
Combining Irradiation and Biological Control against Brown Marmorated Stink Bug: Are Sterile Eggs a Suitable Substrate for the Egg Parasitoid Trissolcus japonicus?

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Rearing
2.2. Egg Sterilization
- (a)
- Irradiation of fresh eggs with gamma rays (irradiated eggs).
- (b)
- Eggs obtained by mating BMSB fertile females with irradiated males (i.e., SIT eggs).
- (c)
- Refrigeration of fresh BMSB eggs at 8 °C for a minimum of 14 days (refrigerated eggs, control).
2.2.1. Irradiation
2.2.2. Egg Refrigeration
2.3. Egg Dose Response
2.4. Male Dose Response
2.5. Evaluation of Sterile Eggs as an Oviposition Substrate
- (a)
- The percentage of parasitoids that emerged on the total number of eggs.
- (b)
- The sex ratio of the emerged parasitoids.
- (c)
- The longevity of the emerged parasitoid females.
- (d)
- The fecundity of the parasitoid females at 0 and 12 days after emergence.
- (e)
- The female dry weight (in the F1 generation).
2.6. Statistical Methods
3. Results
3.1. Egg Dose Response
3.2. Male Dose Response
3.3. Evaluation of Sterile Eggs as an Oviposition Substrate
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, D.-H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Entomol. 2013, 42, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Costi, E.; Haye, T.; Maistrello, L. Biological parameters of the invasive brown marmorated stink bug, Halyomorpha halys, in southern Europe. J. Pest Sci. 2017, 90, 1059–1067. [Google Scholar] [CrossRef]
- Leskey, T.C.; Nielsen, A.L. Impact of the invasive brown marmorated stink bug in North America and Europe: History, Biology, Ecology, and Management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musolin, D.L.; Konjević, A.; Karpun, N.N.; Protsenko, V.Y.; Ayba, L.Y.; Saulich, A.K. Invasive brown marmorated stink bug Halyomorpha halys (Stål) (Heteroptera: Pentatomidae) in Russia, Abkhazia, and Serbia: History of invasion, range expansion, early stages of establishment, and first records of damage to local crops. Arthropod-Plant Interact. 2018, 12, 517–529. [Google Scholar] [CrossRef]
- Kriticos, D.J.; Kean, J.M.; Phillips, C.B.; Senay, S.D.; Acosta, H.; Haye, T. The potential global distribution of the brown marmorated stink bug, Halyomorpha halys, a critical threat to plant biosecurity. J. Pest Sci. 2017, 90, 1033–1043. [Google Scholar] [CrossRef]
- Faúndez, E.I.; Rider, D.A. The brown marmorated stink bug Halyomorpha halys (Stål, 1855) (Heteroptera: Pentatomidae) in Chile. Arq. Entomolóxicos 2017, 17, 305–307. [Google Scholar]
- Bergmann, E.J.; Venugopal, D.; Martinson, H.; Raupp, M.J.; Shrewsbury, P.M. Host Plant Use by the invasive Halyomorpha halys (Stål) on woody ornamental trees and shrubs. PLOS ONE 2016, 11, e0149975. [Google Scholar] [CrossRef]
- Kuhar, T.P.; Kamminga, K.; Wallingford, A.; Philips, C.; Aigner, J.; Doughty, H. Evaluation of foliar insecticides for the control of brown marmorated stink bug in tomatoes in Virginia. Arthropod Manag. Tests 2012, 37, 1. [Google Scholar] [CrossRef] [Green Version]
- Kuhar, T.P.; Kamminga, K. Review of the chemical control research on Halyomorpha halys in the USA. J. Pest Sci. 2017, 90, 1021–1031. [Google Scholar] [CrossRef]
- Leskey, T.C.; Short, B.D.; Lee, D.-H. Efficacy of insecticide residues on adult Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) mortality and injury in apple and peach orchards: Residual insecticide efficacy on Halyomorpha halys. Pest Manag. Sci. 2013, 70, 1097–1104. [Google Scholar] [CrossRef]
- Leskey, T.C.; Hamilton, G.C.; Nielsen, A.L.; Polk, D.F.; Rodriguez-Saona, C.; Bergh, J.C.; Herbert, D.A.; Kuhar, T.P.; Pfeiffer, D.; Dively, G.P.; et al. Pest status of the brown marmorated stink bug, Halyomorpha halys in the USA. Outlooks Pest Manag. 2012, 23, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Leskey, T.C.; Short, B.D.; Butler, B.R.; Wright, S.E. Impact of the invasive brown marmorated stink bug, Halyomorpha halys (Stål), in Mid-Atlantic tree fruit orchards in the United States: Case studies of commercial management. Psyche: A J. Entomol. 2012, 2012, 535062. [Google Scholar] [CrossRef] [Green Version]
- Conti, E.; Avila, G.; Barratt, B.; Cingolani, F.; Colazza, S.; Guarino, S.; Hoelmer, K.; Laumann, R.A.; Maistrello, L.; Martel, G.; et al. Biological control of invasive stink bugs: Review of global state and future prospects. Entomol. Exp. Et Appl. 2021, 169, 28–51. [Google Scholar] [CrossRef]
- Abram, P.K.; Hoelmer, K.A.; Acebes-Doria, A.; Andrews, H.; Beers, E.H.; Bergh, J.C.; Bessin, R.; Biddinger, D.; Botch, P.; Buffington, M.L.; et al. Indigenous arthropod natural enemies of the invasive brown marmorated stink bug in North America and Europe. J. Pest Sci. 2017, 90, 1009–1020. [Google Scholar] [CrossRef] [Green Version]
- Falagiarda, M.; Carnio, V.; Chiesa, S.G.; Pignalosa, A.; Anfora, G.; Angeli, G.; Ioriatti, C.; Mazzoni, V.; Schmidt, S.; Zapponi, L. Factors influencing short-term parasitoid establishment and efficacy for the biological control of Halyomorpha halys with the samurai wasp Trissolcus japonicus. Pest Manag. Sci. 2023, 79, 2397–2414. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, F.; Gariepy, T.; Mason, P.; Gillespie, D.; Talamas, E.; Haye, T. Seasonal parasitism and host specificity of Trissolcus japonicus in northern China. J. Pest Sci. 2017, 90, 1127–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedstrom, C.; Lowenstein, D.; Andrews, H.; Bai, B.; Wiman, N. Pentatomid host suitability and the discovery of introduced populations of Trissolcus japonicus in Oregon. J. Pest Sci. 2017, 90, 1169–1179. [Google Scholar] [CrossRef]
- Haye, T.; Moraglio, S.T.; Stahl, J.; Visentin, S.; Gregorio, T.; Tavella, L. Fundamental host range of Trissolcus japonicus in Europe. J. Pest Sci. 2020, 93, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Talamas, E.J.; Herlihy, M.V.; Dieckhoff, C.; Hoelmer, K.A.; Buffington, M.; Bon, M.-C.; Weber, D.C. Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae) emerges in North America. J. Hymenopt. Res. 2015, 43, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Stahl, J.; Tortorici, F.; Pontini, M.; Bon, M.-C.; Hoelmer, K.; Marazzi, C.; Tavella, L.; Haye, T. First discovery of adventive populations of Trissolcus japonicus in Europe. J. Pest Sci. 2019, 92, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Peverieri, G.S.; Talamas, E.; Bon, M.C.; Marianelli, L.; Bernardinelli, I.; Malossini, G.; Benvenuto, L.; Roversi, P.F.; Hoelmer, K. Two Asian egg parasitoids of Halyomorpha halys (Stål) (Hemiptera, Pentatomidae) emerge in northern Italy: Trissolcus mitsukurii (Ashmead) and Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae). J. Hymenopt. Res. 2018, 67, 37–53. [Google Scholar] [CrossRef]
- Jentsch, P. Expanding the range of the samurai wasp, Trissolcus japonicus, in New York orchards. Fruit Quart 2017, 25, 31–35. [Google Scholar]
- Lowenstein, D.M.; Andrews, H.; Hilton, R.J.; Kaiser, C.; Wiman, N.G. Establishment in an introduced range: Dispersal capacity and winter survival of Trissolcus japonicus, an adventive egg parasitoid. Insects 2019, 10, 443. [Google Scholar] [CrossRef] [Green Version]
- Klassen, W.; Curtis, C.F. History of the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 3–44. [Google Scholar] [CrossRef]
- Knipling, E.F. The Basic Principles of Insect Population Suppression and Management; US Department of Agriculture: Washington, DC, USA, 1979; p. 512. [Google Scholar]
- Liebhold, A.M.; Berec, L.; Brockerhoff, E.G.; Epanchin-Niell, R.S.; Hastings, A.; Herms, D.A.; Kean, J.M.; McCullough, D.G.; Suckling, D.M.; Tobin, P.C.; et al. Eradication of invading insect populations: From concepts to applications. Annu. Rev. Entomol. 2016, 61, 335–352. [Google Scholar] [CrossRef] [Green Version]
- Suckling, D.M.; Cristofaro, M.; Roselli, G.; Levy, M.C.; Cemmi, A.; Mazzoni, V.; Stringer, L.D.; Zeni, V.; Ioriatti, C.; Anfora, G. The competitive mating of irradiated brown marmorated stink bugs, Halyomorpha halys, for the sterile insect technique. Insects 2019, 10, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendrichs, J.; Bloem, K.; Hoch, G.; Carpenter, J.E.; Greany, P.; Robinson, A.S. Improving the cost-effectiveness, trade and safety of biological control for agricultural insect pests using nuclear techniques. Biocontrol Sci. Technol. 2009, 19, 3–22. [Google Scholar] [CrossRef]
- Gurr, G.; Kvedaras, O. Synergizing biological control: Scope for sterile insect technique, induced plant defences and cultural techniques to enhance natural enemy impact. Biol. Control. 2010, 52, 198–207. [Google Scholar] [CrossRef]
- Konopka, J.K.; Gariepy, T.D.; Haye, T.; Zhang, J.; Rubin, B.D.; McNeil, J.N. Exploitation of pentatomids by native egg parasitoids in the native and introduced ranges of Halyomorpha halys: A molecular approach using sentinel egg masses. J. Pest Sci. 2019, 92, 609–619. [Google Scholar] [CrossRef]
- Abram, P.K.; Mills, N.J.; Beers, E.H. Review: Classical biological control of invasive stink bugs with egg parasitoids—what does success look like? Pest Manag. Sci. 2020, 76, 1980–1992. [Google Scholar] [CrossRef] [PubMed]
- Tillman, P.G.; Toews, M.; Blaauw, B.; Sial, A.; Cottrell, T.E.; Talamas, E.; Buntin, D.; Joseph, S.; Balusu, R.; Fadamiro, H.; et al. Parasitism and predation of sentinel eggs of the invasive brown marmorated stink bug, Halyomorpha halys (Stål)(Hemiptera: Pentatomidae), in the southeastern US. Biol. Control 2020, 145, 104247. [Google Scholar] [CrossRef]
- Cornelius, M.L.; Herlihy, M.V.; Vinyard, B.T.; Weber, D.C.; Greenstone, M.H. Parasitism and Predation on Sentinel Egg Masses of Three Stink Bug Species (Hemiptera: Pentatomidae) in Native and Exotic Ornamental Landscapes. J. Econ. Entomol. 2021, 114, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Roversi, P.F.; Binazzi, F.; Marianelli, L.; Costi, E.; Maistrello, L.; Sabbatini Peverieri, G. Searching for native egg-parasitoids of the invasive alien species Halyomorpha halys Stål (Heteroptera, Pentatomidae) in Southern Europe. Redia 2016, 99, 63–70. [Google Scholar]
- McIntosh, H.; Lowenstein, D.M.; Wiman, N.G.; Wong, J.S.; Lee, J.C. Parasitism of frozen Halyomorpha halys eggs by Trissolcus japonicus: Applications for rearing and experimentation. Biocontrol Sci. Technol. 2019, 29, 478–493. [Google Scholar] [CrossRef]
- Yang, S.-Y.; Zhan, H.-X.; Zhang, F.; Babendreier, D.; Zhong, Y.-Z.; Lou, Q.-Z.; Zhong, Y.; Zhang, J.-P. Development and fecundity of Trissolcus japonicus on fertilized and unfertilized eggs of the brown marmorated stink bug, Halyomorpha halys. J. Pest Sci. 2018, 91, 1335–1343. [Google Scholar] [CrossRef]
- Wong, W.H.L.; A Walz, M.; Oscienny, A.B.; Sherwood, J.L.; Abram, P.K. An effective cold storage method for stockpiling Halyomorpha halys (Hemiptera: Pentatomidae) eggs for field surveys and laboratory rearing of Trissolcus japonicus (Hymenoptera: Scelionidae). J. Econ. Entomol. 2021, 114, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Cristofaro, M.; Sforza, R.F.H.; Roselli, G.; Paolini, A.; Cemmi, A.; Musmeci, S.; Anfora, G.; Mazzoni, V.; Grodowitz, M. Effects of gamma irradiation on the fecundity, fertility, and longevity of the invasive stink bug pest Bagrada hilaris (Burmeister) (Hemiptera: Pentatomidae). Insects 2022, 13, 787. [Google Scholar] [CrossRef]
- Suckling, D.M.; Levy, M.C.; Roselli, G.; Mazzoni, V.; Ioriatti, C.; Deromedi, M.; Cristofaro, M.; Anfora, G. Live Traps for Adult Brown Marmorated Stink Bugs. Insects 2019, 10, 376. [Google Scholar] [CrossRef] [Green Version]
- CREA. Proposta di immissione del microimenottero Trissolcus japonicus (Ashmead), agente di controllo biologico della cimice asiatica Halyomorpha halys (Stål)—Studio del rischio. 2020; 1––32. [Google Scholar]
- Baccaro, S.; Cemmi, A.; Di Sarcina, I.; Ferrara, G. Gamma Irradiation Calliope Facility at ENEA–Casaccia Research Centre (Rome, Italy). 2019. Available online: https://www.enea.it/it/seguici/pubblicazioni/pdf-opuscoli/calliope.pdf (accessed on 3 June 2023).
- Nielsen, A.; Hamilton, G.; Matadha, D. Development rate estimation and life table analysis for Halyomorpha halys (Stål) (Hemiptera: Pentatomidae). Environ. Entomol. Environ. Entomol. 2008, 37, 348–355. [Google Scholar] [CrossRef]
- Sabbatini-Peverieri, G.; Dieckhoff, C.; Giovannini, L.; Marianelli, L.; Roversi, P.F.; Hoelmer, K. Rearing Trissolcus japonicus and Trissolcus mitsukurii for biological control of Halyomorpha halys. Insects 2020, 11, 787. [Google Scholar] [CrossRef]
- Kinnear, P.R.; Gray, C.D.; Kinnear, P.R. PASW Statistics 17 Made Simple; Psychology Press: New York, NY, USA, 2010; p. 255. [Google Scholar]
- Nelder, J.A.; Wedderburn, R.W.M. Generalized Linear Models. J. R. Stat. Soc. Ser. A 1972, 135, 370–384. [Google Scholar] [CrossRef]
- Madsen, H.; Thyregod, P. Introduction to General and Generalized Linear Models; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.r-project.org/index.html (accessed on 3 June 2023).
- Wickham, H.; Wickham, H. Programming with ggplot2. In Ggplot2: Elegant Graphics for Data Analysis; Springer: Cham, Switzerland, 2016; pp. 241–253. [Google Scholar]
- Wood, S.N. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. J. R. Stat. Soc. Ser. B Statistical Methodol. 2010, 73, 3–36. [Google Scholar] [CrossRef] [Green Version]
- Van Driesche, R.; Bellows, T.S., Jr. Biological Control; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; pp. 3–20. [Google Scholar]
- Malek, R.; Kaser, J.M.; Anfora, G.; Ciolli, M.; Khrimian, A.; Weber, D.C.; Hoelmer, K.A. Trissolcus japonicus foraging behavior: Implications for host preference and classical biological control. Biol. Control. 2021, 161, 104700. [Google Scholar] [CrossRef]
- Scala, M.; Fouani, J.M.; Zapponi, L.; Mazzoni, V.; Wells, K.E.; Biondi, A.; Baser, N.; Verrastro, V.; Anfora, G. Attraction of egg parasitoids Trissolcus mitsukurii and Trissolcus japonicus to the chemical cues of Halyomorpha halys and Nezara viridula. Insects 2022, 13, 439. [Google Scholar] [CrossRef]
- Qiu, L.F.; Yang, Z.; Tao, W. Biology and population dynamics of Trissolcus halyomorphae. Sci. Silvae Sin. 2007, 43, 62–65. [Google Scholar]
- Roselli, G.; Anfora, G.; Suckling, D.M.; Mazzoni, V.; Vanoni, V.; Menegotti, L.; Fellin, L.; Stacconi, M.V.R.; Ioriatti, C.; Cristofaro, M. Effects of irradiation on biology and mating behaviour of wild males of brown marmorated stink bug using a 6 MV medical linear accelerator. Insects 2023, 14, 460. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.N.; Stringer, L.D.; Hong, K.-J. Influence of irradiation on the biology of the brown marmorated stink bug (Hemiptera: Pentatomidae). J. Econ. Entomol. 2021, 114, 1211–1218. [Google Scholar] [CrossRef]
- Welsh, T.J.; Stringer, L.D.; Caldwell, R.; Carpenter, J.E.; Suckling, D.M. Irradiation biology of male brown marmorated stink bugs: Is there scope for the sterile insect technique? Int. J. Radiat. Biol. 2017, 93, 1357–1363. [Google Scholar] [CrossRef]
- Klassen, W. Suppression of Pest Population with Sterile Male Insects (No. 1182); Agricultural Research Service, US Department of Agriculture: Washington, DC, USA, 1971. [Google Scholar]
- Skillman, V.P.; Lee, J.C. Nutrient content of brown marmorated stink bug eggs and comparisons between experimental uses. J. Insect Sci. 2017, 17, 120. [Google Scholar] [CrossRef] [Green Version]
- Suckling, D.M.; Tobin, P.C.; McCullough, D.G.; Herms, D.A. Combining Tactics to Exploit Allee Effects for Eradication of Alien Insect Populations. J. Econ. Entomol. 2012, 105, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Horrocks, K.J.; Avila, G.A.; Holwell, G.I.; Suckling, D.M. Integrating sterile insect technique with the release of sterile classical biocontrol agents for eradication: Is the Kamikaze Wasp Technique feasible? Biocontrol 2020, 65, 257–271. [Google Scholar] [CrossRef]
- Horrocks, K.J.; Avila, G.A.; Holwell, G.I.; Suckling, D.M. Irradiation-induced sterility in an egg parasitoid and possible implications for the use of biological control in insect eradication. Sci. Rep. 2021, 11, 12326. [Google Scholar] [CrossRef] [PubMed]
- Pezzini, D.T.; Santacruz, E.C.N.; Koch, R.L. Predation and parasitism of Halyomorpha halys (Hemiptera: Pentatomidae) ggs in Minnesota. Environ. Entomol. 2018, 47, 812–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| n. of Visible Embryos | n. of Hatched Eggs | |||
|---|---|---|---|---|
| χ2 | p-Value | χ2 | p-Value | |
| Intercept | 46.186 | 1.1 × 10−11 | 67.274 | 2.4 × 10−16 |
| Gy | 28.514 | 9.3 × 10−8 | 44.762 | 2.2 × 10−11 |
| hours | 0.641 | 0.423 | 0.091 | 0.763 |
| Gy × ours | 0.480 | 0.488 | 7.662 | 5.6 × 10−3 |
| Dose (Gy) | Egg Age (hours) | n | Immature Embryos | Eggs Hatched | ||
|---|---|---|---|---|---|---|
| (n.) 1 | (%) 1 | (n.) 1 | (%) 1 | |||
| 0 | 24 | 10 | 25.67 ± 1.48 a | 91.67 ± 5.29 a | 21.80 ± 2.74 a | 77.86 ± 9.80 a |
| 0 | 48 | 6 | 26.00 ± 1.53 a | 92.86 ± 5.46 a | 24.67 ± 1.86 a | 88.10 ± 6.63 a |
| 16 | 24 | 12 | 9.77 ± 2.58 b | 30.95 ± 5.83 b | 0.00 b | 0.00 b |
| 16 | 48 | 10 | 7.00 ± 3.69 bc | 25.00 ± 13.17 bc | 0.60 ± 0.43 b | 2.14 ± 1.52 b |
| 24 | 24 | 14 | 8.50 ± 3.11 b | 30.36 ± 11.10 b | 0.00 b | 0.00 b |
| 24 | 48 | 10 | 0.00 c | 0.00 c | 1.50 ± 1.50 b | 5.36 ± 5.36 b |
| 32 | 24 | 12 | 0.00 c | 0.00 c | 0.00 b | 0.00 b |
| 32 | 48 | 24 | 2.79 ± 1.56 c | 9.97 ± 5.56 c | 2.04 ± 1.39 b | 9.07 ± 5.20 b |
| 40 | 24 | 10 | 0.00 c | 0.00 c | 0.00 b | 0.00 b |
| 40 | 48 | 10 | 0.00 c | 0.00 c | 0.00 b | 0.00 b |
| Dose (Gy) | n | Immature Embryos | Eggs Hatched | ||
|---|---|---|---|---|---|
| n. 1 | % 1 | n. 1 | % 1 | ||
| 0 | 10 | 26.50 ± 0.582 a | 95.96 ± 1.63 a | 25.50 ± 1.327 a | 92.14 ± 4.39 a |
| 24 | 10 | 8.83 ± 1.662 b | 31.55 ± 5.93 b | 6.57 ± 1.702 b | 23.47 ± 6.08 b |
| 50 | 100 | 1.08 ± 0.176 c | 3.86 ± 0.63 c | 0.23 ± 0.060 c | 0.81 ± 0.21 c |
| Parameter | Age (days) | H | df | p-Value | Irradiated | Refrigerated | SIT |
|---|---|---|---|---|---|---|---|
| Mean ± sd 1 | Mean ± sd 1 | Mean ± sd 1 | |||||
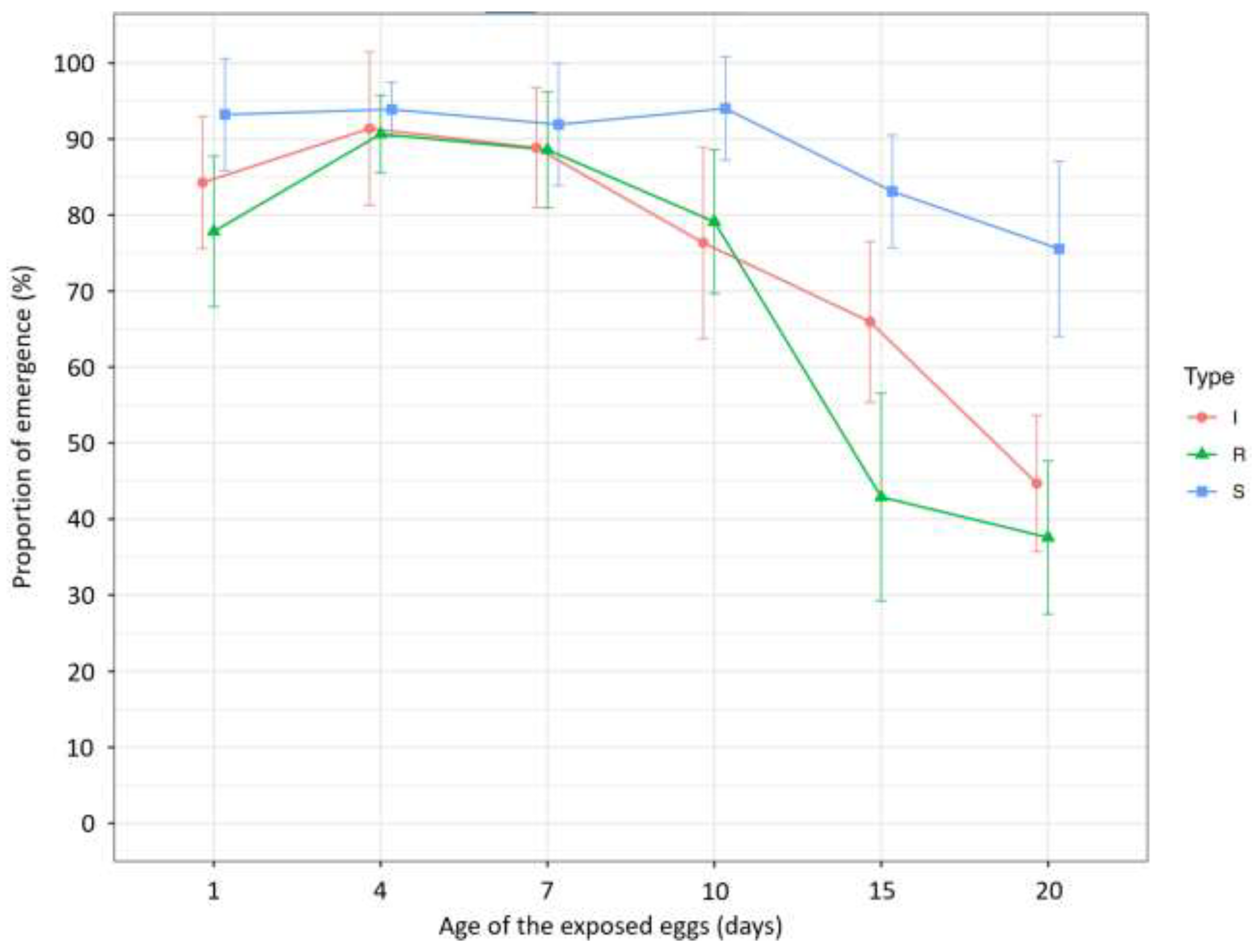
| Proportion of emergence | 1 | 17.591 | 2 | <0.001 | 84.28 ± 27.9 b | 77.85 ± 27 c | 93.23 ± 20.12 a |
| 4 | 4.464 | 2 | 0.107 | 91.39 ± 25.49 | 90.66 ± 14.58 | 93.90 ± 9.45 | |
| 7 | 5.777 | 2 | 0.056 | 88.86 ± 21.23 | 88.58 ± 20.84 | 91.93 ± 23.4 | |
| 10 | 15.244 | 2 | <0.001 | 76.36 ± 34.38 b | 79.13 ± 27.51 b | 94.03 ± 18.23 a | |
| 15 | 19.226 | 2 | <0.001 | 65.96 ± 32.1 b | 42.9 ± 35.9 c | 83.12 ± 19.82 a | |
| 20 | 25.284 | 2 | <0.001 | 44.68 ± 24.9 b | 37.57 ± 27.49 b | 75.56 ± 30.25 a | |
| Sex ratio | 1 | 13.791 | 2 | 0.001 | 81.66 ± 24.06 b | 78.48 ± 18.15 c | 88.34 ± 19.95 a |
| 4 | 2.460 | 2 | 0.292 | 84.20 ± 25.95 | 81.45 ± 25.15 | 89.25 ± 10.45 | |
| 7 | 7.664 | 2 | 0.022 | 84.11 ± 20.44 | 84.15 ± 17.19 | 87.61 ± 22.79 | |
| 10 | 19.095 | 2 | <0.001 | 61.79 ± 38.59 c | 73.82 ± 26.47 b | 91.95 ± 5.28 a | |
| 15 | 4.210 | 2 | 0.122 | 60.37 ± 31.42 b | 53.85 ± 28.8 b | 68.56 ± 32.45 a | |
| 20 | 24.839 | 2 | <0.001 | 40.55 ± 21.95 b | 39.67 ± 23.26 b | 73.45 ± 26.61 a | |
| Fecundity—0 days | 1 | 3.748 | 2 | 0.154 | 28.82 ± 4.19 | 26.14 ± 1.66 | 26.58 ± 3.36 |
| 4 | 1.851 | 2 | 0.396 | 27.7 ± 2.99 | 26.31 ± 2.92 | 27.12 ± 3.60 | |
| 7 | 7.726 | 2 | 0.021 | 23.5 ± 5.43 b | 28.95 ± 3.33 a | 28.15 ± 3.52 a | |
| 10 | 3.520 | 2 | 0.172 | 28.5 ± 3.03 | 26.48 ± 3.65 | 28.78 ± 2.97 | |
| 15 | 12.207 | 2 | 0.002 | 23.59 ± 4.58 b | 26.36 ± 1.96 ab | 28.35 ± 2.31 a | |
| 20 | 24.363 | 2 | <0.001 | 22.43 ± 1.79 b | 27.47 ± 2.07 a | 26.75 ± 2.63 a | |
| Fecundity—12 days | 1 | 2.283 | 2 | 0.319 | 53.78 ± 3.38 | 55.78 ± 3.28 | 55.21 ± 4.24 |
| 4 | 2.556 | 2 | 0.279 | 53.89 ± 4.19 b | 56.33 ± 3.2 a | 55.96 ± 4.28 ab | |
| 7 | 16.595 | 2 | <0.001 | 49.5 ± 3.67 b | 53.63 ± 3.52 b | 56.29 ± 3.11 a | |
| 10 | 2.492 | 2 | 0.288 | 53.06 ± 3.25 | 52.78 ± 3.08 | 55 ± 4.22 | |
| 15 | 17.751 | 2 | <0.001 | 49 ± 3.14 b | 53.75 ± 3.72 a | 56.63 ± 3.89 a | |
| 20 | 5.646 | 2 | 0.059 | 50.83 ± 1.8 ab | 46.53 ± 8.64 b | 53.25 ± 3.05 a | |
| Dry weight | 1 | 7.499 | 2 | 0.024 | 116.57 ± 11.98 a | 108 ± 5.87 b | 115.98 ± 8.04 a |
| 4 | 10.717 | 2 | 0.005 | 107.59 ± 5.65 b | 113.41 ± 6.22 a | 112.65 ± 8.96 a | |
| 7 | 14.239 | 2 | 0.001 | 107.87 ± 7.57 b | 111.65 ± 4.51 ab | 115.68 ± 5.89 a | |
| 10 | 7.715 | 2 | 0.021 | 108.54 ± 7.56 ab | 107.26 ± 9.6 b | 113.6 ± 8.12 a | |
| 15 | 21.866 | 2 | <0.001 | 101.57 ± 7.77 b | 115.25 ± 13.13 a | 112.12 ± 7.85 a | |
| 20 | 14.879 | 2 | 0.001 | 104.57 ± 8.65 b | 106.51 ± 4.83 b | 112.64 ± 5.23 a | |
| Longevity | 1 | 2.898 | 2 | 0.235 | 115.75 ± 44.54 | 93.41 ± 55.16 | 106.2 ± 44.26 |
| 4 | 1.504 | 2 | 0.471 | 109 ± 48.67 | 97.84 ± 39.43 | 96.21 ± 43.19 | |
| 7 | 6.399 | 2 | 0.041 | 133.64 ± 36.1 a | 121.67 ± 39.51 ab | 105.5 ± 49.97 b | |
| 10 | 1.539 | 2 | 0.463 | 107.67 ± 53.8 | 107.35 ± 34.85 | 116.24 ± 40.19 | |
| 15 | 4.877 | 2 | 0.087 | 104.9 ± 39.58 | 74.58 ± 50.52 | 101.65 ± 39.3 | |
| 20 | 11.127 | 2 | 0.004 | 120.46 ± 41.53 a | 78.9 ± 45.54 b | 103.22 ± 43.8 ab |
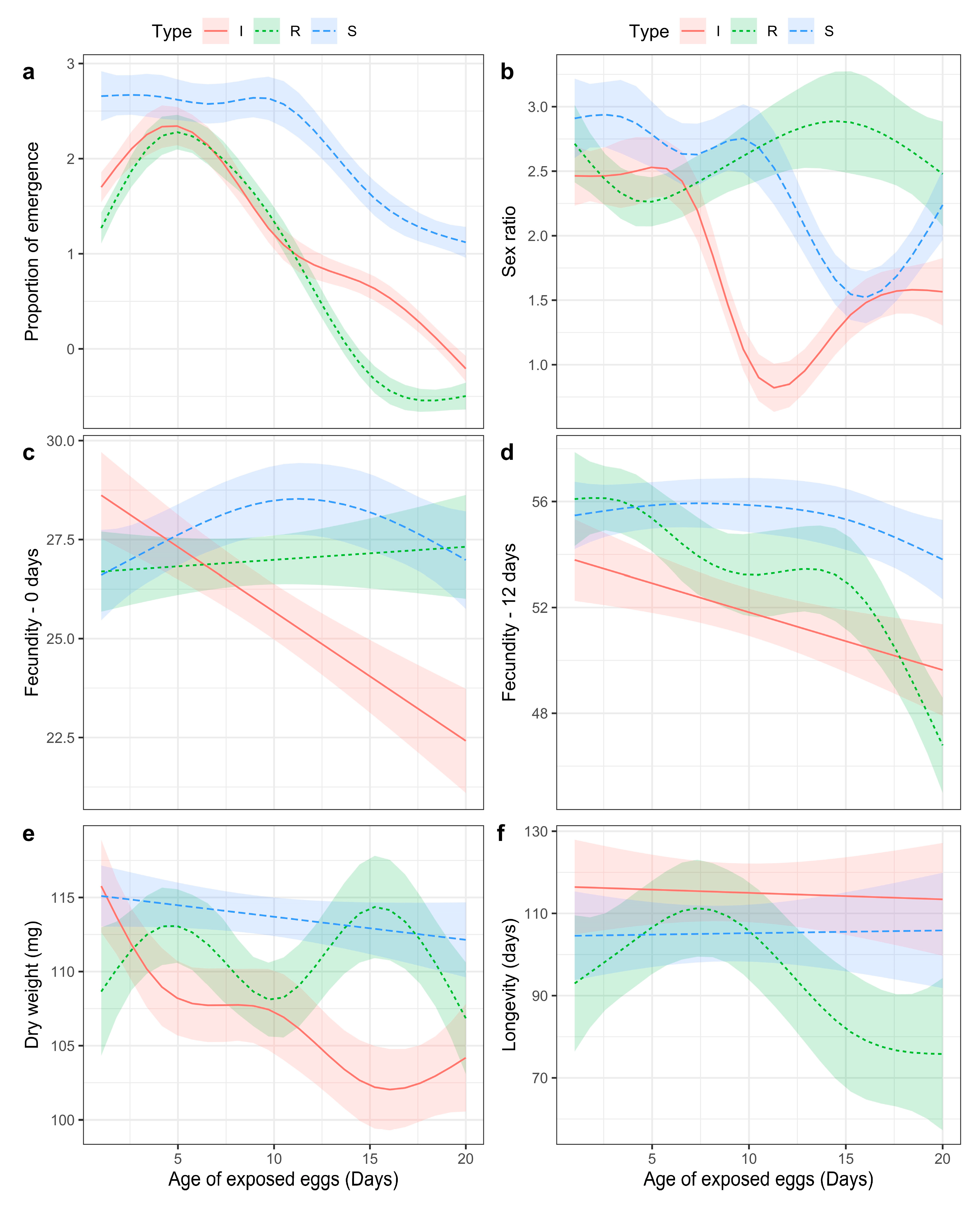
| Model | Adjusted R2 | Deviance Explained | Component | Term | Estimate | Std Error | t-Value | p-Value |
|---|---|---|---|---|---|---|---|---|
| Proportion of emergence | 30.70% | 29% | A. Parametric coefficients | (Intercept) | 1.290 | 0.037 | 34.993 | <0.001 |
| Type R | −0.272 | 0.052 | −5.265 | <0.001 | ||||
| Type S | 0.927 | 0.062 | 14.863 | <0.001 | ||||
| B. Smooth terms | s(Day):Type I | 4.536 | 4.867 | 625.785 | <0.001 | |||
| s(Day):Type R | 4.655 | 4.926 | 926.123 | <0.001 | ||||
| s(Day):Type S | 4.005 | 4.521 | 212.335 | <0.001 | ||||
| Sex ratio | 6.93% | 14% | A. Parametric coefficients | (Intercept) | 1.886 | 0.049 | 38.636 | <0.001 |
| Type R | 0.678 | 0.085 | 8.010 | <0.001 | ||||
| Type S | 0.630 | 0.077 | 8.224 | <0.001 | ||||
| B. Smooth terms | s(Day):Type I | 4.731 | 4.955 | 169.552 | <0.001 | |||
| s(Day):Type R | 3.644 | 4.293 | 11.964 | 0.025 | ||||
| s(Day):Type S | 4.479 | 4.852 | 96.429 | <0.001 | ||||
| Fecundity—0 days | 13.70% | 15% | A. Parametric coefficients | (Intercept) | 25.966 | 0.355 | 73.043 | <0.001 |
| Type R | 0.992 | 0.470 | 2.109 | 0.036 | ||||
| Type S | 1.652 | 0.448 | 3.684 | <0.001 | ||||
| B. Smooth terms | s(Day):Type I | 1.001 | 1.001 | 38.540 | <0.001 | |||
| s(Day):Type R | 1.000 | 1.000 | 0.383 | 0.536 | ||||
| s(Day):Type S | 2.347 | 2.872 | 2.438 | 0.057 | ||||
| Fecundity—12 days | 26.40% | 28% | A. Parametric coefficients | (Intercept) | 52.019 | 0.457 | 113.917 | <0.001 |
| Type R | 1.317 | 0.592 | 2.224 | 0.027 | ||||
| Type S | 3.373 | 0.565 | 5.972 | <0.001 | ||||
| B. Smooth terms | s(Day):Type I | 1.003 | 1.006 | 8.767 | 0.003 | |||
| s(Day):Type R | 3.624 | 4.224 | 17.204 | <0.001 | ||||
| s(Day):Type S | 2.073 | 2.540 | 1.984 | 0.102 | ||||
| Dry weight | 19.50% | 22% | A. Parametric coefficients | (Intercept) | 107.723 | 0.687 | 156.807 | <0.001 |
| Type R | 2.764 | 0.993 | 2.783 | 0.006 | ||||
| Type S | 6.092 | 0.951 | 6.405 | <0.001 | ||||
| B. Smooth terms | s(Day):Type I | 3.689 | 4.275 | 10.147 | <0.001 | |||
| s(Day):Type R | 4.209 | 4.705 | 3.543 | 0.004 | ||||
| s(Day):Type S | 1.000 | 1.000 | 2.335 | 0.127 | ||||
| Longevity | 4.59% | 6% | A. Parametric coefficients | (Intercept) | 115.147 | 3.612 | 31.875 | <0.001 |
| Type R | −19.087 | 5.082 | −3.755 | <0.001 | ||||
| Type S | −10.033 | 4.999 | −2.007 | 0.045 | ||||
| B. Smooth terms | s(Day):Type I | 1.000 | 1.001 | 0.079 | 0.779 | |||
| s(Day):Type R | 2.995 | 3.613 | 3.946 | 0.009 | ||||
| s(Day):Type S | 1.003 | 1.006 | 0.015 | 0.913 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roselli, G.; Anfora, G.; Sasso, R.; Zapponi, L.; Musmeci, S.; Cemmi, A.; Suckling, D.M.; Hoelmer, K.A.; Ioriatti, C.; Cristofaro, M. Combining Irradiation and Biological Control against Brown Marmorated Stink Bug: Are Sterile Eggs a Suitable Substrate for the Egg Parasitoid Trissolcus japonicus? Insects 2023, 14, 654. https://doi.org/10.3390/insects14070654
Roselli G, Anfora G, Sasso R, Zapponi L, Musmeci S, Cemmi A, Suckling DM, Hoelmer KA, Ioriatti C, Cristofaro M. Combining Irradiation and Biological Control against Brown Marmorated Stink Bug: Are Sterile Eggs a Suitable Substrate for the Egg Parasitoid Trissolcus japonicus? Insects. 2023; 14(7):654. https://doi.org/10.3390/insects14070654
Chicago/Turabian StyleRoselli, Gerardo, Gianfranco Anfora, Raffaele Sasso, Livia Zapponi, Sergio Musmeci, Alessia Cemmi, David Maxwell Suckling, Kim Alan Hoelmer, Claudio Ioriatti, and Massimo Cristofaro. 2023. "Combining Irradiation and Biological Control against Brown Marmorated Stink Bug: Are Sterile Eggs a Suitable Substrate for the Egg Parasitoid Trissolcus japonicus?" Insects 14, no. 7: 654. https://doi.org/10.3390/insects14070654