
Phylogeny of Rhynchium and Its Related Genera (Hymenoptera: Eumeninae) Based on Universal Single-Copy Orthologs and Ultraconserved Elements
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Sequencing
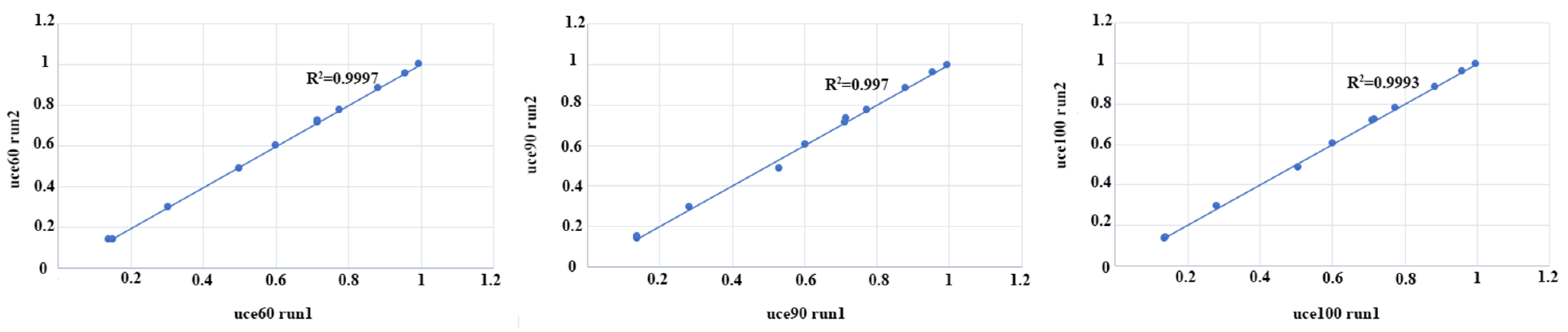
2.2. Genome Assembly and Matrix Generation
2.3. Phylogenetic Analyses
2.4. Hypothesis Testing
2.5. Divergence Time Analyses
3. Results
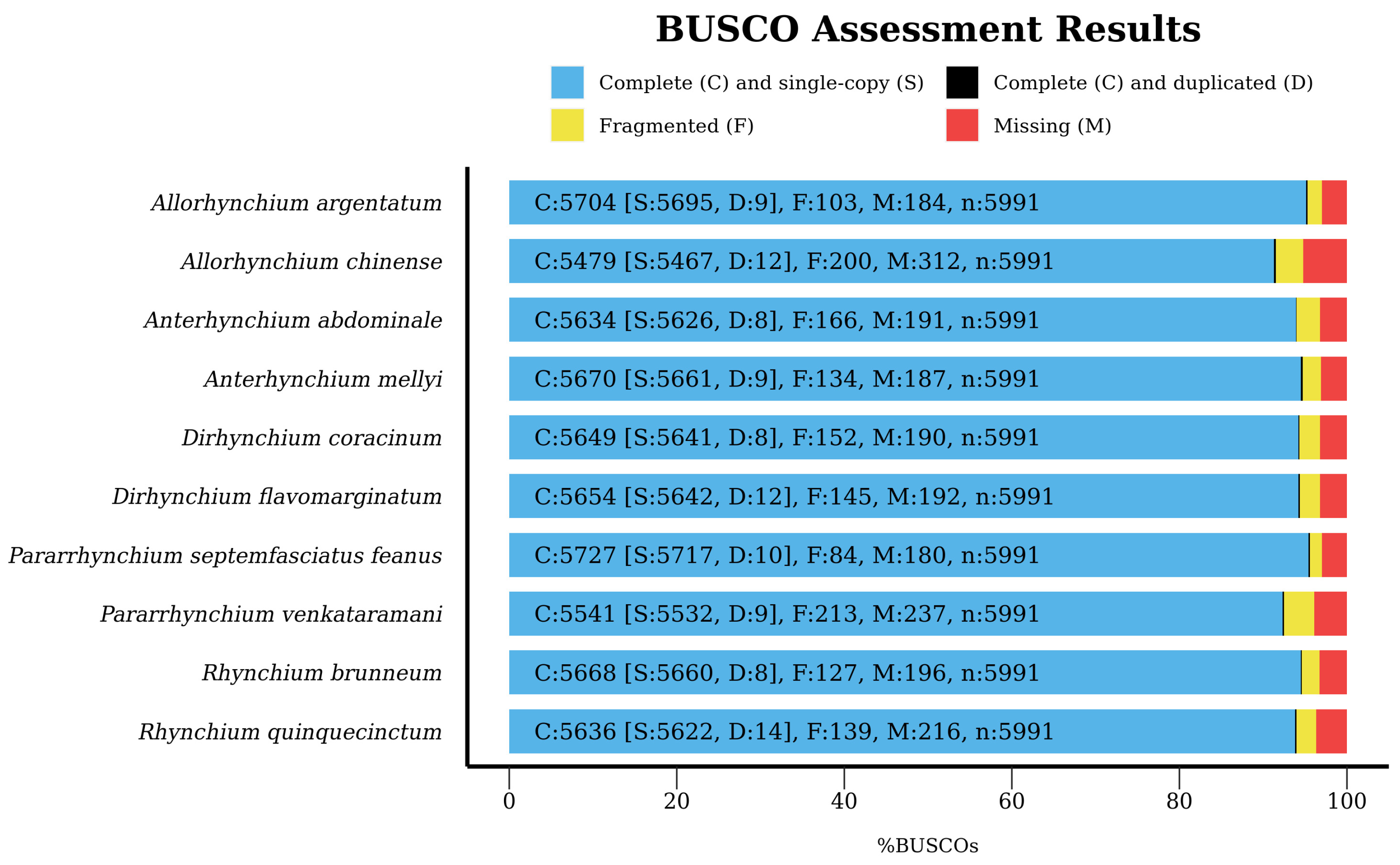
3.1. Genome Assembly and Matrix Generation
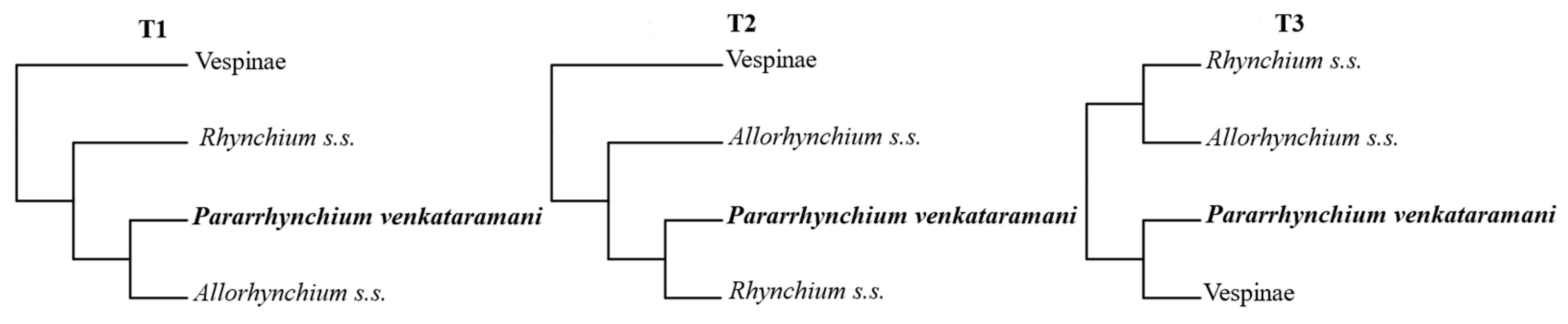
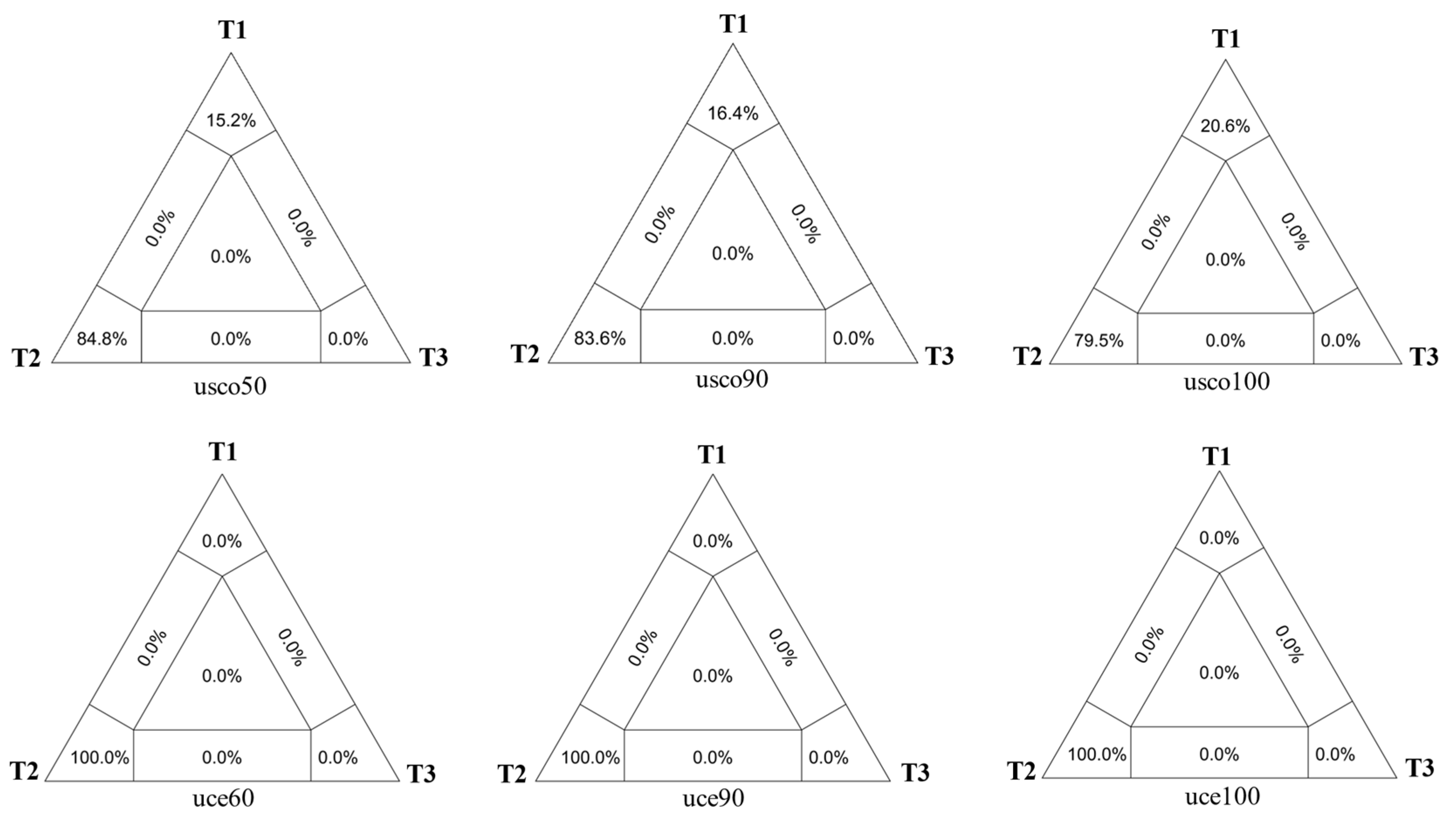
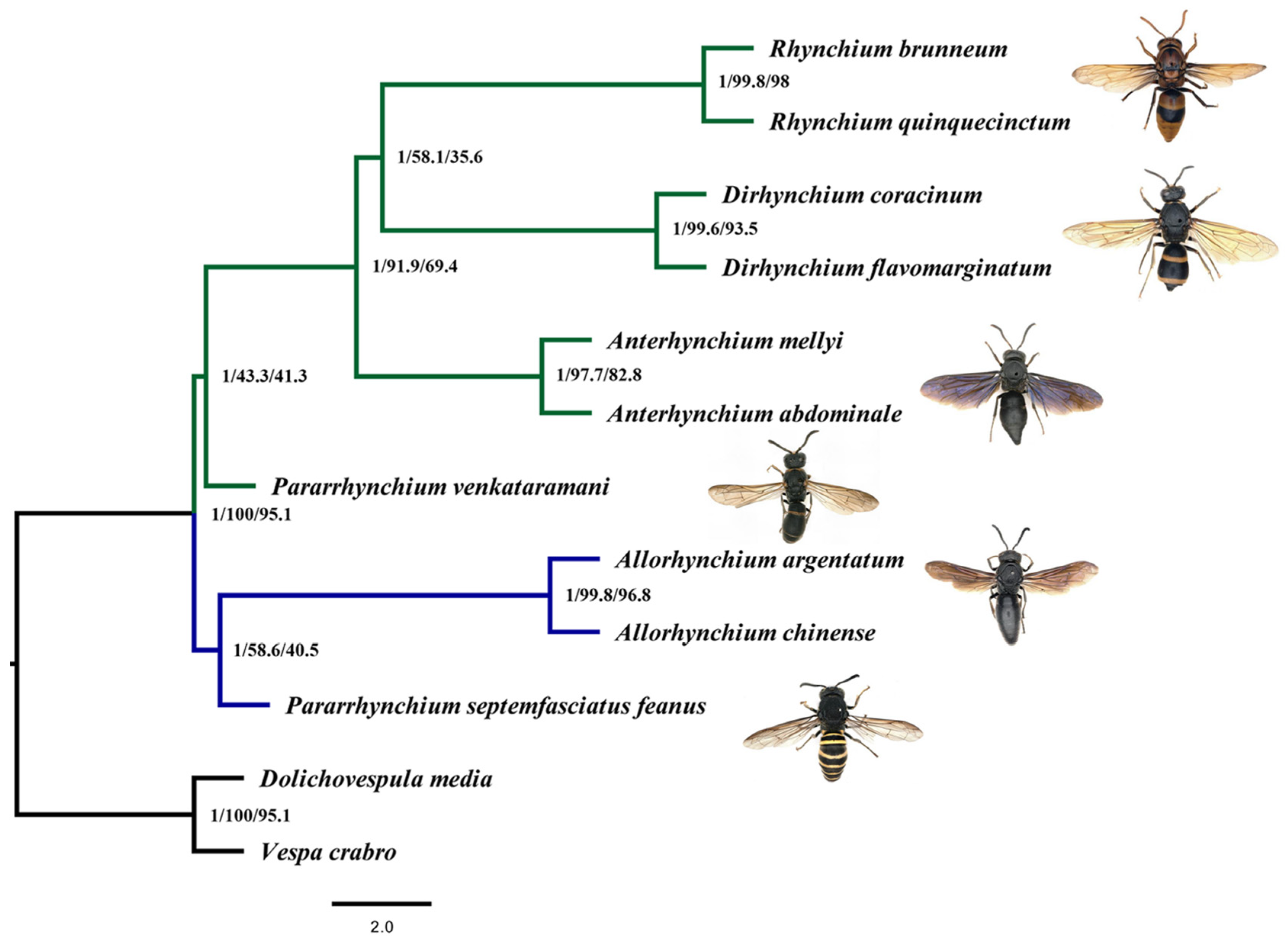
3.2. Phylogenetic Relationships among the Rhynchium Genera Group
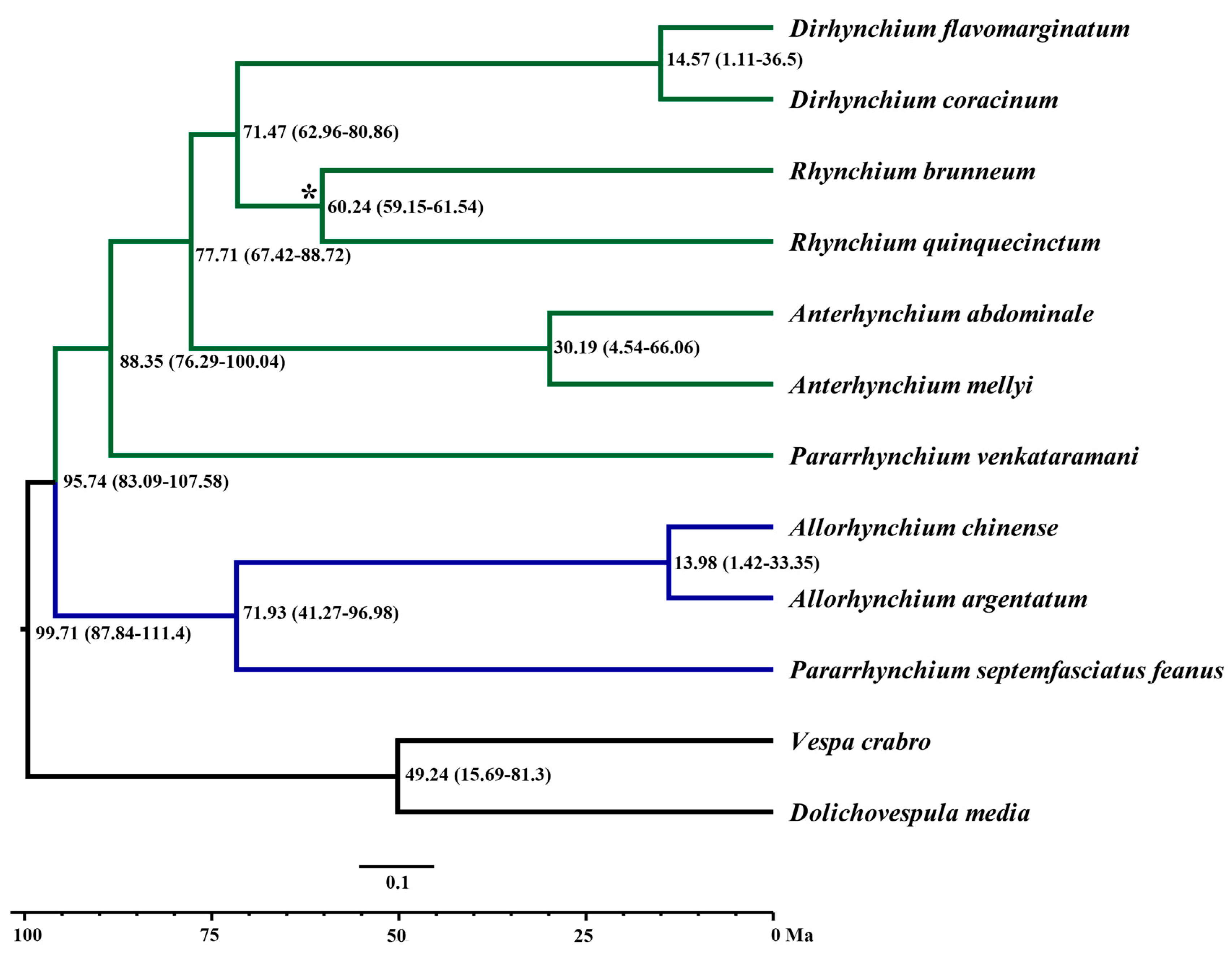
3.3. Molecular Dating of the Rhynchium Genera Group
4. Discussion
4.1. Sources of Gene Tree Discordance
4.2. Phylogenetic Relationships
4.3. Effects of Continental Drifts on the Distribution of Rhynchium Genera Group
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jennings, D.T.; Houseweart, M.W. Predation by Eumenid Wasps (Hymenoptera: Eumenidae) on Spruce Budworm (Lepidoptera: Tortricidae) and Other Lepidopterous Larvae in Spruce-Fir Forests of Maine. Ann. Èntomol. Soc. Am. 1984, 49, 39–45. [Google Scholar] [CrossRef]
- Cowan, D.P. The Social Biology of Wasps; Cornell University Press: Ithaca, NY, USA, 1991; pp. 33–73. [Google Scholar]
- Carpenter, J.M.; Gusenleitner, J.; Madl, M. A catalogue of the Eumeninae (Hymenoptera: Vespidae) of the Ethiopian Region excluding Malagasy Subregion. Part I: Introduction, Key to Genera, Genera Aethiopicodynerus Gusenleitner 1997 to Cyrtolabulus van der Vecht 1969. Linz. Biol. Beitr. 2009, 41, 513–638. [Google Scholar]
- Carpenter, J.M.; Gusenleitner, J.; Madl, M. A Catalogue of the Eumeninae (Hymenoptera: Vespidae) of the Ethiopian Region excluding Malagasy Subregion. Part II: Genera Delta de Saussure 1885 to Zethus Fabricius 1804 and Species Incertae Sedis. Linz. Biol. Beitr. 2010, 42, 95–315. [Google Scholar]
- Tan, J.L.; Carpenter, J.M.; Achterberg, C. An illustrated key to the genera of Eumeninae from China, with a checklist of species (Hymenoptera, Vespidae). ZooKeys 2018, 740, 109–149. [Google Scholar] [CrossRef] [PubMed]
- Girish Kumar, P.; Sharma, G. A review of the genus Allorhynchium van der vecht, 1963 (Hymenoptera: Vespidae: Eumeninae) from the Indian subcontinent. Prommalia 2015, 3, 20–34. [Google Scholar]
- Luo, L.; Zhang, Q.H.; Chen, B.; Li, T.J. Additions to the knowledge of the genus Allorhynchium van der Vecht, 1963 from China (Hymenoptera, Vespidae, Eumeninae). J. Hymenopt. Res. 2020, 77, 119–137. [Google Scholar] [CrossRef]
- Kim, J.K. Taxonomic review of the genus Anterhynchium Saussure (Eumeninae, Vespidae, Hymenoptera) from East Asia. Korean J. Entomol. 2003, 33, 105–117. [Google Scholar] [CrossRef]
- Kumar, P.G. A taxonomic study on the genus Anterhynchium de Saussure (Hymenoptera: Vespidae: Eumeninae) from Indian subcontinent. Rec. Zool. Surv. India 2013, 113, 139–158. [Google Scholar]
- Selis, M. The genus Anterhynchium in the Philippine Islands (Hymenoptera: Vespidae, Eumeninae). Fragm. Entomol. 2017, 49, 175–178. [Google Scholar] [CrossRef]
- van der Vecht, J. Studies on Indo–Australian and East Asiatic Eumenidae (Hymenoptera: Vespoidea). Zool. Verh. 1963, 60, 3–116. [Google Scholar]
- Li, T.J.; Chen, B. Descriptions of four new species of Pararrhynchium de Saussure (Hymenoptera: Vespidae: Eumeninae) from China, with one newly recorded species and a key to Chinese species. Orient. Insects 2017, 52, 175–189. [Google Scholar] [CrossRef]
- Kumar, P.G.; Carpenter, J.M.; Kishore, L. A review of the genus Pararrhynchium de Saussure (Hymenoptera: Vespidae: Eumeninae) from India with the description of a new species. Halteres 2017, 8, 85–91. [Google Scholar] [CrossRef]
- Kumar, P.G.; Sharma, G. A taxonomic study on the genus Rhynchium Spinola (Hymenoptera: Vespidae: Eumeninae) from the India subcontinent. Rec. Zool. Surv. India 2013, 113, 105–122. [Google Scholar]
- Pham, P.H.; Kumar, P.G. Taxonomic notes on the genus Rhynchium Spinola (Hymenoptera: Vespidae: Eumeninae) from Vietnam, with a first checklist of the species worldwide. Ecol. Montenegrina 2016, 7, 530–537. [Google Scholar] [CrossRef]
- Luo, L.; Carpenter, J.M.; Chen, B.; Li, T.J. First Comprehensive Analysis of Both Mitochondrial Characteristics and Mitogenome-Based Phylogenetics in the Subfamily Eumeninae (Hymenoptera: Vespidae). Insects 2022, 13, 529. [Google Scholar] [CrossRef] [PubMed]
- Selis, M.; Carpenter, J.M. Revision of the Afrotropical species of the genus Anterhynchium de Saussure (Hymenoptera: Vespidae: Eumeninae). Zootaxa 2023, 5233, 1–102. [Google Scholar] [CrossRef]
- Carpenter, J.M. Catalogue of the Australian Eumeninae (Hymenoptera: Vespidae). Zootaxa 2021, 4919, 1–68. [Google Scholar] [CrossRef]
- Giordani Soika, A. Contributo alla conoscenza degli Eumenidi dell’Australia e della Nuova Guinea (Hym. Vespoidea). Bolletino Mus. Civ. Stor. Nat. Venezia 1993, 42, 125–149. [Google Scholar] [CrossRef]
- Giordani Soika, A. Ricerche sistematiche su alcuni generi di eumenidi della regione orientale e della Papuasia (Hymenoptera, Vespoidea). Ann. Mus. Civ. Stor. Nat. Giacomo Doria 1994, 90, 1–384. [Google Scholar] [CrossRef]
- Li, T.J.; Chen, B. Two newly recorded genera Stenodyneriellus and Lissodynerus with three new species from China (Hymenoptera, Vespidae, Eumeninae). J. Hymenopt. Res. 2016, 49, 111–127. [Google Scholar] [CrossRef]
- Lemmon, E.M.; Lemmon, A.R. High-Throughput Genomic Data in Systematics and Phylogenetics. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 99–121. [Google Scholar] [CrossRef]
- Scornavacca, C.; Delsuc, F.; Galtier, N. Phylogenetics in the Genomic Era 2020, Open Access Book. Available online: https://hal.inria.fr/PGE (accessed on 25 February 2021).
- Derti, A.; Roth, F.P.; Roth, F.P.; Wu, C.T. Mammalian ultraconserved elements are strongly depleted among segmental duplications and copy number variants. Nat. Genet. 2006, 38, 1216–1220. [Google Scholar] [CrossRef] [PubMed]
- McCormack, J.E.; Faircloth, B.C.; Crawford, N.G.; Gowaty, P.A.; Brumfield, R.T.; Glenn, T.C. Ultraconserved elements are novel phylogenomic markers that resolve placental mammal phylogeny when combined with species-tree analysis. Genome Res. 2012, 22, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.Y.; Ding, Y.H.; Tihelka, E.; Chen, Y.C.; Hu, F.; Liu, M.Q.; Zhang, F. Phylogenomics of Elongate-Bodied Springtails Reveals Independent Transitions from Aboveground to Belowground Habitats in Deep Time. Syst. Biol. 2022, 71, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Niu, Z.Q.; Luo, A.R.; Orr, M.C.; Ferrari, R.R.; Jin, J.F.; Wu, Q.T.; Zhang, F.; Zhu, C.D. Testing the systematic status of Homalictus and Rostrohalictus with weakened cross-vein groups within Halictini (Hymenoptera: Halictidae) using low-coverage whole-genome sequencing. Insect Sci. 2022, 29, 1819–1833. [Google Scholar] [CrossRef] [PubMed]
- Parada, A.; Hanson, J.; D’Eiía, G. Ultraconserved Elements Improve the Resolution of Difficult Nodes within the Rapid Radiation of Neotropical Sigmodontine Rodents (Cricetidae: Sigmodontinae). Syst. Biol. 2021, 70, 1090–1100. [Google Scholar] [CrossRef]
- Timilsena, P.R.; Wafula, E.K.; Barrett, C.F.; Ayyampalayam, S.; McNeal, J.R.; Rentsch, J.D.; McKain, M.R.; Heyduk, K.; Harkess, A.; Villegente, M.; et al. Phylogenomic resolution of order- and family-level monocot relationships using 602 single-copy nuclear genes and 1375 BUSCO genes. Front. Plant Sci. 2022, 13, 876779. [Google Scholar] [CrossRef]
- Peters, R.S.; Krogmann, L.; Mayer, C.; Donath, A.; Gunkel, S.; Meusemann, K.; Kozlov, A.; Podsiadlowski, L.; Petersen, M.; Lanfear, R.; et al. Evolutionary History of the Hymenoptera. Curr. Biol. 2017, 27, 1013–1018. [Google Scholar] [CrossRef]
- Bushnell, B. BBtools. 2014. Available online: https://sourceforge.net/projects/bbmap/ (accessed on 25 February 2022).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Xu, M.; Guo, L.; Gu, S.; Wang, O.; Zhang, R.; Peters, B.A.; Fan, G.; Liu, X.; Xu, X.; Deng, L.; et al. TGS-GapCloser: A fast and accurate gap closer for large genomes with low coverage of error-prone long reads. Gigascience 2020, 9, giaa094. [Google Scholar] [CrossRef]
- Waterhouse, R.M.; Seppey, M.; Simao, F.A.; Manni, M.; Ioannidis, P.; Klioutchnikov, G.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO Applications from Quality Assessments to Gene Prediction and Phylogenomics. Mol. Biol. Evol. 2018, 35, 543–548. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): Selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef]
- Naser-Khdour, S.; Minh, B.Q.; Zhang, W.; Stone, E.A.; Lanfear, R. The prevalence and impact of model violations in phylogenetic analysis. Genome Biol. Evol. 2019, 11, 3341–3352. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Kück, P.; Longo, G.C. FASconCAT-G: Extensive functions for multiple sequence alignment preparations concerning phylogenetic studies. Front. Zool. 2014, 11, 81. [Google Scholar] [CrossRef] [PubMed]
- Branstetter, M.G.; Longino, J.T.; Ward, P.S.; Faircloth, B.C. Enriching the ant tree of life: Enhanced UCE bait set for genome-scale phylogenetics of ants and other Hymenoptera. Methods Ecol. Evol. 2017, 8, 768–776. [Google Scholar] [CrossRef]
- Faircloth, B.C. PHYLUCE is a software package for the analysis of conserved genomic loci. Bioinformatics 2016, 32, 786–788. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods. 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Kainer, D.; Mayer, C.; Stamatakis, A. Selecting optimal partitioning schemes for phylogenomic datasets. BMC Evol. Biol. 2014, 14, 82. [Google Scholar] [CrossRef]
- Crotty, S.M.; Minh, B.Q.; Bean, N.G.; Holland, B.R.; Tuke, J.; Jermiin, L.S.; von Haeseler, A. GHOST: Recovering historical signal from heterotachously-evolved sequence alignments. Syst. Biol. 2020, 69, 249–264. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum–likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Zhang, C.; Rabiee, M.; Sayyari, E.; Mirarab, S. ASTRAL-III: Polynomial time species tree reconstruction from partially resolved gene trees. BMC Bioinform. 2018, 19, 153. [Google Scholar] [CrossRef] [PubMed]
- Sayyari, E.; Mirarab, S. Fast coalescent-based computation of local branch support from quartet frequencies. Mol. Biol. Evol. 2016, 33, 1654–1668. [Google Scholar] [CrossRef] [PubMed]
- Salichos, L.; Rokas, A. Inferring ancient divergences requires genes with strong phylogenetic signals. Nature 2013, 497, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.C.; Susko, E.; Roger, A.J. The relative importance of modeling site pattern heterogeneity versus partition-wise heterotachy in phylogenomic inference. Syst. Biol. 2019, 68, 1003–1019. [Google Scholar] [CrossRef]
- Wang, H.C.; Minh, B.Q.; Susko, E.; Roger, A.J. Modeling site heterogeneity with posterior mean site frequency profiles accelerates accurate phylogenomic estimation. Syst. Biol. 2018, 67, 216–235. [Google Scholar] [CrossRef]
- Molloy, E.K.; Warnow, T. To include or not to include: The impact of gene filtering on species tree estimation methods. Syst. Biol. 2018, 67, 285–303. [Google Scholar] [CrossRef]
- Strimmer, K.; von Haeseler, A. Likelihood-mapping: A simple method to visualize phylogenetic content of a sequence alignment. Proc. Natl. Acad. Sci. USA 1997, 94, 6815–6819. [Google Scholar] [CrossRef]
- Yang, Z. PAML 4: A program package for phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef]
- Piton, L. Paléontologie du Gisement Éocéne de Menat (Puy-de-Dôme) (Flore et Faune); Pascal and Francis: Clermont-Ferrand, France, 1940; pp. 1–303. [Google Scholar]
- dos Reis, M.; Yang, Z. Approximate Likelihood Calculation on a Phylogeny for Bayesian Estimation of Divergence Times. Mol. Biol. Evol. 2011, 28, 2161–2172. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.C. Microevolutionary processes generate phylogenomic discordance at ancient divergences. Evolution 2013, 67, 1823–1830. [Google Scholar] [CrossRef] [PubMed]
- Pollard, D.; Iyer, V.N.; Moses, A.M.; Eisen, M.B. Widespread discordance of gene trees with species tree in Drosophila: Evidence for incomplete lineage sorting. PLoS Genet. 2006, 2, e173. [Google Scholar] [CrossRef]
- Pag’es, M.; Bazin, E.; Galan, M.; Chaval, Y.; Claude, J.; Herbreteau, V.; Michaux, J.; Piry, S.; Morand, S.; Cosson, J.F. Cytonuclear discordance among Southeast Asian black rats (Rattus rattus complex). Mol. Ecol. 2013, 22, 1019–1034. [Google Scholar] [CrossRef]
- Ebersberger, I.; Galogoczy, P.; Taudien, S.; Taenzer, S.; Platzer, M.; von Haeseler, A. Mapping Human Genetic Ancestry. Mol. Biol. Evol. 2007, 24, 2266–2276. [Google Scholar] [CrossRef]
- Syring, J.; Farrell, K.; Businský, R.; Cronn, R.; Liston, A. Widespread Genealogical Nonmonophyly in Species of Pinus Subgenus Strobus. Syst. Biol. 2007, 56, 163–181. [Google Scholar] [CrossRef]
- Takahashi, K.; Terai, Y.; Nishida, M.; Okada, N. Phylogenetic relationships and ancient incomplete lineage sorting among cichlid fishes in Lake Tanganyika as revealed by analysis of the insertion of retroposons. Mol. Biol. Evol. 2001, 18, 2057–2066. [Google Scholar] [CrossRef]
- Jennings, W.B.; Edwards, S.V. Speciational history of Australian grassfinches (Poephila) inferred from thirty gene trees. Evolution 2005, 59, 2033–2047. [Google Scholar] [CrossRef]
- Carstens, B.C.; Knowles, L.L. Estimating species phylogeny from gene-tree probabilities despite incomplete lineage sorting: An example from Melanoplus grasshoppers. Syst. Biol. 2007, 56, 400–411. [Google Scholar] [CrossRef]
- Bossert, S.; Murray, E.A.; Almeida, E.A.B.; Brady, S.G.; Blaimer, B.B.; Danforth, B.N. Combining transcriptomes and ultraconserved elements to illuminate the phylogeny of Apidae. Mol. Phylogenet. Evol. 2019, 130, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Branstetter, M.G.; Müller, A.; Griswold, T.L.; Orr, M.C.; Zhu, C.D. Ultraconserved element phylogenomics and biogeography of the agriculturally important mason bee subgenus Osmia (Osmia). Syst. Entomol. 2021, 46, 453–472. [Google Scholar] [CrossRef]
- Pisanty, G.; Richter, R.; Martin, T.; Dettman, J.; Cardinal, S. Molecular phylogeny, historical biogeography and revised classification of andrenine bees (Hymenoptera: Andrenidae). Mol. Phylogenet. Evol. 2021, 170, 107151. [Google Scholar] [CrossRef]
- Cox, C.B.; Ladle, R.J.; Moore, P.D. Biogeography, an Ecological and Evolutionary Approach, 10th ed.; Science Ecology & Organismal Biology; John Wiley & Sons: Hoboken, NJ, USA, 2019. [Google Scholar]
- Wright, N.M.; Seton, M.; Williams, S.E.; Müller, R.D. The Late Cretaceous to recent tectonic history of the Pacific Ocean basin. Earth-Sci. Rev. 2016, 154, 138–173. [Google Scholar] [CrossRef]
- Willis, B. Principles of paleogeography. Science 1910, 31, 241–260. [Google Scholar] [CrossRef] [PubMed]
- Saito, F.; Kojima, J. Taxonomy and biogeography of Australian species of the Ropalidia stigma group and R. variegata group (Hymenoptera: Vespidae). Entomol. Sci. 2005, 8, 179–188. [Google Scholar] [CrossRef]
- Luo, L.; Huang, P.; Chen, B.; Li, T.J. Mitochondrial composition of and diffusion limiting factors of three social wasp genera Polistes, Ropalidia, and parapolybia (Hymenoptera: Vespidae). BMC Ecol. Evol. 2022, 22, 63. [Google Scholar] [CrossRef]
- Heine, C.; Zoethout, J.; Müller, R.D. Kinematics of the South Atlantic rift. Solid Earth 2013, 4, 215–253. [Google Scholar] [CrossRef]
- Perez-Diaz, L.; Eagles, G. Constraining South Atlantic growth with seafloor spreading data. Tectonics 2014, 33, 1848–1873. [Google Scholar] [CrossRef]
- Granot, R.; Dyment, J. The Cretaceous opening of the South Atlantic Ocean. Earth Planet. Sci. Lett. 2015, 414, 156–163. [Google Scholar] [CrossRef]
- Tominaga, M.; Sager, W.W.; Tivey, M.A.; Lee, S.M. Deep-tow magnetic anomaly study of the Pacific Jurassic Quiet Zone and implications for the geomagnetic polarity reversal timescale and geomagnetic field behavior. Solid Earth 2008, 113, B07710. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Accession Number | Average Read Coverage (X) | BUSCO Completeness (%) | The Number of Scaffolds | Assembly Size (Mb) | Max Read Length (kb) | N50 Scaffold (kb) | GC (%) |
|---|---|---|---|---|---|---|---|---|
| Rhynchium brunneum | SAMN36845277 | 78.85 | 94.6 | 42,348 | 194.79 | 1170.58 | 134.32 | 37.24 |
| Rhynchium quinquecinctum | SAMN36845333 | 78.26 | 94.0 | 48,619 | 196.27 | 1281.6 | 105.73 | 37.24 |
| Allorhynchium argentatum | SAMN36845335 | 84.95 | 95.3 | 49,682 | 180.81 | 894.85 | 160.54 | 37.27 |
| Allorhynchium chinense | SAMN36845336 | 72.63 | 91.5 | 187,058 | 211.49 | 318.76 | 23.39 | 36.31 |
| Anterhynchium (A.) abdominale | SAMN36845337 | 83.57 | 94.0 | 60,405 | 183.79 | 586.96 | 63.68 | 38.61 |
| Anterhynchium (A.) mellyi | SAMN36845339 | 90.69 | 94.7 | 23,487 | 169.37 | 842.5 | 112.22 | 38.66 |
| Anterhynchium (Dirhynchium) flavomarginatum | SAMN36845338 | 86.10 | 94.4 | 23,366 | 178.40 | 786.82 | 74.28 | 37.83 |
| Anterhynchium (Dirhynchium) coracinum | SAMN36845334 | 86.89 | 94.3 | 21,837 | 176.77 | 739.09 | 68.99 | 37.69 |
| Pararrhynchium septemfasciatus feanus | SAMN36845341 | 100.40 | 95.6 | 15,712 | 152.99 | 4723.7 | 670.26 | 38.53 |
| Pararrhynchium venkataramani | SAMN36845340 | 85.82 | 92.5 | 51,762 | 178.97 | 728.23 | 51.56 | 38.76 |
| Matrix | Average Missing Taxa per Locus (%) | Number of Loci | Number of Sites | Missing Sites (%) | Average Locus Length |
|---|---|---|---|---|---|
| USCO50 | 2.99 | 5714 | 3,467,928 | 3.19 | 606.92 |
| USCO90 | 1.05 | 5115 | 2,945,110 | 1.18 | 575.78 |
| USCO100 | 0 | 4398 | 2,423,609 | 0.01 | 551.07 |
| UCE60 | 2.56 | 2138 | 2,396,740 | 2.56 | 1121.02 |
| UCE90 | 0.77 | 1928 | 2,159,911 | 0.85 | 1120.29 |
| UCE100 | 0 | 1726 | 1,933,308 | 0.24 | 1120.11 |
| Matrix | Partitioning | GHOST | ASTRAL | PMSF |
|---|---|---|---|---|
| USCO50 | T1 | T1 | T2 | T1 |
| USCO90 | T1 | T1 | T2 | T1 |
| USCO100 | T1 | T1 | T1 | T1 |
| UCE60 | T1 | T2 | T2 | - |
| UCE90 | T1 | T2 | T2 | - |
| UCE100 | T1 | T2 | T2 | - |
| Topologies | logL | deltaL | bp-RELL | P-WKH | P-WSH | P-AU |
|---|---|---|---|---|---|---|
| T1 | −8,408,349.19 | 964.1 | 0 * | 0 * | 0 * | 2.88 × 10−6 |
| T2 | −8,407,385.093 | 0 * | 1 | 1 | 1 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, M.; He, S.-L.; Chen, B.; Li, T.-J. Phylogeny of Rhynchium and Its Related Genera (Hymenoptera: Eumeninae) Based on Universal Single-Copy Orthologs and Ultraconserved Elements. Insects 2023, 14, 775. https://doi.org/10.3390/insects14090775
Dai M, He S-L, Chen B, Li T-J. Phylogeny of Rhynchium and Its Related Genera (Hymenoptera: Eumeninae) Based on Universal Single-Copy Orthologs and Ultraconserved Elements. Insects. 2023; 14(9):775. https://doi.org/10.3390/insects14090775
Chicago/Turabian StyleDai, Min, Shu-Lin He, Bin Chen, and Ting-Jing Li. 2023. "Phylogeny of Rhynchium and Its Related Genera (Hymenoptera: Eumeninae) Based on Universal Single-Copy Orthologs and Ultraconserved Elements" Insects 14, no. 9: 775. https://doi.org/10.3390/insects14090775