

Effect of the Sterile Insect Technique and Augmentative Parasitoid Releases in a Fruit Fly Suppression Program in Mango-Producing Areas of Southeast Mexico

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
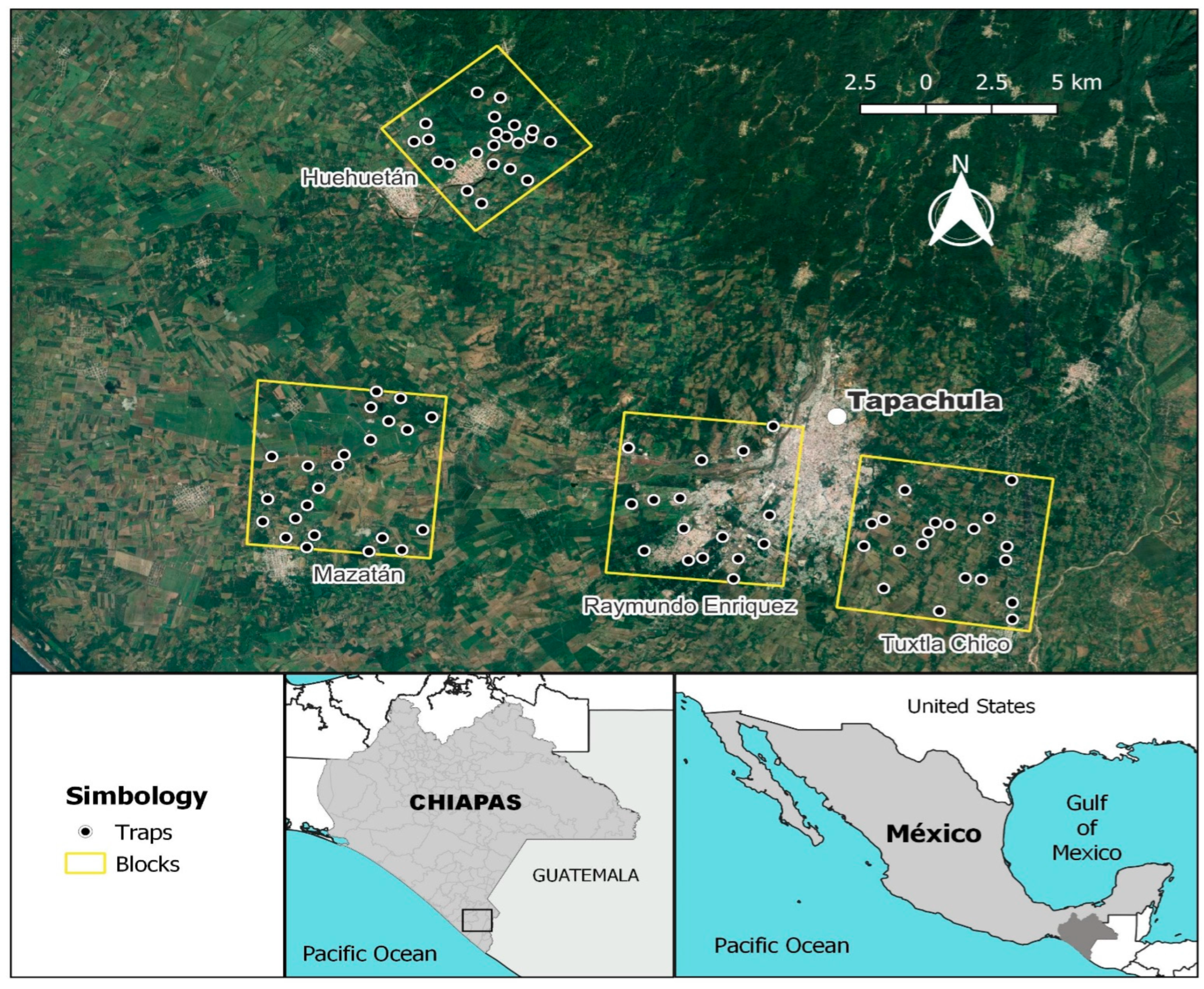
2.1. Study Areas
2.1.1. Raymundo Enriquez (ABC + SIT Treatment)
2.1.2. Mazatán (SIT Treatment)
2.1.3. Tuxtla Chico (ABC Treatment)
2.1.4. Huehuetán (Control Treatment)
2.2. Biological Material
2.3. Packing and Release of Insects
2.3.1. Parasitoids
2.3.2. Flies
2.4. Distribution of Insect Releases
2.5. Indicators of Parasitism and Fly Population Levels
2.5.1. Parasitism
2.5.2. Flies per Trap per Day (FTD)
2.6. Data Analysis
3. Results
3.1. Parasitism
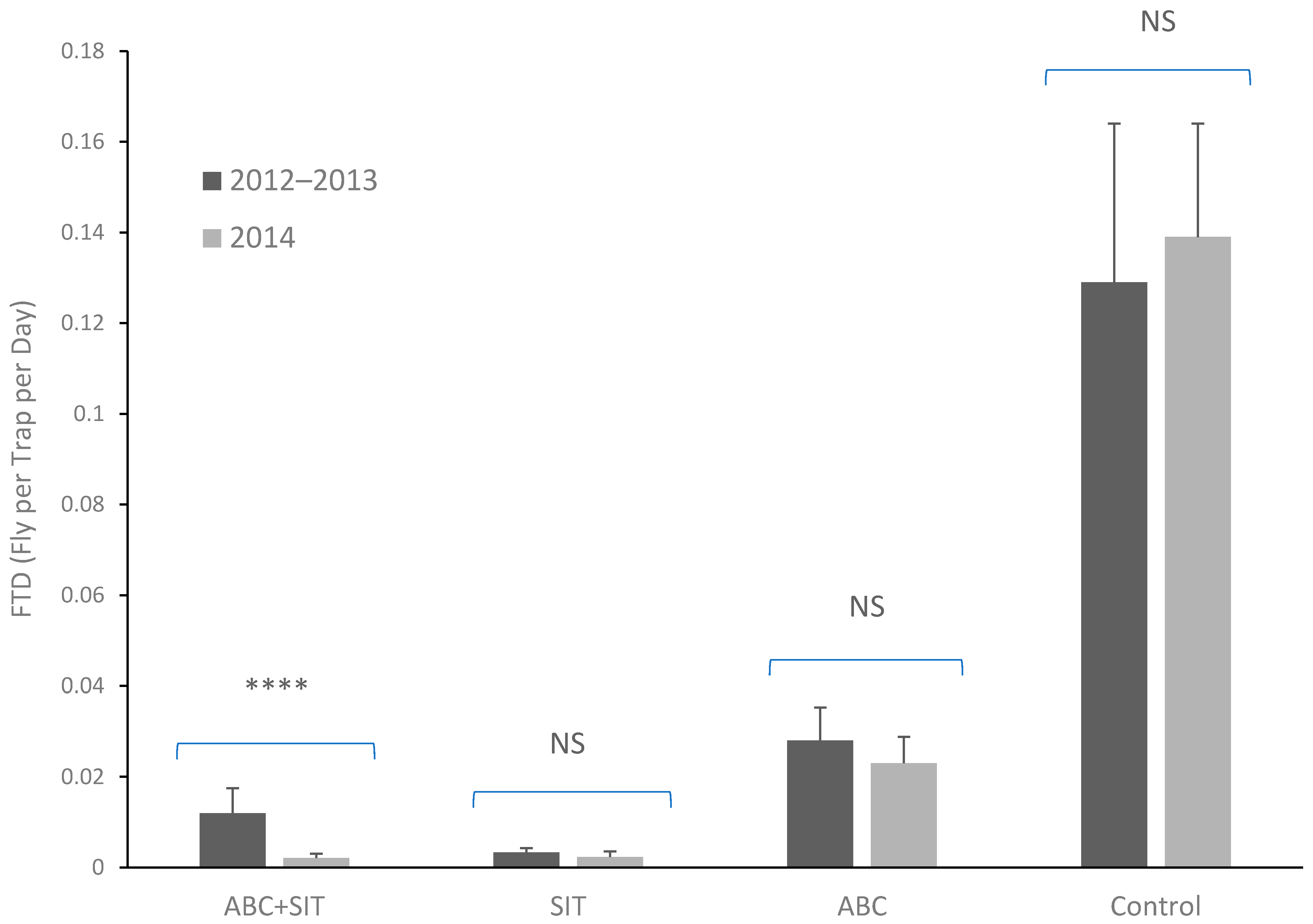
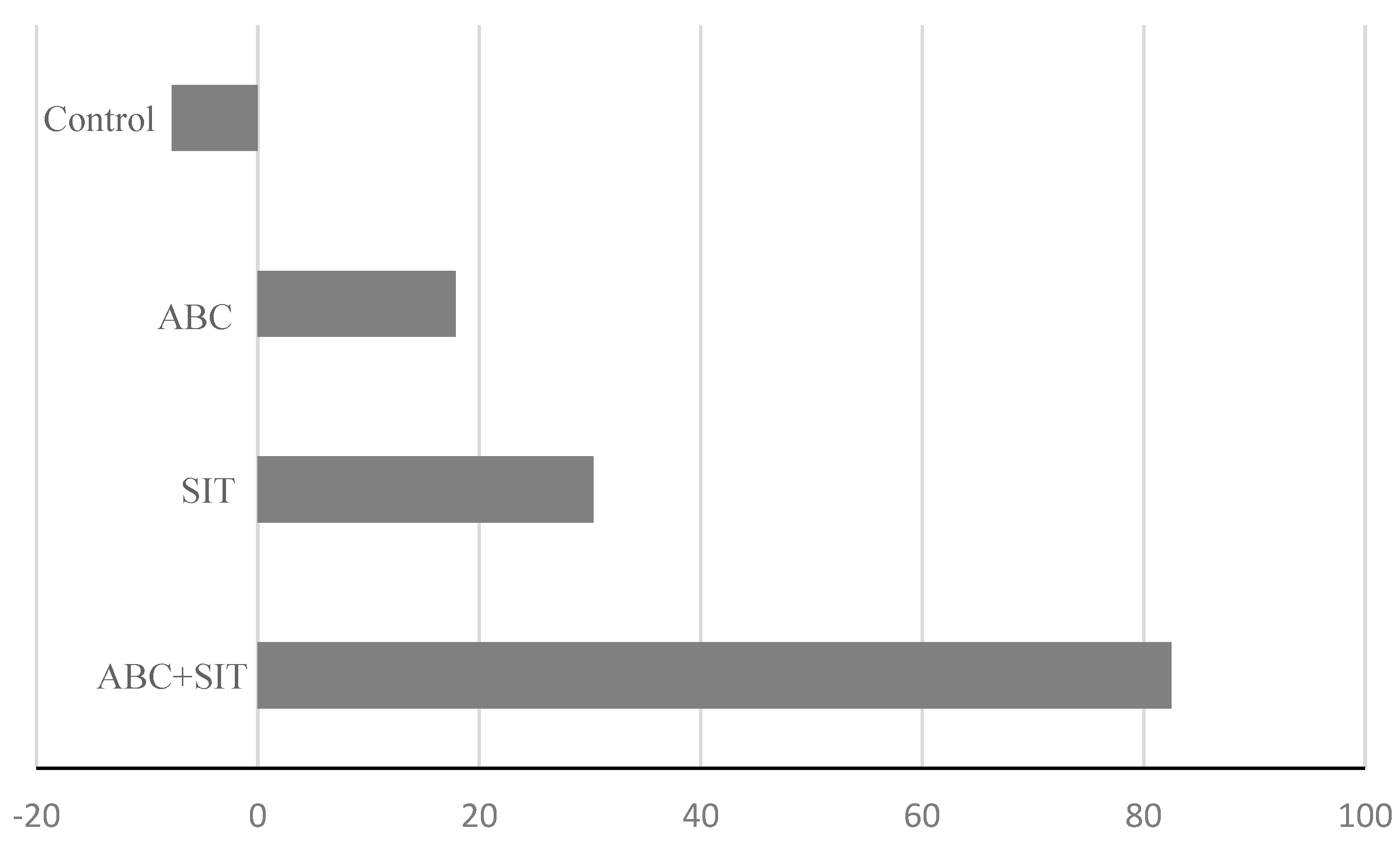
3.2. Flies per Trap per Day (FTD)
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bourtzis, K.; Vreysen, M.J.B. Sterile Insect Technique (SIT) and its applications. Insects 2021, 13, 638. [Google Scholar] [CrossRef]
- Wyss, J.H. Screwworm eradication in the Americas. Ann. N. Y. Acad. Sci. 2000, 916, 86–93. [Google Scholar] [CrossRef]
- Hendrichs, J.; Robinson, A. Sterile insect technique. In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé, R.T., Eds.; Academic Press: Burlington, NJ, USA, 2009; pp. 953–957. [Google Scholar] [CrossRef]
- Shelly, T.; McInnis, D. Sterile insect technique and control of Tephritid Fruit Flies: Do species with complex courtship require higher overflooding ratios? Ann. Entomol. Soc. Amer. 2016, 109, 1–11. [Google Scholar] [CrossRef]
- Enkerlin, W.R. Impact of fruit flies control programmes using the sterile insect technique. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 980–1002. [Google Scholar]
- Mastrangelo, T.; Kovaleski, A.; Maset, B.; Costa, M.L.Z.; Barros, C.; Lopes, L.A.; Caceres, C. Improvement of the mass-rearing protocols for the South American fruit fly for application of the sterile insect technique. Insects 2021, 9, 622. [Google Scholar] [CrossRef]
- Cáceres, C.; Hendrichs, J.; Vreysen, M. Development and improvement of rearing techniques for fruit flies (Diptera: Tephritidae) of economic importance. Int. J. Trop. Insect Sci. 2014, 34, S1–S12. [Google Scholar] [CrossRef]
- Garcia, F.R.M.; Ovruski, S.M.; Suárez, L.; Cancino, J.; Liburd, O.E. Biological control of tephritid fruit flies in the Americas and Hawaii: A Review of the Use of Parasitoids and Predators. Insects 2020, 11, 662. [Google Scholar] [CrossRef]
- Wong, T.T.Y.; Ramadan, M.M.; McInnis, D.O.; Mochizuki, N.; Nishimoto, J.I.; Herr, J.C. Augmentative releases of Diachasmimorpha tryoni (Hymenoptera: Braconidae) to suppress a Mediterranean fruit fly (Diptera: Tephritidae) population in Kula, Maui, Hawaii. Biol. Control 1991, 1, 2–7. [Google Scholar] [CrossRef]
- Montoya, P.; Liedo, P.; Benrey, B.; Cancino, J.; Barrera, J.F.; Sivinski, J.; Aluja, M. Biological control of Anastrepha spp. (Diptera: Tephritidae) in mango orchards through augmentative releases of Diachasmimorpha longicaudata (Ashmead) (Hymenoptera: Braconidae). Biol. Control 2000, 18, 216–224. [Google Scholar] [CrossRef]
- Sivinski, J.M.; Calkins, C.O.; Baranowski, R.; Harris, D.; Brambila, J.; Diaz, J.; Burns, R.E. Suppression of a Caribbean Fruit Fly (Anastrepha suspensa (Loew) Diptera: Tephritidae) population through augmented releases of the parasitoid Diachasmimorpha longicaudata (Ashmead) (Hymenoptera: Braconidae). Biol. Control 1996, 6, 177–185. [Google Scholar] [CrossRef]
- Bautista, R.C.; Mochizuki, N.; Spencer, J.P.; Harris, E.J.; Ichimura, D.M. Mass-rearing of the Tephritid fruit fly parasitoid Fopius arisanus (Hymenoptera: Braconidae). Biol. Control 1999, 15, 137–144. [Google Scholar] [CrossRef]
- Cancino, J.; Ruíz, L.; Viscarret, M.; Sivinski, J.; Hendrichs, J. Application of nuclear techniques to improve the mass production and management of fruit fly parasitoids. Insects 2012, 3, 1105–1125. [Google Scholar] [CrossRef]
- Marec, F.; Vreysen, M.J.B. Advances and challenges of using the sterile insect technique for the management of pest Lepidoptera. Insects 2019, 10, 371. [Google Scholar] [CrossRef]
- Bale, J.S.; van Lenteren, J.C.; Bigler, F. Biological control and sustainable food production. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 761–776. [Google Scholar] [CrossRef]
- Mangan, R.L.; Bouyer, J. Population suppression in support of the sterile insect technique. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 549–574. [Google Scholar]
- Bakhtiar, T.; Fitri, I.R.; Hanum, F.; Kusnanto, A. Mathematical model of pest control using different release rates of sterile insects and natural enemies. Mathematics 2022, 10, 883. [Google Scholar] [CrossRef]
- Barclay, H.J. Models for pest control: Complementary effects of periodic releases of sterile pests and parasitoids. Theor. Popul. Biol. 1987, 32, 76–89. [Google Scholar]
- Knipling, E.F. Principles of insect parasitism analyzed from new perspectives: Practical implications for regulating insect populations by biological means. In Agriculture Handbook No. 963; U.S. Department of Agriculture: Washington, DC, USA, 1992; p. 337. [Google Scholar]
- Liedo, P.; Montoya, P.; Toledo, J. Area-Wide management of fruit flies in a tropical mango growing area integrating the sterile insect technique and biological control: From a research to an operational programme. In Area-Wide Integrated Pest Management: Development and Field Application; Hendrichs, J., Pereira, R., Vreysen, M.J.B., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 197–214. [Google Scholar]
- Flores, S.; Gómez, E.; Campos, S.; Gálvez, F.; Toledo, J.; Liedo, P.; Pereira, R.; Montoya, P. Evaluation of mass trapping and bait stations to control Anastrepha (Diptera: Tephritidae) fruit flies in mango orchards of Chiapas, Mexico. Fla. Entomol. 2017, 100, 358–365. [Google Scholar] [CrossRef]
- Aluja, M.; Celedonio-Hurtado, H.; Liedo, P.; Cabrera, M.; Castillo, F.; Guillén, J.; Rios, E. Seasonal population fluctuations and ecological implications for management of Anastrepha fruit flies (Diptera: Tephritidae) in commercial mango orchards in Southern Mexico. J. Econ. Entomol. 1996, 89, 654–667. [Google Scholar] [CrossRef]
- Orozco-Dávila, D.; Artiaga-López, T.; Solís, E.; Hernández, E. Métodos de cría masiva. In Moscas de la Fruta, Fundamentos y Procedimientos Para su Manejo, 2nd ed.; Montoya, P., Toledo, J., Hernández, E., Eds.; S y G Editores: Ciudad de México, Mexico, 2020; pp. 377–397. [Google Scholar]
- Orozco-Dávila, D.; Quintero, L.; Hernández, E.; Solís, E.; Artiaga, T.; Hernández, R.; Ortega, C.; Montoya, P. Mass rearing and sterile insect releases for the control of Anastrepha spp. pests in Mexico—A review. Entomol. Exp. Appl. 2017, 164, 176–187. [Google Scholar] [CrossRef]
- Mubarqui, R.L.; Perez, R.C.; Kladt, R.A.; Lopez, J.L.Z.; Parker, A.; Seck, M.T.; Sall, B.; Bouyer, J. The smart aerial release machine, a universal system for applying the sterile insect technique. PLoS ONE 2014, 9, e103077. [Google Scholar] [CrossRef]
- Hernández, E.; Escobar, A.; Bravo, B.; Montoya, P. Chilled packing systems for fruit flies (Diptera: Tephritidae) in the sterile insect Technique. Neotrop. Entomol. 2010, 39, 601–607. [Google Scholar] [CrossRef]
- Montoya, P.; Cancino, J.; Zenil, M.; Santiago, G.; Gutiérrez, J.M. The augmentative biological control component in the Mexican campaign against Anastrepha spp. fruit flies. In Area-Wide Control of Insect Pests: From Research to Field Implementation, 1st ed.; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 661–670. ISBN 978-4020-6058-8. [Google Scholar]
- Montoya, P.; Cancino, J.; Ruiz, L. Packing of fruit fly parasitoids for augmentative releases. Insects 2012, 3, 889–899. [Google Scholar] [CrossRef]
- FAO/IAEA. Guideline for Packing, Shipping, Holding and Release of Sterile Flies in Area-Wide Fruit Fly Control Programmes, 2nd ed.; Zavala-López, J.L., Enkerlin, W.R., Eds.; FAO: Rome, Italy, 2017; p. 140. [Google Scholar]
- Contreras-Navarro, Y.; Luis-Alvarez, H.; García-Coapio, G.; Hernández, R.; Flores, S.; Montoya, P. Quality control traceability during the packing and release process of Ceratitis capitata for Sterile Insect Technique. J. Appl. Entomol. 2022, 146, 1252–1259. [Google Scholar] [CrossRef]
- Hernández-Ortiz, V. El Género Anastrepha Schiner en México. Taxonomía, Distribución y sus Plantas Huéspedes; Publicación # 33; Instituto de Ecología: Xalapa, Mexico, 1992; p. 167. [Google Scholar]
- Wharton, R.A.; Marsh, P.M. New world opiinae (hymenoptera: Braconidae) parasitic on Tephritidae (Diptera). J. Wash. Acad. Sci. 1978, 68, 147–167. [Google Scholar]
- Aluja, M.; Arredondo, J.; Díaz-Fleischer, F.; Birke, A.; Rull, J.; Niogret, J.; Epsky, N. Susceptibility of 15 Mango (Sapindales: Anacardiaceae) cultivars to the attack by Anastrepha ludens and Anastrepha obliqua (Diptera: Tephritidae) and the role of underdeveloped fruit as pest reservoirs: Management implications. J. Econ. Entomol. 2014, 107, 375–388. [Google Scholar] [CrossRef]
- Vreysen, M.J.B. Monitoring sterile and wild insect in area-wide integrated pest management programmes. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 485–528. [Google Scholar]
- Plá, I.; García de Oteyza, J.; Tur, C.; Martínez, M.A.; Laurín, M.C.; Alonso, E.; Martínez, M.; Martín, A.; Sanchis, R.; Navarro, M.C.; et al. Sterile insect technique programme against Mediterranean fruit fly in the Valencian Community (Spain). Insects 2021, 12, 415. [Google Scholar] [CrossRef]
- Enkerlin, W.; Gutiérrez-Ruelas, J.M.; Villaseñor-Cortes, A.; Cotoc-Roldan, E.; Midgarden, D.; Lira, E.; Zavala-López, J.L.; Hendrichs, J.; Liedo, P.; Trujillo-Arriaga, F.J. Area freedom in Mexico from Mediterranean fruit fly (Diptera: Tephritidae): A review of over 30 years of a successful containment program using an integrated area-wide SIT approach. Fla. Entomol. 2015, 98, 665–681. [Google Scholar] [CrossRef]
- Zavala-Lopez, J.L.; Marte-Diaz, G.; Martínez-Pujols, F. Successful area-wide eradication of the invading Medi-terranean fruit fly in the Dominican Republic. In Area-Wide Integrated Pest Management: Development and Field Application; Hendrichs, J., Pereira, R., Vreysen, M.J.B., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 519–537. [Google Scholar]
- Flores, S.; Montoya, P.; Toledo, J.; Enkerlin, W.; Liedo, P. Estimation of populations and sterility induction in Anastrepha ludens (Diptera: Tephritidae) fruit flies. J. Econ. Entomol. 2014, 107, 1502–1507. [Google Scholar] [CrossRef]
- Quintero-Fong, L.; Luis, J.H.; Montoya, P.; Orozco-Dávila, D. In situ sexual competition between sterile males of the genetic sexing Tapachula-7 strain and wild Anastrepha ludens flies. Crop Prot. 2018, 106, 1–5. [Google Scholar] [CrossRef]
- Flores, S.; Campos, S.; Gómez, E.; Espinoza, E.; Wilson, W.; Montoya, P. Evaluation of field dispersal and survival capacity of the genetic sexing strain Tapachula-7 of Anastrepha ludens (Diptera: Tephritidae). Fla. Entomol. 2015, 98, 209–214. [Google Scholar] [CrossRef]
- Flores, S.; Campos, S.; Gómez, E.; Leal-Mubarqui, R.; Morales-Marin, J.L.; Vélez, J.; Bello-Rivera, A.; Montoya, P. Performance of the Tap-7 genetic sexing strain used to control Anastrepha ludens populations in the citrus region of Tamaulipas, Mexico. In Area-Wide Management of Fruit Fly Pests; Pérez-Staples, D., Diaz-Fleischer, F., Montoya, P., Vera, M., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 177–189. [Google Scholar]
- Harbi, A.; de Pedro, L.; Ferrara, F.A.A.; Tormos, J.; Chermiti, B.; Beitia, F.; Sabater-Munoz, B. Diachasmimorpha longicaudata Parasitism response to medfly host fruit and fruit infestation age. Insects 2019, 10, 211. [Google Scholar] [CrossRef]
- Suarez, L.; Murua, F.; Lara, N.; Escobar, J.; Taret, G.; Rubio, J.; Nieuwenhove, G.; Bezdjian, L.; Schliserman, P.; Ovruski, S. Biological control of Ceratitis capitata (Diptera: Tephritidae) in Argentina: Releases of Diachasmimorpha longicaudata (Hymenoptera: Braconidae) in fruit-producing semi-arid areas of San Juan. Nat. Sci. 2014, 6, 664–675. [Google Scholar] [CrossRef]
- Isiordia-Aquino, N.; Robles-Bermudez, A.; Cambero-Campos, O.J.; Díaz-Heredia, M.; Gonzalez-Castellon, L.; Flores-Canales, R.J. Biological control of Anastrepha Schiner (Diptera: Tephritidae) through augmentative releases of Diachasmimorpha longicaudata Ashmead (Hymenoptera; Braconidae) in fruit-producing marginal areas of Northern Nayarit, México. Afr. J. Agric. Res. 2017, 12, 3331–3338. [Google Scholar] [CrossRef]
- Montoya, P.; López, P.; Cruz, J.; López, F.; Cadena, C.; Cancino, J.; Liedo, P. Effect of Diachasmimorpha longicaudata releases on the native parasitoid guild attacking Anastrepha spp. larvae in disturbed zones of Chiapas, Mexico. BioControl 2017, 62, 581–593. [Google Scholar] [CrossRef]
- Kindlmann, P.; Ameixa, O.M.C.C.; Dixon, A.F.G. Ecological effects of invasive alien species on native communities, with particular emphasis on the interactions between aphids and ladybirds. BioControl 2011, 56, 469–476. [Google Scholar] [CrossRef]
- Meirelles, R.N.; Rodrigues Redaelli, L.; Mundstock Jahnke, S.; Bernardes Ourique, C.; Branco Ozorio, V.D. Parasitism of fruit flies (Tephritidae) in field, after releases of Diachasmimorpha longicaudata (Ashmead) (Hymenoptera: Braconidae) in Rio Grande do Soul. Rev. Bras. Frutic. 2016, 38, e-673. [Google Scholar] [CrossRef]
- Vanoye-Eligio, V.; Vázquez-Sauceda, M.L.; Chacón-Hernández, J.L.; Rocandio-Rodríguez, M.; Rosas-Mejía, M.; Vanoye-Eligio, M. Insights into the population dynamics of Anastrepha obliqua and Anastrepha ludens in the Neotropical limits of Mexico. Entomol. Exp. Appl. 2023, 171, 35–44. [Google Scholar] [CrossRef]
- Sivinski, J.; Piñero, J.; Aluja, M. The distributions of parasitoids (Hymenoptera) of Anastrepha fruit flies (Diptera: Tephritidae) along an altitudinal gradient in Veracruz, Mexico. Biol. Control 2000, 18, 258–269. [Google Scholar] [CrossRef]
- Wong, T.; Ramadan, M.; Herr, J.; Mcinnis, D. Suppression of a Mediterranean fruit fly (Diptera: Tephritidae) population with concurrent parasitoid and sterile fly releases in Kula, Maui, Hawaii. J. Econ. Entomol. 1992, 85, 1671–1681. [Google Scholar] [CrossRef]
- Sivinski, J.; Jeronimo, F.; Holler, T. Development of aerial releases of Diachasmimorpha tryoni (Cameron) (Hymenoptera: Braconidae), a parasitoid that attacks the Mediterranean fruit fly, Ceratitis capitata (Weidemann) (Diptera: Tephritidae), in the Guatemalan highlands. Biocontrol Sci. Technol. 2000, 10, 15–25. [Google Scholar] [CrossRef]
- Montoya, P.; Flores-Sarmiento, E.; López, P.; Ayala, A.; Cancino, J. Additive effect of releasing sterile insects plus biocontrol agents against fruit fly pests (Diptera: Tephritidae) under confined conditions. Insects 2023, 14, 337. [Google Scholar]
- Leyva, J.L.; Browning, H.W.; Gilstrap, F.G. Effect of host fruit species, size, and color on parasitization of Anastrepha ludens (Diptera: Tephritidae) by Diachasmimorpha longicaudata (Hymenoptera: Braconidae). Environ. Entomol. 1991, 20, 1469–1474. [Google Scholar] [CrossRef]
- Ovruski, S.; Oroño, L.; Schliserman, P.; Nuñez-Campero, S. The effect of four fruit species on the parasitization rate of Anastrepha fraterculus (Diptera: Tephritidae, Trypetinae) by Diachasmimorpha longicaudata (Hymenoptera: Braconidae, Opiinae) under laboratory rearing conditions. Biocontrol Sci. Technol. 2007, 17, 1079–1085. [Google Scholar] [CrossRef]
- Sivinski, I. The influence of host fruit morphology on parasitization rates in the Caribbean fruit fly, Anastrepha suspensa. Entomophaga 1991, 36, 447–454. [Google Scholar] [CrossRef]
- Eben, A.; Benrey, B.; Sivinski, J.; Aluja, M. Host species and host plant effects on preference and performance of Diachasmimorpha longicaudata (Hymenoptera: Braconidae). Environ. Entomol. 2000, 29, 87–94. [Google Scholar] [CrossRef]
- Carrasco, M.; Montoya, P.; Cruz-Lopez, L.; Rojas, J.C. Response of the fruit fly parasitoid Diachasmimorpha longicaudata (Hymenoptera: Braconidae) to mango fruit volatiles. Environ. Entomol. 2005, 34, 576–583. [Google Scholar] [CrossRef]
- Paranhos, B.J.; Nava, D.E.; Malavasi, A. Biological control of fruit flies in Brazil. Pesqui. Agropecu. Bras. 2019, 54, e26037. [Google Scholar] [CrossRef]
- Marsaro Júnior, A.L.; Adaime, R.; Ronchi-Teles, B.; Lima, C.R.; Pereira, P.R.V.S. Anastrepha species (Diptera: Tephritidae), their hosts and parasitoids in the extreme north of Brazil. Biota Neotrop. 2011, 11, 4. [Google Scholar] [CrossRef]
- Kepner, J. Synergy: The big unknowns of pesticide exposure. Pestic. You 2004, 23, 17–20. [Google Scholar]
- Zhao, R.; Li, D.; Wang, X.; Li, Z.; Yu, X.; Shentu, X. Synergistic and Additive interactions of Zhongshengmycin to the chemical insecticide Pymetrozine for controlling Nilaparvata lugens (Hemiptera: Delphacidae). Front. Physiol. 2022, 13, 875610. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Host Fruits | Flies | Parasitoids | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Emergence | Emergence | Parasitism * | |||||||||||
| Species | NL + | L/F + | SpF + | NF+ | % | TF + | SpP + | NP + | % | TP + | |||
| ABC + SIT | Caimito, Chrysophyllum cainito (L.) | 127 | 10.71 | A. serpentina | 735 | 100 | 735 | D. longicaudata | 67 | 72.04 | 93 | 11.23 | |
| O. anastrephae | 26 | 27.96 | |||||||||||
| Guava, Psidium guajava (L.) | 2519 | 1 | A. striata | 1506 | 98.69 | 1526 | D. longicaudata | 424 | 99.76 | 425 | 21.78 | ||
| A. obliqua | 20 | 1.31 | A. pelleranoi | 1 | 0.24 | ||||||||
| Mamey, Mammea americana (L.) | 970 | 9.59 | A. serpentina | 5348 | 100 | 5348 | D. longicaudata | 340 | 98.55 | 345 | 6.06 | ||
| D. crawfordi | 5 | 1.45 | |||||||||||
| Mango var. Ataulfo, Mangifera indica L. | 232 | 0.18 | A. ludens | 14 | 73.68 | 19 | D. longicaudata | 6 | 100 | 6 | 24 | ||
| A. obliqua | 5 | 26.32 | |||||||||||
| Creole mango, Mangifera indica L. | 4207 | 1.3 | A. ludens | 437 | 20.26 | 2157 | D. longicaudata | 1479 | 99.60 | 1485 | 40.77 | ||
| A. obliqua | 1720 | 79.74 | D. areolatus | 3 | 0.20 | ||||||||
| U. anastrephae | 3 | 0.20 | |||||||||||
| Sour orange, Citrus aurantium L. | 640 | 2.1 | A. ludens | 775 | 100 | 775 | D. longicaudata | 78 | 100 | 78 | 9.14 | ||
| Hog plums, Spondias mombin L. | 2018 | 2.15 | A. obliqua | 1378 | 100 | 1378 | D. longicaudata | 1767 | 90.94 | 1943 | 58.51 | ||
| D. areolatus | 66 | 3.40 | |||||||||||
| O. anastrephae | 7 | 0.36 | |||||||||||
| U. anastrephae | 103 | 5.30 | |||||||||||
| Mean | 3.86 ± 1.64 a | 100 | 24.49 ± 7.20 a | ||||||||||
| SIT | Grapefruit, Citrus paradisi Mcfad | 160 | 0.46 | A. ludens | 50 | 100 | 50 | D. longicaudata | 4 | 100 | 4 | 7.41 | |
| Guava, Psidium guajava (L.) | 685 | 1.33 | A. striata | 765 | 100 | 765 | D. longicaudata | 10 | 100 | 10 | 1.29 | ||
| Creole mango, Mangifera indica L. | 162 | 0.01 | A. obliqua | 1 | 100 | 1 | |||||||
| Sour orange, Citrus aurantium L. | 760 | 0.95 | A. ludens | 477 | 100 | 477 | D. longicaudata | 7 | 100 | 7 | 1.45 | ||
| Mean | 0.68 ± 0.28 a | 2.53 ± 1.65 b | |||||||||||
| ABC | Grapefruit, Citrus paradisi Mcfad | 80 | 0.74 | A. ludens | 39 | 100 | 39 | ||||||
| Guava, Psidium guajava (L.) | 33 | 1.45 | A. striata | 17 | 100 | 17 | D. longicaudata | 4 | 80 | 5 | 22.73 | ||
| D. areolatus | 1 | 20 | |||||||||||
| Mamey, Mammea americana (L.) | 4 | 21 | A.serpentina | 67 | 100 | 67 | D. longicaudata | 7 | 100 | 7 | 9.46 | ||
| Mango var. Ataulfo, Mangifera indica L. | 744 | 3.32 | A. ludens | 680 | 99.85 | 681 | D. longicaudata | 417 | 100 | 417 | 37.98 | ||
| A. obliqua | 1 | 0.15 | |||||||||||
| ABC | Creole mango, Mangifera indica L. | 1141 | 2.77 | A. ludens | 423 | 54.94 | 770 | D. longicaudata | 395 | 89.16 | 443 | 36.52 | |
| A. obliqua | 347 | 45.06 | D. areolatus | 48 | 10.84 | ||||||||
| Sour orange, Citrus aurantium L. | 26 | 4.69 | A. ludens | 88 | 100 | 88 | D. longicaudata | 9 | 90 | 10 | 10.2 | ||
| D. crawfordi | 1 | 10 | |||||||||||
| Hog plums, Spondias mombin L. | 176 | 3.15 | A. obliqua | 427 | 100 | 427 | D. longicaudata | 65 | 65.66 | 99 | 18.82 | ||
| D. areolatus | 32 | 32.32 | |||||||||||
| A. pelleranoi | 2 | 2.02 | |||||||||||
| Mean | 5.3 ± 2.66 a | 19.38 ± 5.36 a | |||||||||||
| Control | Creole mango, Mangifera indica L. | 1371 | 1.39 | A. ludens | 46 | 4.23 | 1088 | D. longicaudata | 13 | 100 | 13 | 1.18 | |
| A. obliqua | 1042 | 95.77 | |||||||||||
| Bitter orange, Citrus aurantium L. | 20 | 1.25 | A. ludens | 10 | 100 | 10 | |||||||
| Mean | 1.32 ± 0.07 a | 0.59 ± 0.59 b | |||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cancino, J.; Montoya, P.; Gálvez, F.O.; Gálvez, C.; Liedo, P. Effect of the Sterile Insect Technique and Augmentative Parasitoid Releases in a Fruit Fly Suppression Program in Mango-Producing Areas of Southeast Mexico. Insects 2023, 14, 719. https://doi.org/10.3390/insects14090719
Cancino J, Montoya P, Gálvez FO, Gálvez C, Liedo P. Effect of the Sterile Insect Technique and Augmentative Parasitoid Releases in a Fruit Fly Suppression Program in Mango-Producing Areas of Southeast Mexico. Insects. 2023; 14(9):719. https://doi.org/10.3390/insects14090719
Chicago/Turabian StyleCancino, Jorge, Pablo Montoya, Fredy Orlando Gálvez, Cesar Gálvez, and Pablo Liedo. 2023. "Effect of the Sterile Insect Technique and Augmentative Parasitoid Releases in a Fruit Fly Suppression Program in Mango-Producing Areas of Southeast Mexico" Insects 14, no. 9: 719. https://doi.org/10.3390/insects14090719