
Resistance Mechanism of Plutella xylostella (L.) Associated with Amino Acid Substitutions in Acetylcholinesterase-1: Insights from Homology Modeling, Docking and Molecular Dynamic Simulation

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. RNA Extraction, cDNA Synthesis, Cloning and Sequencing
2.3. qRT-PCR Assay
2.4. Computational Study
2.4.1. Structural Modeling
2.4.2. Structure Validation
2.4.3. Molecular Simulation
2.5. Data Analysis
3. Results
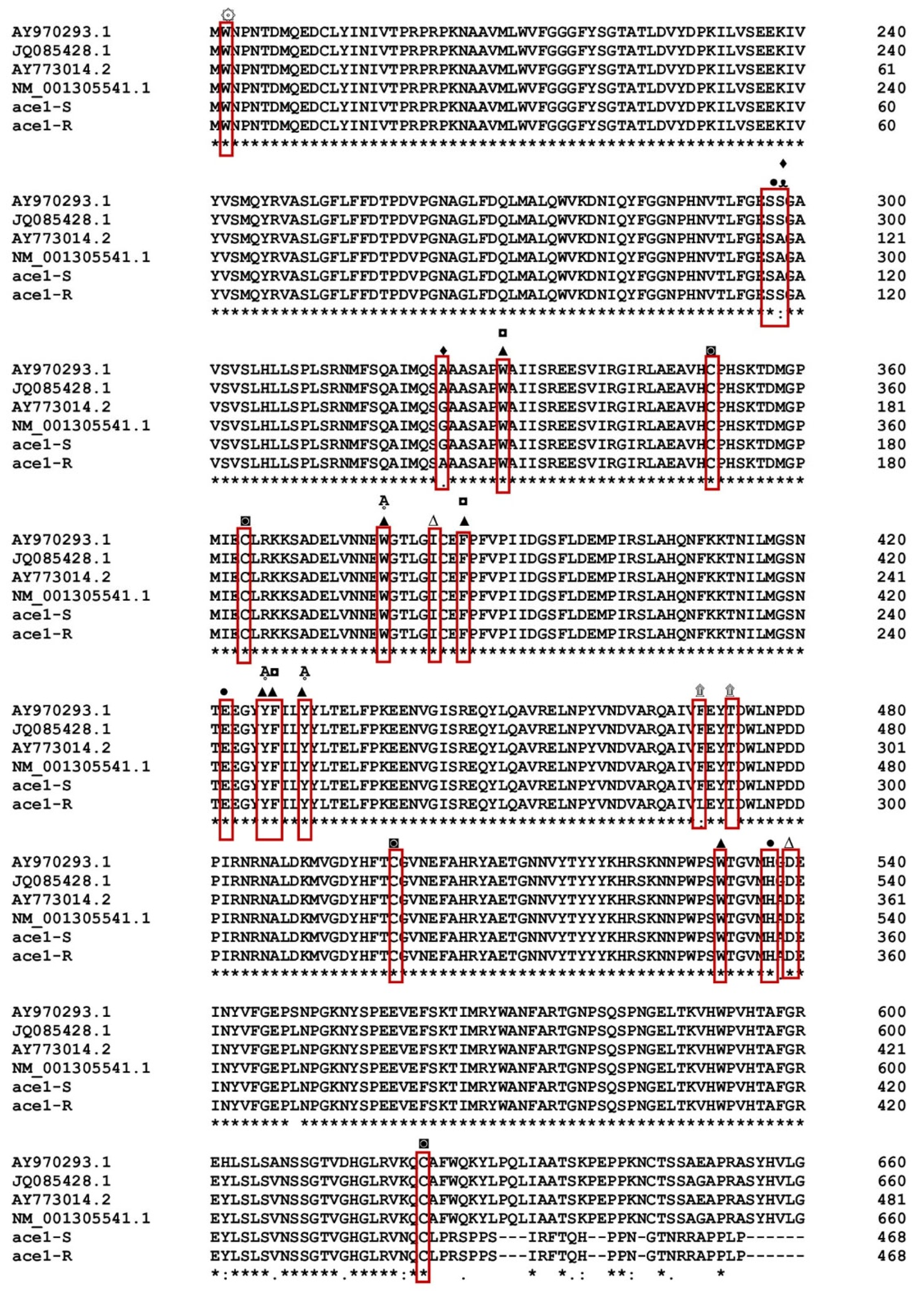
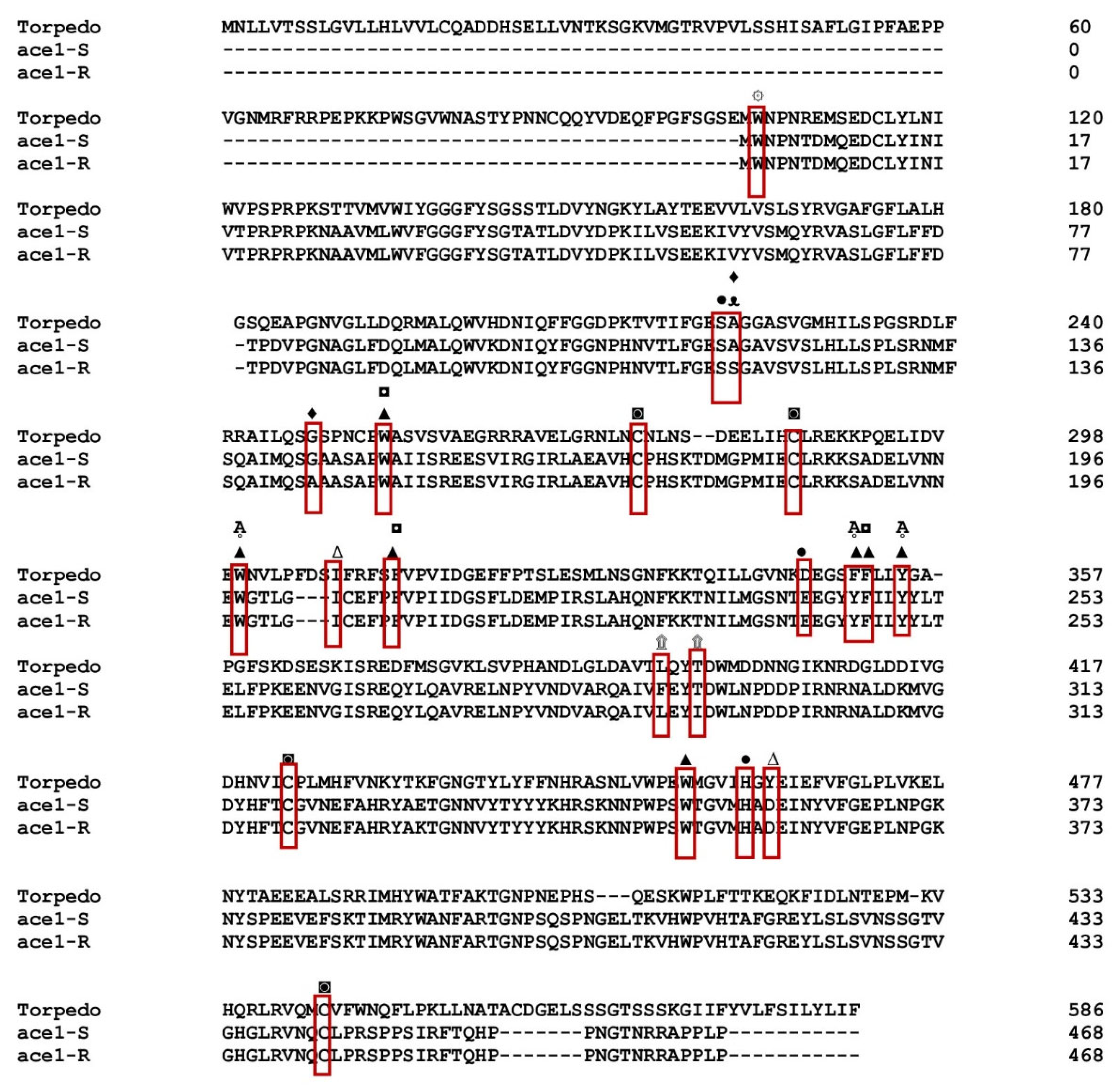
3.1. Gene Sequencing Revealed Amino Acid Substitutions in the ace1
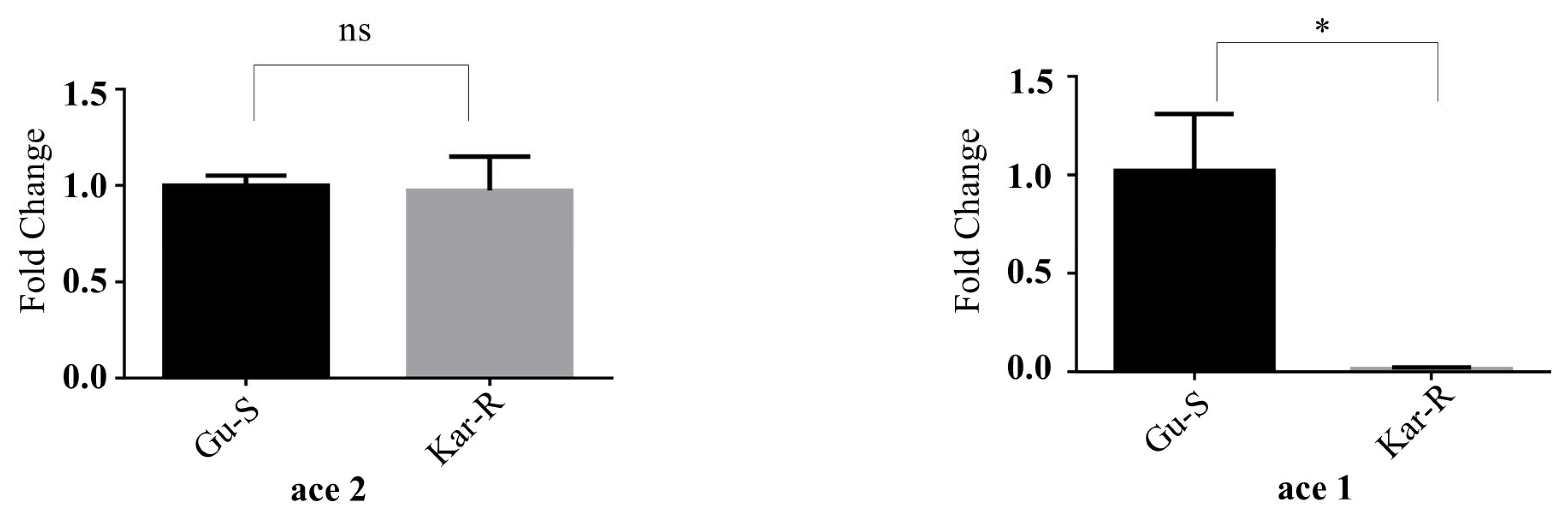
3.2. Significant Differences in ace1 and ace2 Gene Expression
3.3. Molecular Dynamics Simulations
3.3.1. Amino Acid Substitutions Altered Binding Energy between Mutant Protein and Ligand
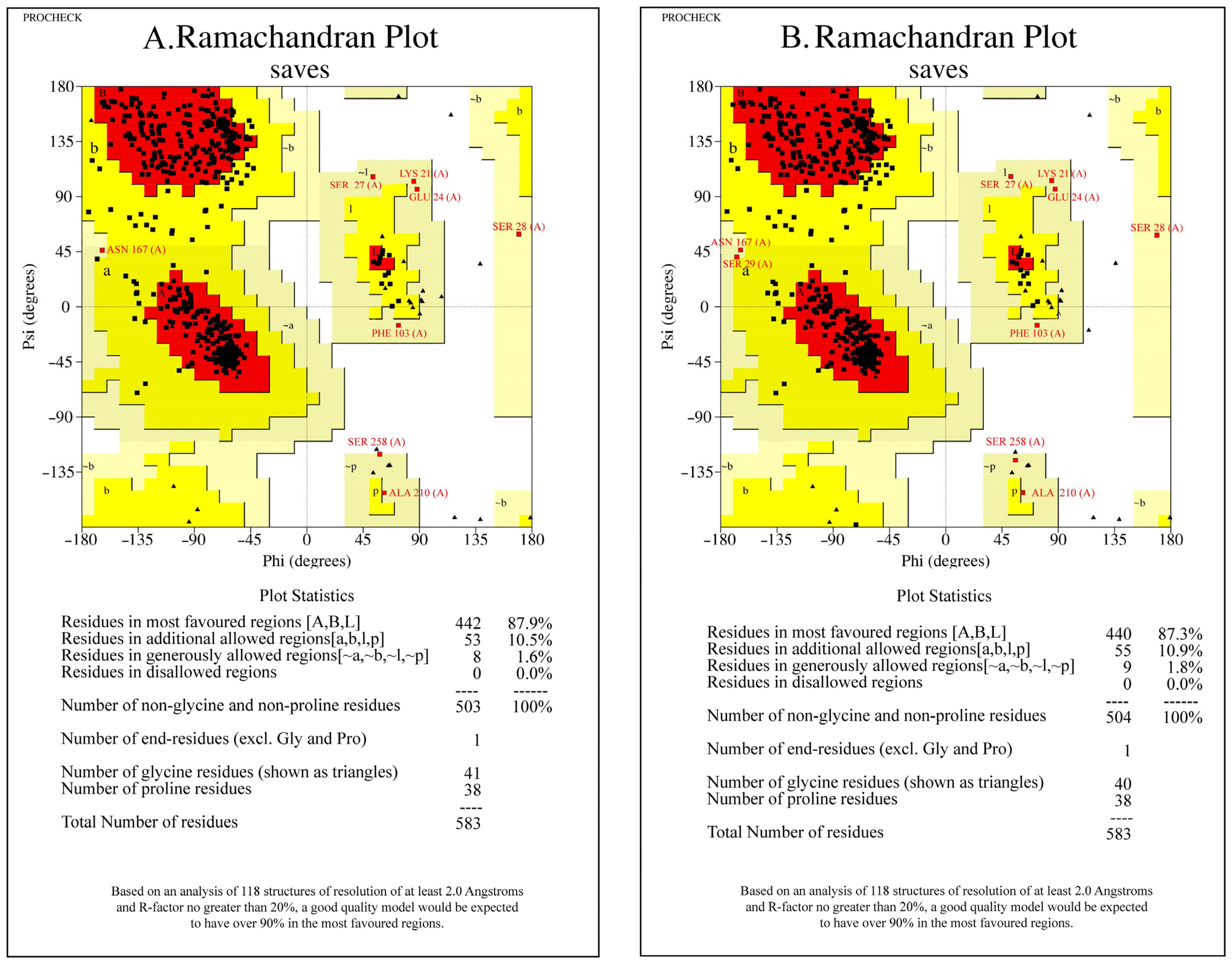
3.3.2. Ramachandran Plots Validated the Structural Models
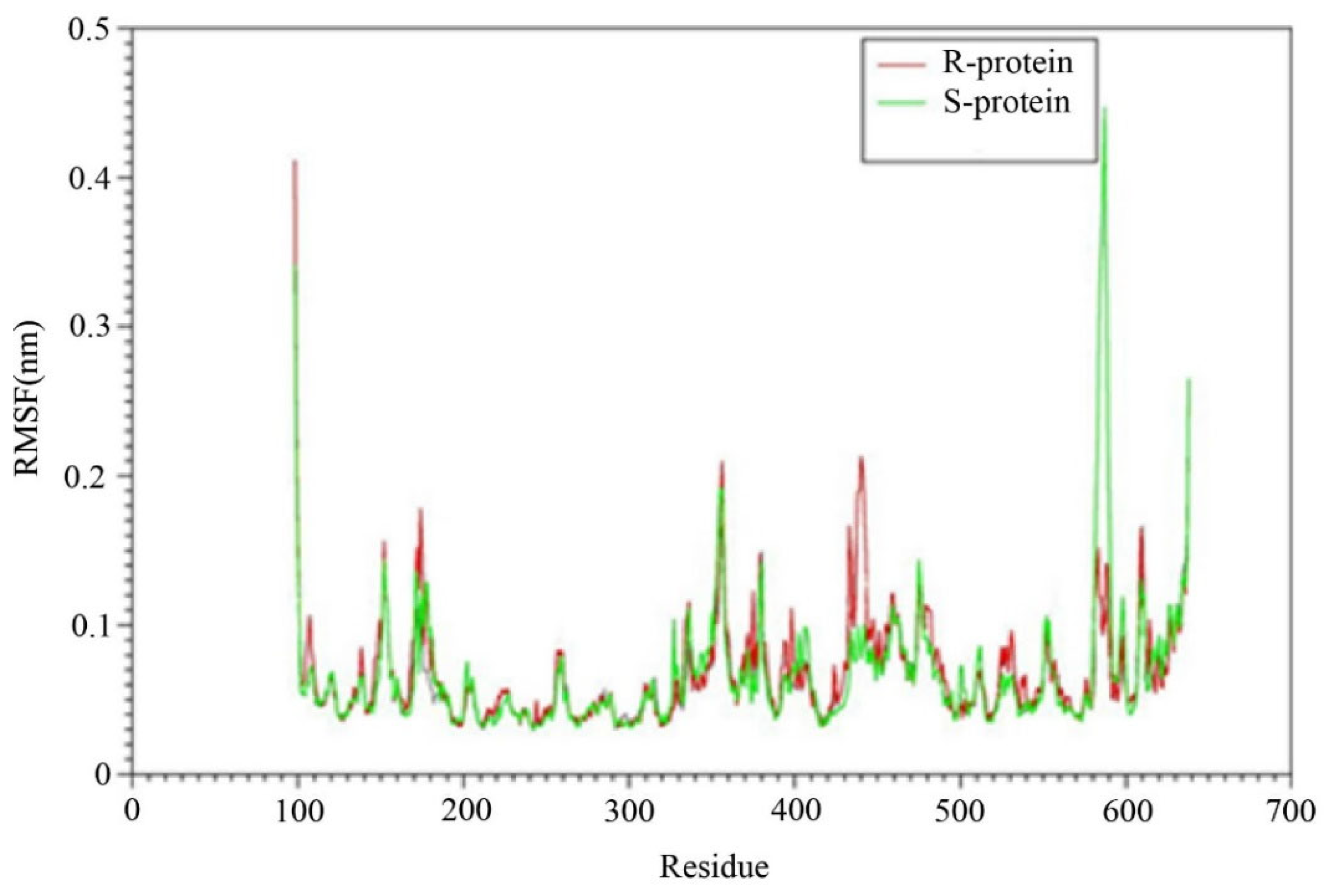
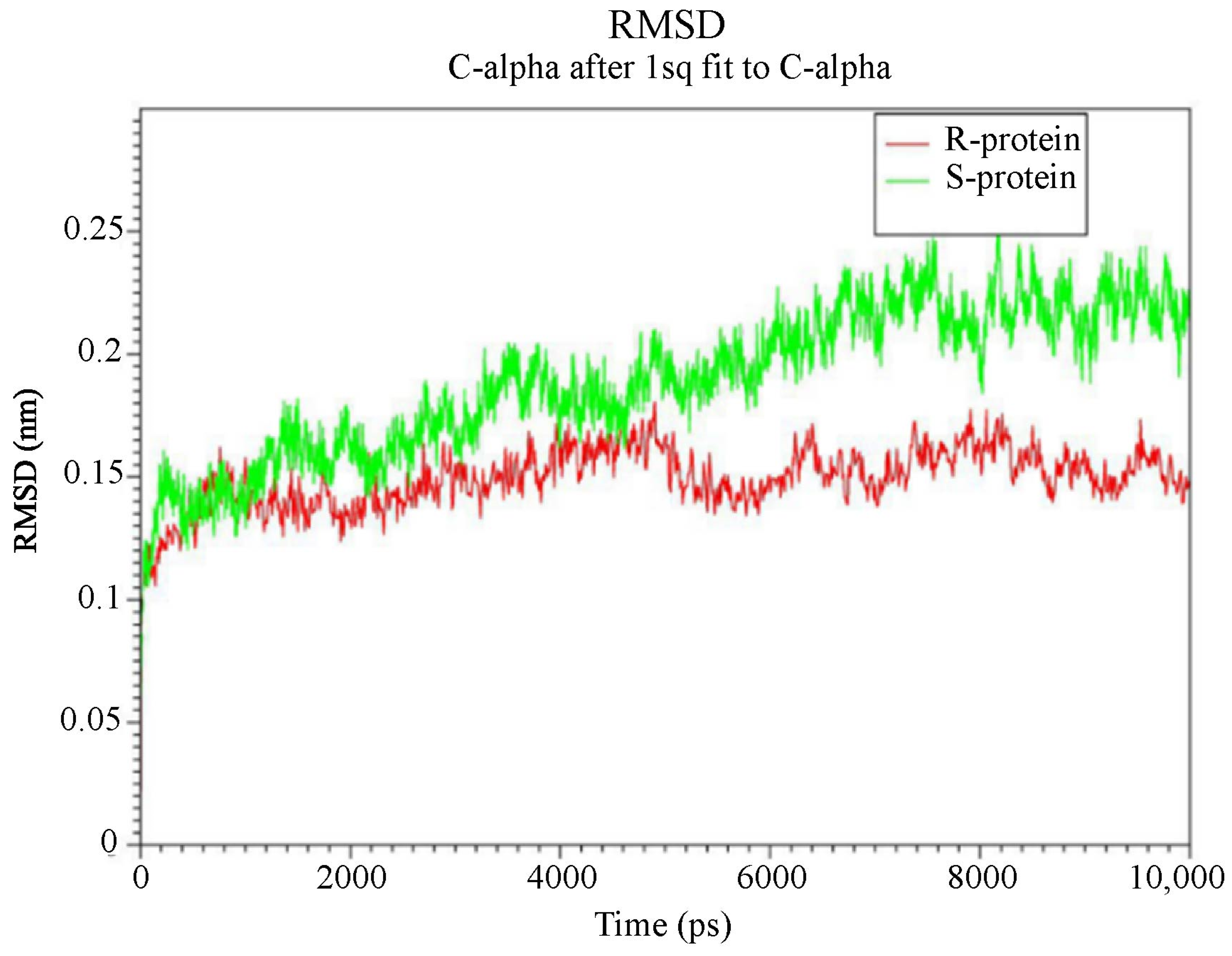
3.3.3. Higher Flexibility and Lower Affinity of Resistant AChE for Chlorpyriphos
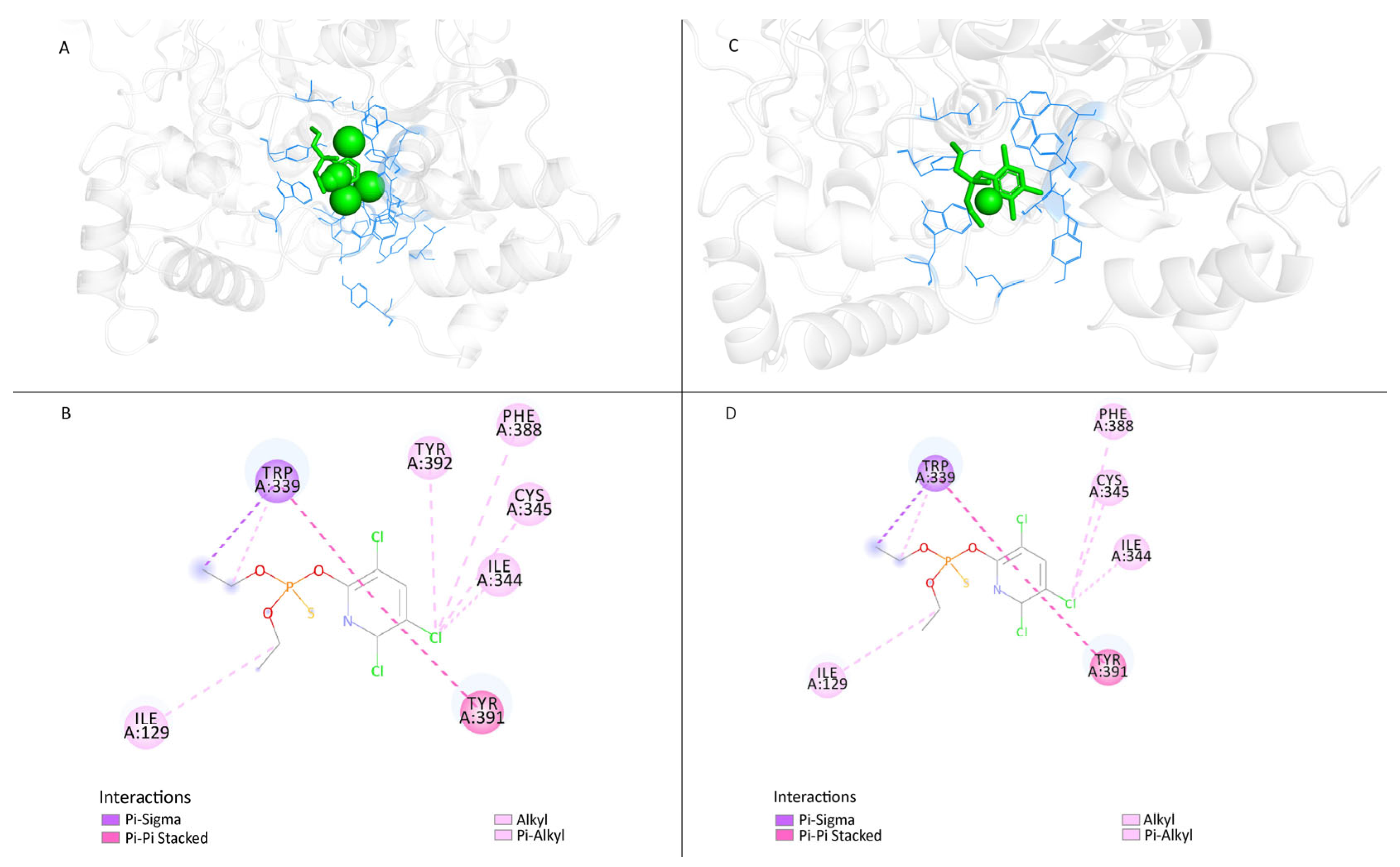
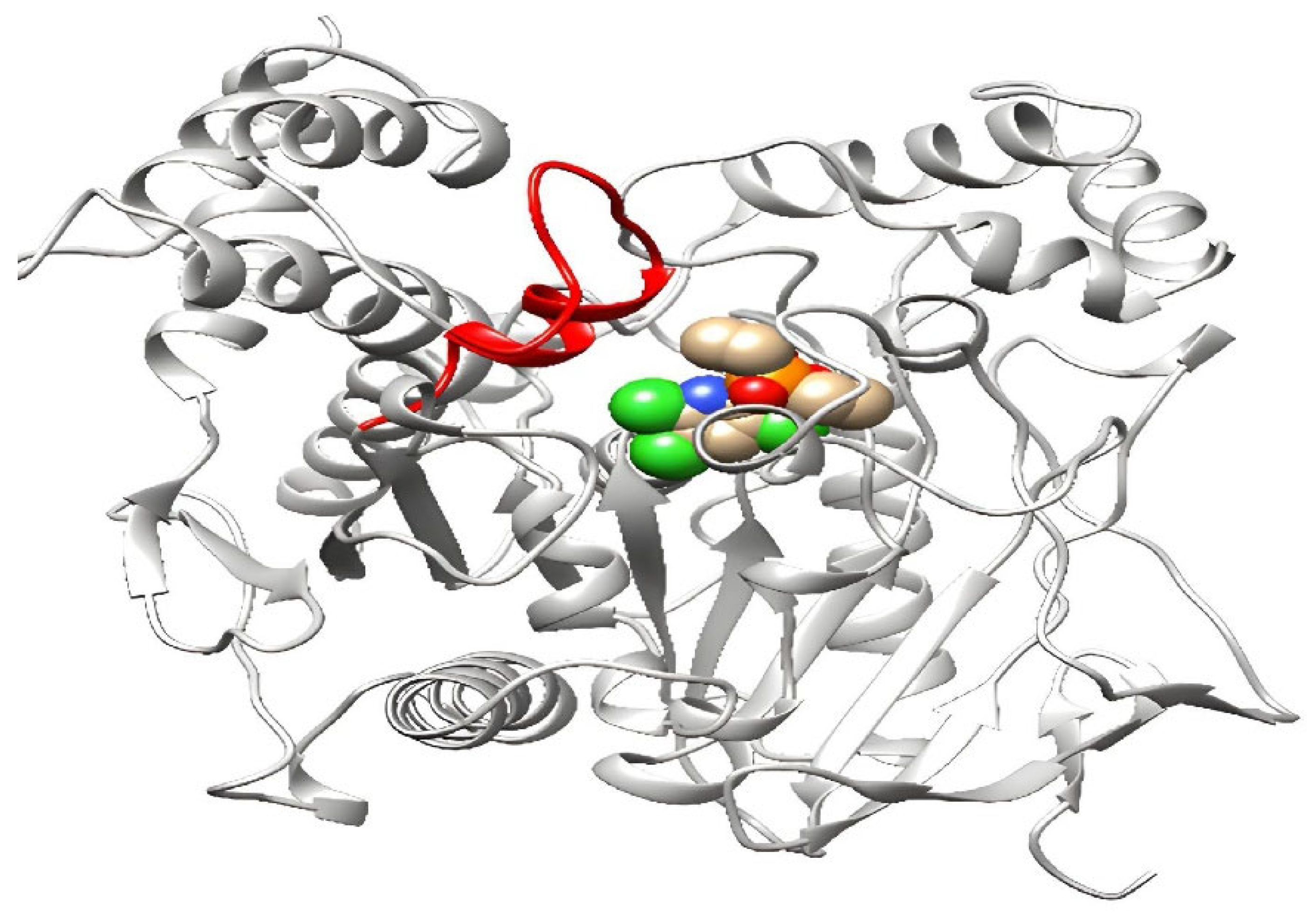
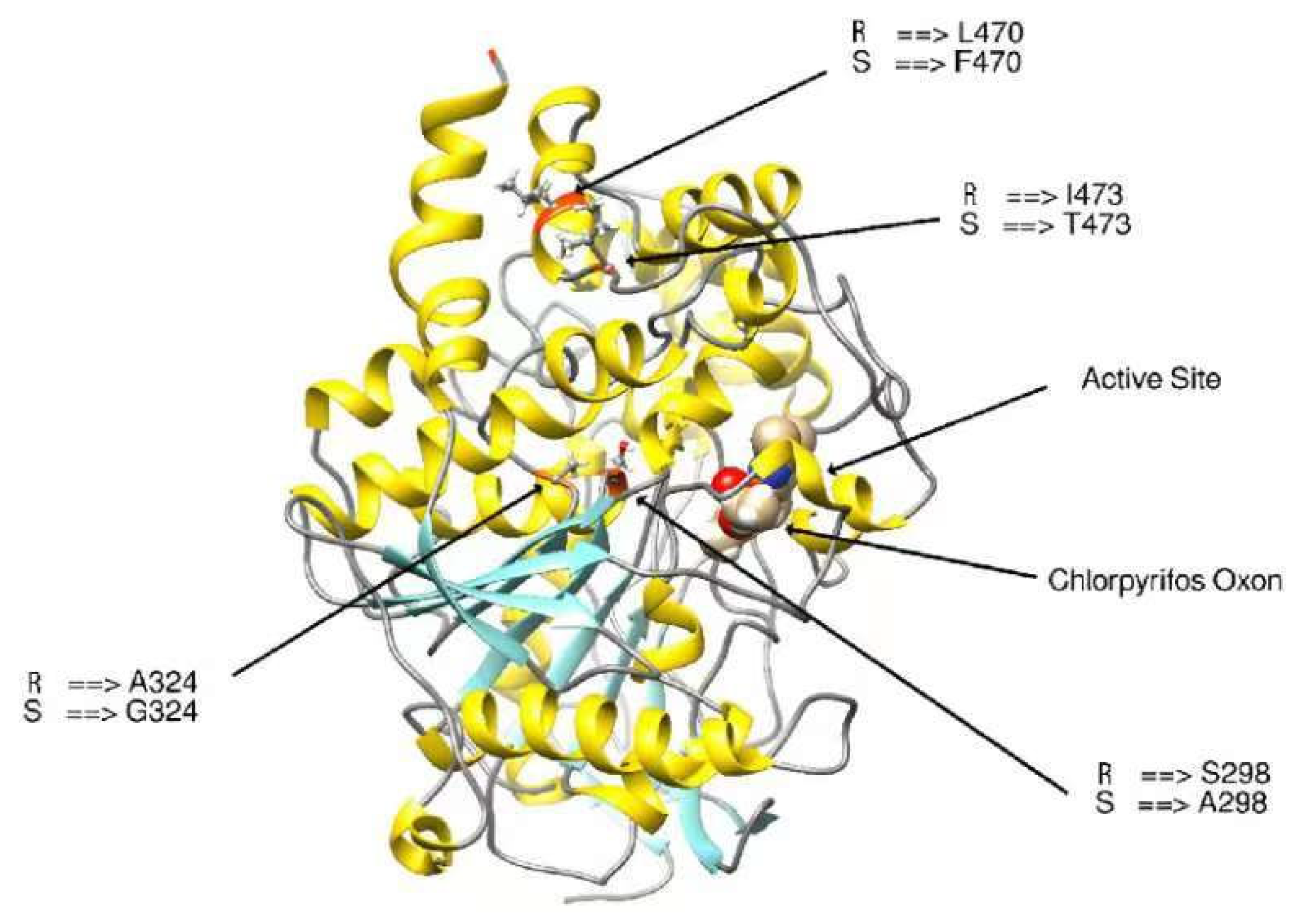
3.3.4. Modifying the Active Site of Resistant AChE Protein through Amino Acid Substitutions
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback moth ecology and management: Problems, progress, and prospects. Annu. Rev. Entomol. 2013, 58, 517–541. [Google Scholar] [CrossRef] [PubMed]
- Farias, E.S.; Santos, A.A.; Ribeiro, A.V.; Carmo, D.G.; Paes, J.S.; Picanco, M.C. Climate and host plants mediating seasonal dynamics and within-plant distribution of the diamondback moth (Plutella xylostella). Crop Prot. 2020, 134, 105172. [Google Scholar] [CrossRef]
- Vasquez, B.L. Resistant to most insecticides. In Book of Insects Records; Walker, T.J., Ed.; University of Florida: Gainesville, FL, USA, 1995; pp. 34–36. [Google Scholar]
- Nauen, R. Activities of the Insecticide Resistance Action Committee (IRAC): A brief introduction. Insecticide Resistance Action Committee. 2012. 18p. Available online: http://www.irac-online.org (accessed on 8 February 2024).
- Ware, G.W. Pesticides: Chemical Tools. The Pesticide Book; Thomson Publications: Fresno, CA, USA, 1994; pp. 2–20. [Google Scholar]
- Toutant, J.-P. Insect acetylcholinesterase: Catalytic properties, tissue distribution and molecular forms. Prog. Neurobiol. 1989, 32, 423–446. [Google Scholar] [CrossRef] [PubMed]
- Bourguet, D.; Raymond, M.; Fournier, D.; Malcolm, C.A.; Toutant, J.P.; Arpagaus, M. Existence of two acetylcholinesterases in the mosquito Culex pipiens (Diptera: Culicidae). J. Neurochem. 1996, 67, 2115–2123. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Kim, J.I.; Lee, D.-W.; Chung, B.K.; Miyata, T.; Lee, S.H. Identification and characterization of ace1-type acetylcholinesterase likely associated with organophosphate resistance in Plutella xylostella. Pestic. Biochem. Physiol. 2005, 81, 164–175. [Google Scholar] [CrossRef]
- Nabeshima, T.; Kozaki, T.; Tomita, T.; Kono, Y. An amino acid substitution on the second acetylcholinesterase in the pirimicarb-resistant strains of the peach potato aphid, Myzus persicae. Biochem. Biophys. Res. Commun. 2003, 307, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Weill, M.; Lutfalla, G.; Mogensen, K.; Chandre, F.; Berthomieu, A.; Berticat, C.; Pasteur, N.; Philips, A.; Fort, P.; Raymond, M. Insecticide resistance in mosquito vectors. Nature 2003, 423, 136–137. [Google Scholar] [CrossRef] [PubMed]
- Nabeshima, T.; Mori, A.; Kozaki, T.; Iwata, Y.; Hidoh, O.; Harada, S.; Kasai, S.; Severson, D.W.; Kono, Y.; Tomita, T. An amino acid substitution attributable to insecticide-insensitivity of acetylcholinesterase in a Japanese encephalitis vector mosquito, Culex tritaeniorhynchus. Biochem. Biophys. Res. Commun. 2004, 313, 794–801. [Google Scholar] [CrossRef]
- Omuut, G.; Mollel, H.G.; Kanyesigye, D.; Akohoue, F.; Adumo Aropet, S.; Wagaba, H.; Otim, M.H. Genetic analyses and detection of point mutations in the acetylcholinesterase-1 gene associated with organophosphate insecticide resistance in fall armyworm (Spodoptera frugiperda) populations from Uganda. BMC Genom. 2023, 24, 22. [Google Scholar] [CrossRef]
- Lee, D.-W.; Choi, J.Y.; Kim, W.T.; Je, Y.H.; Song, J.T.; Chung, B.K.; Boo, K.S.; Koh, Y.H. Mutations of acetylcholinesterase1 contribute to prothiofos-resistance in Plutella xylostella (L.). Biochem. Biophys. Res. Commun. 2007, 353, 591–597. [Google Scholar] [CrossRef]
- Fukuto, T.R. Mechanism of action of organophosphorus and carbamate insecticides. Environ. Health Perspect. 1990, 87, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Bajgar, J. Organophosphates/nerve agent poisoning: Mechanism of action, diagnosis, prophylaxis, and treatment. Adv. Clin. Chem. 2004, 38, 151–216. [Google Scholar]
- Fournier, D.; Mutero, A. Modification of acetylcholinesterase as a mechanism of resistance to insecticides. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1994, 108, 19–31. [Google Scholar] [CrossRef]
- Malekmohammadi, M. Resistance of Colorado potato beetle (Leptinotarsa decemlineata Say) to commonly used insecticides in Iran. J. Asia-Pac. Entomol. 2014, 17, 213–220. [Google Scholar] [CrossRef]
- Vontas, J.G.; Hejazi, M.; Hawkes, N.J.; Cosmidis, N.; Loukas, M.; Hemingway, J. Resistance-associated point mutations of organophosphate insensitive acetylcholinesterase, in the olive fruit fly Bactrocera oleae. Insect Mol. Biol. 2002, 11, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.B.; Dolden, T.A.; Moores, G.D.; Kristensen, M.; Lewis, T.; Devonshire, A.L.; Williamson, M.S. Identification and characterization of mutations in housefly (Musca domestica) acetylcholinesterase involved in insecticide resistance. Biochem. J. 2001, 359, 175–181. [Google Scholar] [CrossRef]
- Mutero, A.; Pralavorio, M.; Bride, J.-M.; Fournier, D. Resistance-associated point mutations in insecticide-insensitive acetylcholinesterase. Proc. Natl. Acad. Sci. USA 1994, 91, 5922–5926. [Google Scholar] [CrossRef]
- Zolfaghari, M.; Ghadamyari, M.; Hassan Sajedi, R. Resistance mechanisms of a field population of diamond back moth, Plutella xylostella (Lepidoptera: Plutellidae) to current organophosphate pesticides. J. Crop Prot. 2019, 8, 403–416. [Google Scholar]
- Sindhu, T.; Venkatesan, T.; Prabhu, D.; Jeyakanthan, J.; Gracy, G.R.; Jalali, S.K.; Rai, A. Insecticide-resistance mechanism of Plutella xylostella (L.) associated with amino acid substitutions in acetylcholinesterase-1: A molecular docking and molecular dynamics investigation. Comput. Biol. Chem. 2018, 77, 240–250. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- DeLano, W.L. Pymol: An open-source molecular graphics tool. CCP4 Newsl. Protein Crystallogr. 2002, 40, 82–92. [Google Scholar]
- Ramachandran, G.t.; Sasisekharan, V. Conformation of polypeptides and proteins. Adv. Protein Chem. 1968, 23, 283–437. [Google Scholar] [PubMed]
- Ho, B.K.; Brasseur, R. The Ramachandran plots of glycine and pre-proline. BMC Struct. Biol. 2005, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, Y. LOMETS: A local meta-threading-server for protein structure prediction. Nucleic Acids Res. 2007, 35, 3375–3382. [Google Scholar] [CrossRef]
- Laskowski, R.A. Structural quality assurance. In Structural Bioinformatics, 2nd ed.; Wiley: Hoboken, NJ, USA, 2009; Volume 2. [Google Scholar]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef]
- Berendsen, H.J.; Postma, J.v.; Van Gunsteren, W.F.; Dinola, A.; Haak, J.R. Molecular dynamics with coupling to an external bath. J. Chem. Phys. 1984, 81, 3684–3690. [Google Scholar] [CrossRef]
- Case, D.A.; Darden, T.A.; Cheatham, T.E.; Simmerling, C.L.; Wang, J.; Duke, R.E.; Luo, R.; Crowley, M.; Walker, R.C.; Zhang, W. Amber 10; University of California: Los Angeles, CA, USA, 2008. [Google Scholar]
- Kim, Y.H.; Lee, S.H. Invertebrate acetylcholinesterases: Insights into their evolution and non-classical functions. J. Asia-Pac. Entomol. 2018, 21, 186–195. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, Y.H.; Kwon, D.H.; Cha, D.J.; Kim, J.H. Mutation and duplication of arthropod acetylcholinesterase: Implications for pesticide resistance and tolerance. Pestic. Biochem. Physiol. 2015, 120, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Talebi-Jahromi, K.; Hosseininaveh, V.; Ghadamyari, M. Toxicological and biochemical characterizations of AChE in phosalone-susceptible and resistant populations of the common pistachio psyllid, Agonoscena pistaciae. J. Insect Sci. 2014, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.-G.; Wang, S.; Dou, T.-T.; Liu, S.; Li, M.-Y.; Hua, R.-M.; Li, S.-G.; Lin, H.-F. Aphicidal activity of Illicium verum fruit extracts and their effects on the acetylcholinesterase and glutathione S-transferases activities in Myzus persicae (Hemiptera: Aphididae). J. Insect Sci. 2016, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Lee, S.H. Which acetylcholinesterase functions as the main catalytic enzyme in the Class Insecta? Insect Biochem. Mol. Biol. 2013, 43, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, S.; Shi, X.; Song, D.; Liang, P.; Gao, X.; Zhang, Y.; Li, J.; Liu, Y.; Li, M.; Matsumura, M. Duplication of acetylcholinesterase gene in diamondback moth strains with different sensitivities to acephate. Insect Biochem. Mol. Biol. 2014, 48, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Han, Z.; Wang, Y. Mechanisms of monocrotophos resistance in cotton bollworm, Helicoverpa armigera (Hübner). Arch. Insect Biochem. Physiol. Publ. Collab. Entomol. Soc. Am. 2002, 51, 103–110. [Google Scholar] [CrossRef]
- Chen, J.; Hua, G.; Jurat-Fuentes, J.L.; Abdullah, M.A.; Adang, M.J. Synergism of Bacillus thuringiensis toxins by a fragment of a toxin-binding cadherin. Proc. Natl. Acad. Sci. USA 2007, 104, 13901–13906. [Google Scholar] [CrossRef]
- Hemingway, J.; Hawkes, N.J.; McCarroll, L.; Ranson, H. The molecular basis of insecticide resistance in mosquitoes. Insect Biochem. Mol. Biol. 2004, 34, 653–665. [Google Scholar] [CrossRef]
- Gressel, J. Evolving understanding of the evolution of herbicide resistance. Pest Manag. Sci. Former. Pestic. Sci. 2009, 65, 1164–1173. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H. The molecular genetics of insecticide resistance. Genetics 2013, 194, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, S.; Igaki, C. Characterization of acephate resistance in the diamondback moth Plutella xylostella. Pestic. Biochem. Physiol. 2010, 98, 121–127. [Google Scholar] [CrossRef]
- Yeh, S.-C.; Lin, C.-L.; Chang, C.; Feng, H.-T.; Dai, S.-M. Amino acid substitutions and intron polymorphism of acetylcholinesterase1 associated with mevinphos resistance in diamondback moth, Plutella xylostella (L.). Pestic. Biochem. Physiol. 2014, 112, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Fournier, D. Mutations of acetylcholinesterase which confer insecticide resistance in insect populations. Chem.-Biol. Interact. 2005, 157, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Murúa, M.G.; Nagoshi, R.N.; Dos Santos, D.A.; Hay-Roe, M.M.; Meagher, R.L.; Vilardi, J.C. Demonstration using field collections that Argentina fall armyworm populations exhibit strain-specific host plant preferences. J. Econ. Entomol. 2015, 108, 2305–2315. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Han, Z. Mutations in acetylcholinesterase associated with insecticide resistance in the cotton aphid, Aphis gossypii Glover. Insect Biochem. Mol. Biol. 2004, 34, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.; Callaghan, A.; Field, L.; Williamson, M.; Moores, G. Identification of mutations conferring insecticide-insensitive AChE in the cotton-melon aphid, Aphis gossypii Glover. Insect Mol. Biol. 2004, 13, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zuo, K.; Kang, Z.; Yang, Y.; Oakeshott, J.G.; Wu, Y. A point mutation in the acetylcholinesterase-1 gene is associated with chlorpyrifos resistance in the plant bug Apolygus lucorum. Insect Biochem. Mol. Biol. 2015, 65, 75–82. [Google Scholar] [CrossRef]
- Jiang, X.; Qu, M.; Denholm, I.; Fang, J.; Jiang, W.; Han, Z. Mutation in acetylcholinesterase1 associated with triazophos resistance in rice stem borer, Chilo suppressalis (Lepidoptera: Pyralidae). Biochem. Biophys. Res. Commun. 2009, 378, 269–272. [Google Scholar] [CrossRef]
- Menozzi, P.; Shi, M.A.; Lougarre, A.; Tang, Z.H.; Fournier, D. Mutations of acetylcholinesterase which confer insecticide resistance in Drosophila melanogaster populations. BMC Evol. Biol. 2004, 4, 1–7. [Google Scholar] [CrossRef]
- Moradi, S.; Khani, S.; Ansari, M.; Shahlaei, M. Atomistic details on the mechanism of organophosphates resistance in insects: Insights from homology modeling, docking and molecular dynamic simulation. J. Mol. Liq. 2019, 276, 59–66. [Google Scholar] [CrossRef]
- Yu, R.; Xu, X.; Liang, Y.; Tian, H.; Pan, Z.; Jin, S.; Wang, N.; Zhang, W. The insect ecdysone receptor is a good potential target for RNAi-based pest control. Int. J. Biol. Sci. 2014, 10, 1171. [Google Scholar] [CrossRef]
- Gao, J.-R.; Zhu, K.Y. Increased expression of an acetylcholinesterase gene may confer organophosphate resistance in the greenbug, Schizaphis graminum (Homoptera: Aphididae). Pestic. Biochem. Physiol. 2002, 73, 164–173. [Google Scholar] [CrossRef]
- Pan, Y.; Shang, Q.; Fang, K.; Zhang, J.; Xi, J. Down-regulated transcriptional level of Ace1 combined with mutations in Ace1 and Ace2 of Aphis gossypii are related with omethoate resistance. Chem.-Biol. Interact. 2010, 188, 553–557. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence (5′-3′) | Primer | Amplicon Length (bp) | E a (%) | R2 b | Purpose |
|---|---|---|---|---|---|
| F: CTACAACCCCGAGCTGGACACCATC R: ACACTCGATCATAGCCCATGTCAGTC | ace1-FM2 ace-R2 | 900 | 98.4 | 995 | ace1 ORF-PCR ace1 ORF-PCR |
| F: GACACTCCCGATGTCCCTGGAAAC R: TGGCTGAAACTAACGGCTGCGACG | ace1-F2 ace-RM | 1200 | 98.3 | 0.999 | ace1 ORF-PCR ace1 ORF-PCR |
| F: TACGCCAAGACCGTGATGGGAG R:CGAAGTAGTTGGTGGGACACACGAAGAAG | ace-FM5 ace-RM | 1400 | 99.3 | 0.997 | ace2 ORF-PCR ace2 ORF-PCR |
| F: GACACTCCCGATGTCCCTGGAAAC R: ACACTCGATCATAGCCCATGTCAGTC | ace-F2 ace1-R2 | 300 | 99.2 | 0.995 | ace1 expression-qPCR |
| F: AATGCACCGTGGAGTTGGATGACAGG R: TTTTGATTGCTTCCAAGAAGAACTTC | ace2-F2 ace2-R2 | 250 | 98.4 | 0.996 | ace2 expression-qPCR |
| F: GTTGTTGGGAAGTTGACC R: CAGTGCGGCATTCAGT | 18SrRNA-F 18SrRNA-R | 187 | 96.9 | 0.997 | Reference gene for ace1 and ace2 |
| F: CCAATTTACCGCCCTACC R: TACCCTGTTGTCAATACCTCT | RPL32-F RPL32-R | 168 | 103.8 | 0.997 | Reference gene for ace1 and ace2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zolfaghari, M.; Xiao, Y.; Safiul Azam, F.M.; Yin, F.; Peng, Z.-K.; Li, Z.-Y. Resistance Mechanism of Plutella xylostella (L.) Associated with Amino Acid Substitutions in Acetylcholinesterase-1: Insights from Homology Modeling, Docking and Molecular Dynamic Simulation. Insects 2024, 15, 144. https://doi.org/10.3390/insects15030144
Zolfaghari M, Xiao Y, Safiul Azam FM, Yin F, Peng Z-K, Li Z-Y. Resistance Mechanism of Plutella xylostella (L.) Associated with Amino Acid Substitutions in Acetylcholinesterase-1: Insights from Homology Modeling, Docking and Molecular Dynamic Simulation. Insects. 2024; 15(3):144. https://doi.org/10.3390/insects15030144
Chicago/Turabian StyleZolfaghari, Maryam, Yong Xiao, Fardous Mohammad Safiul Azam, Fei Yin, Zheng-Ke Peng, and Zhen-Yu Li. 2024. "Resistance Mechanism of Plutella xylostella (L.) Associated with Amino Acid Substitutions in Acetylcholinesterase-1: Insights from Homology Modeling, Docking and Molecular Dynamic Simulation" Insects 15, no. 3: 144. https://doi.org/10.3390/insects15030144