, R.M.; Dahanukar, A.A. Control of Sugar and Amino Acid Feeding via Pharyngeal Taste Neurons. J. Neurosci. 2021, 41, 5791–5808. [Google Scholar] [CrossRef]
- King, B.H.; Gunathunga, P.B. Gustation in Insects: Taste Qualities and Types of Evidence Used to Show Taste Function of Specific Body Parts. J. Insect Sci. 2023, 23, 11. [Google Scholar] [CrossRef]
- Freeman, E.G.; Dahanukar, A. Molecular Neurobiology of Drosophila Taste. Curr. Opin. Neurobiol. 2015, 34, 140–148. [Google Scholar] [CrossRef]
- Adams, J.R.; Holbert, P.E.; Forgash, A.J. Electron Microscopy of the Contact Chemoreceptors of the Stable Fly, Stomoxys Calcitrans (Diptera: Muscidae). Ann. Entomol. Soc. Am. 1965, 58, 909–917. [Google Scholar] [CrossRef]
- Rajashekhar, K.P.; Singh, R.N. Organization of Motor Neurons Innervating the Proboscis Musculature in Drosophila melanogaster Meigen (Diptera: Drosophilidae). Int. J. Insect Morphol. Embryol. 1994, 23, 225–242. [Google Scholar] [CrossRef]
- Dunipace, L.; Meister, S.; McNealy, C.; Amrein, H. Spatially Restricted Expression of Candidate Taste Receptors in the Drosophila Gustatory System. Curr. Biol. 2001, 11, 822–835. [Google Scholar] [CrossRef]
- Scott, J.G. Evolution of Resistance to Pyrethroid Insecticides in Musca domestica. Pest Manag. Sci. 2017, 73, 716–722. [Google Scholar] [CrossRef] [PubMed]
- Zahn, L.; Gerry, A. House Fly (Musca domestica) laboratory rearing protocol—Gerry lab (UC Riverside). protocols.io. 2018. [CrossRef]
- Shiraiwa, T.; Carlson, J.R. Proboscis Extension Response (PER) Assay in Drosophila. J. Vis. Exp. 2007, 3, 193. [Google Scholar] [CrossRef]
- Hubbard, C.B.; Murillo, A.C. Concentration Dependent Feeding on Imidacloprid by Behaviorally Resistant House Flies, Musca domestica L. (Diptera: Muscidae). J. Med. Entomol. 2022, 59, 2066–2071. [Google Scholar] [CrossRef] [PubMed]
- R Core Team (2021). Available online: https://www.R-project.org/ (accessed on 21 December 2023).
- Smith, D.V.; Bowdan, E.; Dethier, V.G. Information Transmission in Tarsal Sugar Receptors of the Blowfly. Chem. Senses 1983, 8, 81–101. [Google Scholar] [CrossRef]
- Edgecomb, R.S.; Murdock, L.L.; Smith, A.B.; Stephen, M.D. Regulation of Tarsal Taste Threshold in the Blowfly, Phormia Regina. J. Exp. Biol. 1987, 127, 79–94. [Google Scholar] [CrossRef]
- Stoffolano, J.G.; Angioy, A.M.; Crnjar, R.; Liscia, A.; Pietra, P. Electrophysiological Recordings from the Taste Chemosensilla of Tabanus Nigrovittatus (Diptera: Tabanidae) and Determination of Behavioral Tarsal Acceptance Thresholds. J. Med. Entomol. 1990, 27, 14–23. [Google Scholar] [CrossRef]
- Loy, F.; Solari, P.; Isola, M.; Crnjar, R.; Masala, C. Morphological and Electrophysiological Analysis of Tarsal Sensilla in the Medfly Ceratitis capitata (Wiedemann, 1824) (Diptera: Tephritidae). Ital. J. Zool. 2016, 83, 456–468. [Google Scholar] [CrossRef]
- Wang, Z.; Singhvi, A.; Kong, P.; Scott, K. Taste Representations in the Drosophila Brain. Cell 2004, 117, 981–991. [Google Scholar] [CrossRef]
- Dethier, V.G. The Hungry Fly: A Physiological Study of the Behavior Associated with Feeding; Harvard University Press: Cambridge, MA, USA, 1976; p. 489. [Google Scholar] [CrossRef]
- Marella, S.; Fischler, W.; Kong, P.; Asgarian, S.; Rueckert, E.; Scott, K. Imaging Taste Responses in the Fly Brain Reveals a Functional Map of Taste Category and Behavior. Neuron 2006, 49, 285–295. [Google Scholar] [CrossRef]
- Isono, K.; Morita, H. Molecular and Cellular Designs of Insect Taste Receptor System. Front. Cell. Neurosci. 2010, 4, 1398. [Google Scholar] [CrossRef] [PubMed]
- Stocker, R.F. The Organization of the Chemosensory System in Drosophila melanogaster: A Rewiew. Cell Tissue Res. 1994, 275, 3–26. [Google Scholar] [CrossRef] [PubMed]
- Falk, R.; Atidia, J. Mutation Affecting Taste Perception in Drosophila melanogaster. Nature 1975, 254, 325–326. [Google Scholar] [CrossRef] [PubMed]
- Calas, D.; Thiéry, D.; Marion-Poll, F. 20-Hydroxyecdysone Deters Oviposition and Larval Feeding in the European Grapevine Moth, Lobesia botrana. J. Chem. Ecol. 2006, 32, 2443–2454. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, W.; Li, Q.; Ali Khan, K.; Rahman Khan, F.; Ullah, F.; Azad, R.; Shi, Q.; Qi, G.; Lyu, L. Optimization of Treatment Blocking the Gustatory Sense and Feeding Ethogram of Red Imported Fire Ant, Solenopsis invicta Buren (Hymenoptera: Formicidae) to Sugar. J. King Saud Univ.—Sci. 2021, 33, 101555. [Google Scholar] [CrossRef]
- Gaaboub, I.; Schuppe, H.; Newland, P.L. Position-Dependent Sensitivity and Density of Taste Receptors on the Locust Leg Underlies Behavioural Effectiveness of Chemosensory Stimulation. J. Comp. Physiol. A 2005, 191, 281–289. [Google Scholar] [CrossRef]
- Yosano, S.; Kutsuwada, Y.; Akatsu, M.; Masuta, S.; Kakazu, R.; Masuoka, N.; Matsuda, K.; Hori, M. Taste Recognition through Tarsal Gustatory Sensilla Potentially Important for Host Selection in Leaf Beetles (Coleoptera: Chrysomelidae). Sci. Rep. 2020, 10, 4931. [Google Scholar] [CrossRef] [PubMed]
- Rajashekar, Y.; Nandagopal, B.; Shivanandappa, T. Botanicals as Grain Protectants. Psyche 2012, 2012, 646740. [Google Scholar] [CrossRef]
- Georghiou, G.P. The Evolution of Resistance to Pesticides. Annu. Rev. Ecol. Syst. 1972, 3, 133–168. [Google Scholar] [CrossRef]
- French, A.S.; Sellier, M.-J.; Ali Agha, M.; Guigue, A.; Chabaud, M.-A.; Reeb, P.D.; Mitra, A.; Grau, Y.; Soustelle, L.; Marion-Poll, F. Dual Mechanism for Bitter Avoidance in Drosophila. J. Neurosci. 2015, 35, 3990–4004. [Google Scholar] [CrossRef] [PubMed]
- Rimal, S.; Lee, Y. Molecular Sensor of Nicotine in Taste of Drosophila melanogaster. Insect Biochem. Mol. Biol. 2019, 111, 103178. [Google Scholar] [CrossRef]
- Masek, P.; Scott, K. Limited Taste Discrimination in Drosophila. Proc. Natl. Acad. Sci. USA 2010, 107, 14833–14838. [Google Scholar] [CrossRef]
- Wada-Katsumata, A.; Silverman, J.; Schal, C. Differential Inputs from Chemosensory Appendages Mediate Feeding Responses to Glucose in Wild-Type and Glucose-Averse German Cockroaches, Blattella germanica. Chem. Senses 2011, 36, 589–600. [Google Scholar] [CrossRef]
- Wada-Katsumata, A.; Silverman, J.; Schal, C. Changes in Taste Neurons Support the Emergence of an Adaptive Behavior in Cockroaches. Science 2013, 340, 972–975. [Google Scholar] [CrossRef]
- Wada-Katsumata, A.; Robertson, H.M.; Silverman, J.; Schal, C. Changes in the Peripheral Chemosensory System Drive Adaptive Shifts in Food Preferences in Insects. Front. Cell. Neurosci. 2018, 12, 281. [Google Scholar] [CrossRef]
- Scott, K.; Brady, R.; Cravchik, A.; Morozov, P.; Rzhetsky, A.; Zuker, C.; Axel, R. A Chemosensory Gene Family Encoding Candidate Gustatory and Olfactory Receptors in Drosophila. Cell 2001, 104, 661–673. [Google Scholar] [CrossRef]
- Montell, C. A Taste of the Drosophila Gustatory Receptors. Curr. Opin. Neurobiol. 2009, 19, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, C.B.; Gerry, A.C.; Murillo, A.C. Evaluation of the Stability of Physiological and Behavioral Resistance to Imidacloprid in the House Fly (Musca domestica L.) (Diptera: Muscidae). Pest Manag. Sci. 2024, 80, 1361–1366. [Google Scholar] [CrossRef] [PubMed]
- Mullens, B.A.; Gerry, A.C.; Diniz, A.N. Field and Laboratory Trials of a Novel Metaflumizone House Fly (Diptera: Muscidae) Bait in California. J. Econ. Entomol. 2010, 103, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Murillo, A.C.; Gerry, A.C.; Gallagher, N.T.; Peterson, N.G.; Mullens, B.A. Laboratory and Field Assessment of Cyantraniliprole Relative to Existing Fly Baits. Pest Manag. Sci. 2015, 71, 752–758. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial | Strain | Imidacloprid Concentration | Observation Time (s) | PER (Mean + SE) | p Value | W |
|---|---|---|---|---|---|---|
| Proboscis assay | UCR (susceptible) | High | 0 | 15.60 ± 0.98 | 0.27 | 18 |
| Low | 14.00 ± 1.18 | |||||
| High | 2 | 9.00 ± 1.38 | 0.98 | 13 | ||
| Low | 8.80 ± 1.28 | |||||
| High | 10 | 4.60 ± 0.81 | 0.66 | 10 | ||
| Low | 5.40 ± 0.75 | |||||
| BRS (resistant) | High | 0 | 18.40 ± 0.51 | 0.8 | 14.5 | |
| Low | 18.00 ± 0.71 | |||||
| High | 2 | 6.40 ± 0.68 | 0.007 | 0 | ||
| Low | 14.20 ± 0.97 | |||||
| High | 10 | 3.20 ± 0.58 | 0.006 | 0 | ||
| Low | 10.00 ± 1.30 |
| Trial | Strain | Imidacloprid Concentration | Observation Time (s) | PER (Mean + SE) | p Value | W |
|---|---|---|---|---|---|---|
| Tarsal assay | UCR (susceptible) | High | 0 | 13.8 ± 1.43 | 0.95 | 13 |
| Low | 13.0 ± 0.77 | |||||
| High | 2 | 9.4 ± 0.81 | 0.5 | 9 | ||
| Low | 10.0 ± 1.45 | |||||
| High | 10 | 5.6 ± 1.36 | 1 | 12.5 | ||
| Low | 5.4 ± 1.72 | |||||
| BRS (resistant) | High | 0 | 12.4 ± 2.25 | 0.26 | 6.5 | |
| Low | 16.0 ± 1.18 | |||||
| High | 2 | 6.4 ± 1.21 | 0.09 | 4 | ||
| Low | 10.0 ± 1.30 | |||||
| High | 10 | 4.0 ± 0.89 | 0.04 | 2.5 | ||
| Low | 7.2 ± 0.73 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Arco, S.; Maistrello, L.; Hubbard, C.B.; Murillo, A.C.; Gerry, A.C. Use of the Proboscis Extension Response Assay to Evaluate the Mechanism of House Fly Behavioral Resistance to Imidacloprid. Insects 2024, 15, 168. https://doi.org/10.3390/insects15030168
D’Arco S, Maistrello L, Hubbard CB, Murillo AC, Gerry AC. Use of the Proboscis Extension Response Assay to Evaluate the Mechanism of House Fly Behavioral Resistance to Imidacloprid. Insects. 2024; 15(3):168. https://doi.org/10.3390/insects15030168
Chicago/Turabian StyleD’Arco, Sara, Lara Maistrello, Caleb B. Hubbard, Amy C. Murillo, and Alec C. Gerry. 2024. "Use of the Proboscis Extension Response Assay to Evaluate the Mechanism of House Fly Behavioral Resistance to Imidacloprid" Insects 15, no. 3: 168. https://doi.org/10.3390/insects15030168