
Planting Date of Cotton in the Brazilian Cerrado Drives Boll Weevil (Coleoptera: Curculionidae) Infestation

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
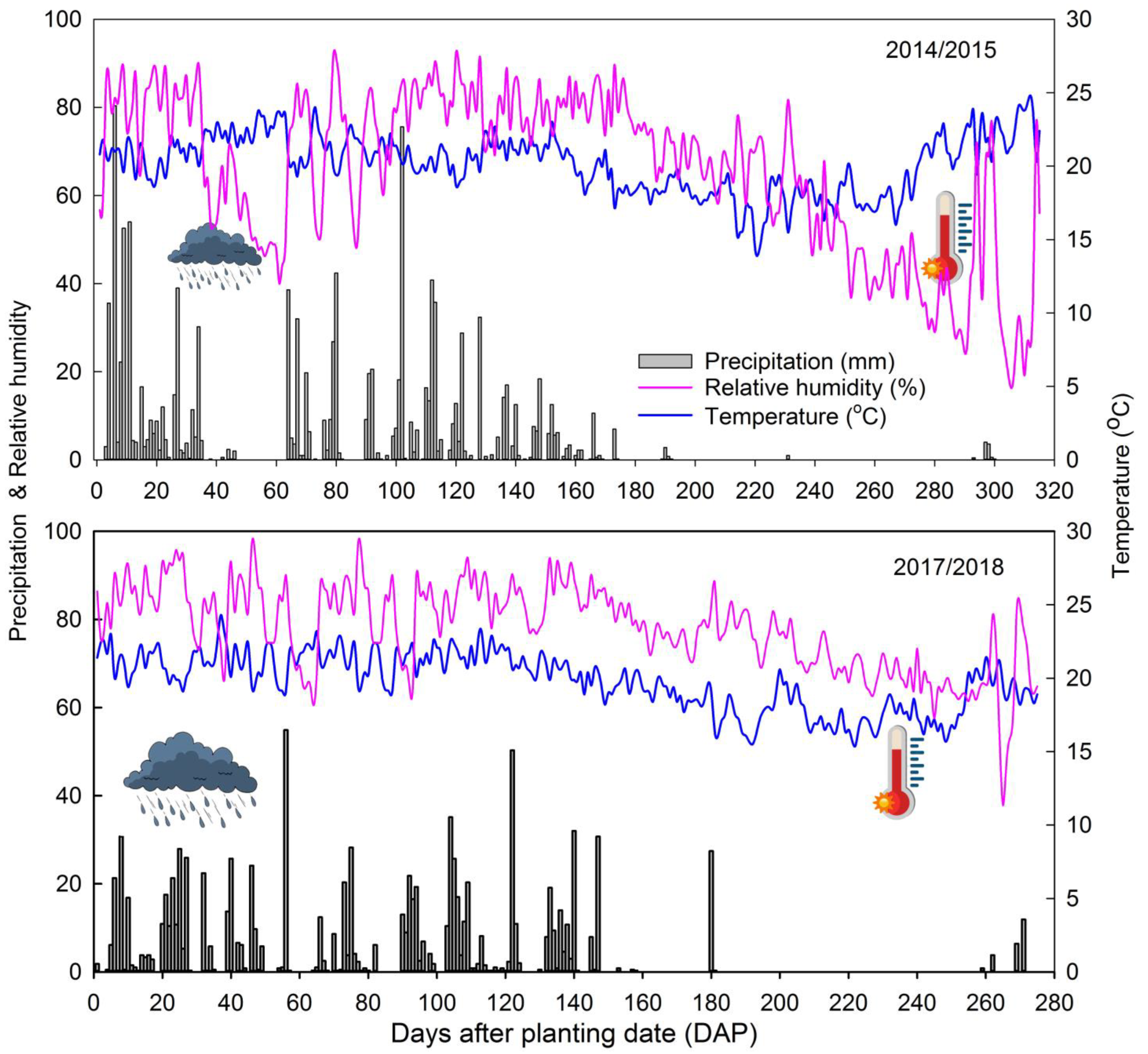
2. Materials and Methods
2.1. General Conditions
2.2. Variables
2.3. Statistical Analysis
3. Results
3.1. Time for Producing the First Reproductive Structures
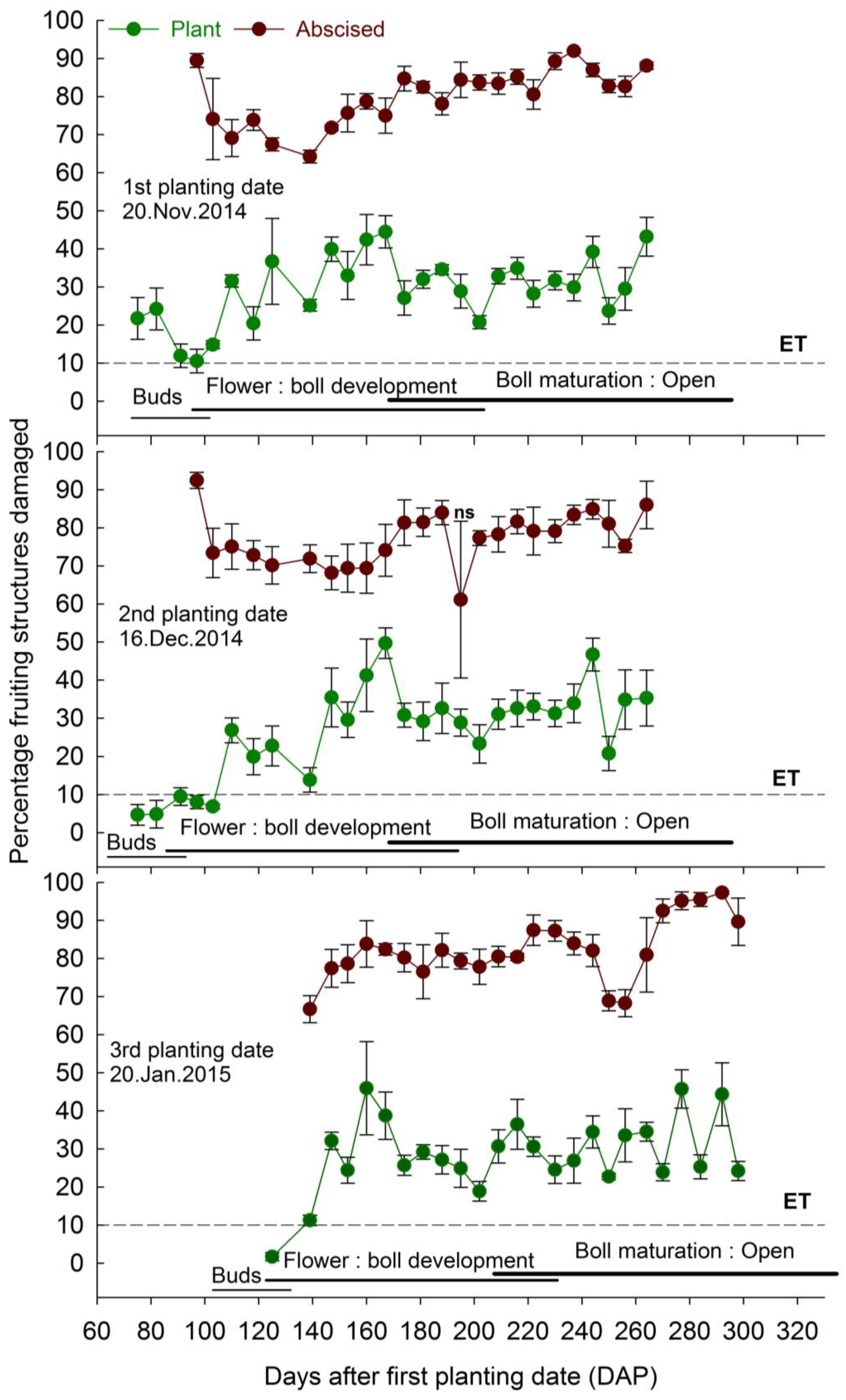
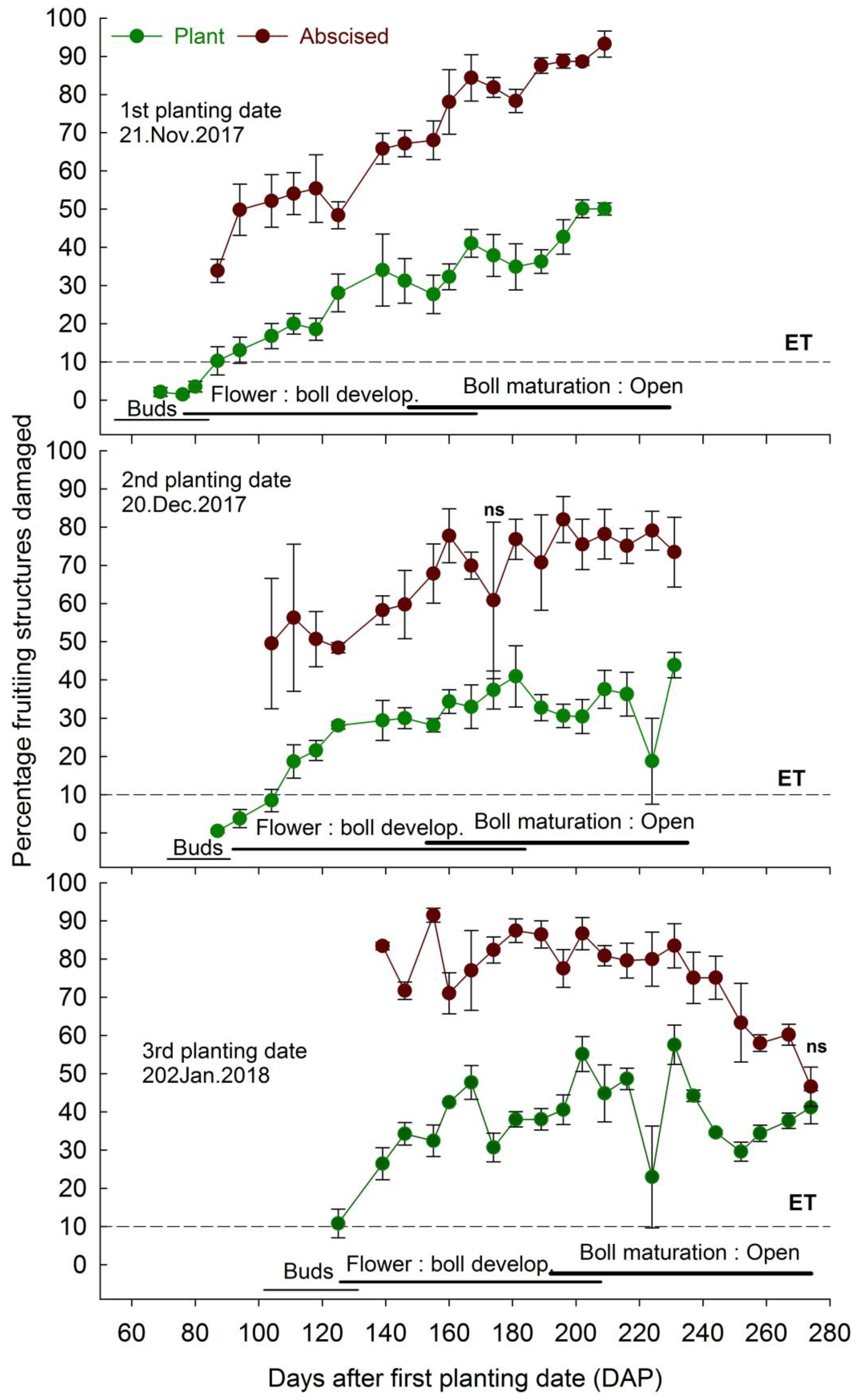
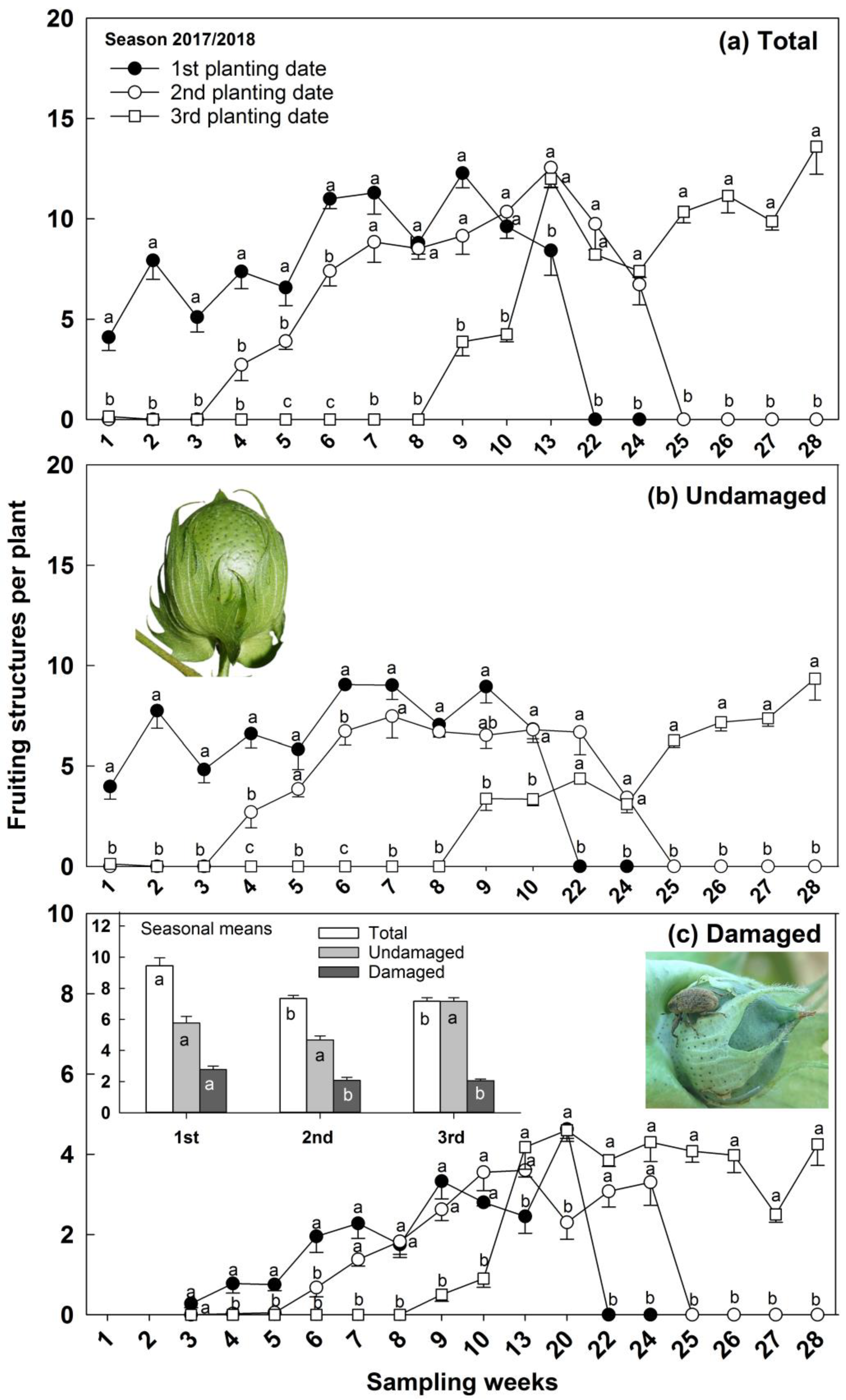
3.2. Infestation on Plant and Abscised Fruiting Structures
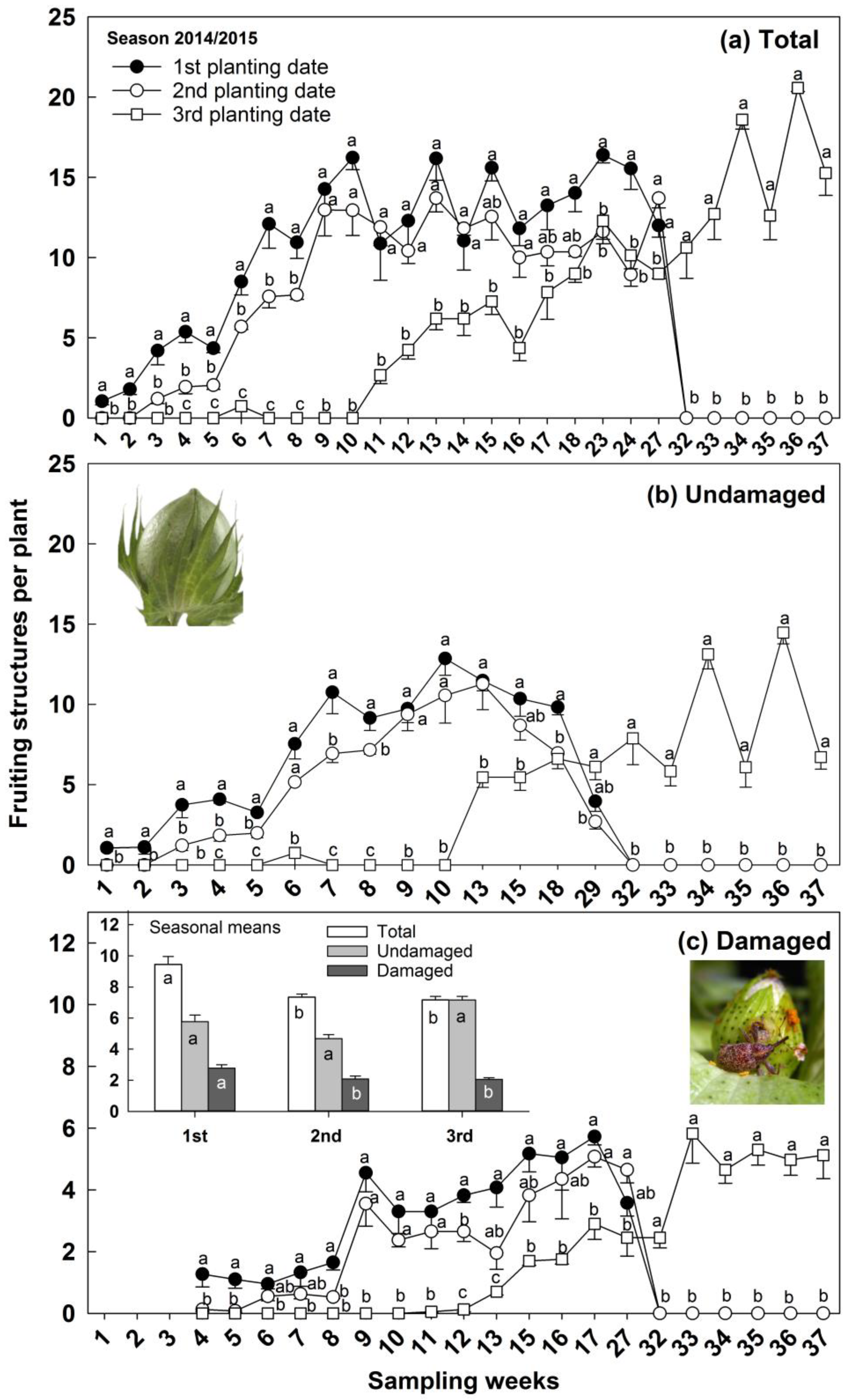
3.3. Reproductive Structures Total, Undamaged, and Damaged, and Boll Weevil Immature and Adult Numbers
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cividanes, F.J.; Souza, V.P. Exigências térmicas e tabelas de vida de fertilidade de Myzus persicae (Sulzer) (Hemiptera: Aphididae) em laboratório. Neotrop. Entomol. 2003, 32, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Slosser, J.E. Influence of planting date and insecticide treatment on insect pest abundance and damage in dryland cotton. J. Econ. Entomol. 1993, 86, 1213–1222. [Google Scholar] [CrossRef]
- Greenberg, S.M.; Jones, G.D.; Adamczyk, J.J., Jr.; Eischen, F.; Armstrong, J.S.; Coleman, R.J.; Sétamou, T.X.L. Reproductive potential of field-collected overwintering boll weevils (Coleoptera: Curculionidae) fed on pollen in the laboratory. Insect Sci. 2009, 16, 321–327. [Google Scholar] [CrossRef]
- Showler, A.T.; Greenberg, S.M.; Scott, A.W., Jr.; Robinson, J.R.C. Effects of planting dates on boll weevils (Coleoptera: Curculionidae) and cotton fruit in the subtropic. J. Econ. Entomol. 2005, 98, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Showler, A.T. Relationships of abscised cotton fruit to boll weevil (Coleoptera: Curculionidae) feeding, oviposition, and development. J. Econ. Entomol. 2008, 101, 68–73. [Google Scholar] [CrossRef]
- Soares, J.J.; Lara, F.M.; Carvalho, L.P. Uso de algodão resistente. Rev. Cult. Grandes Cult. 1999, 8, 16–18. [Google Scholar]
- Freire, E.C.; Medeiros, J.C.; Silva, C.A.D.; Azevedo, D.M.P.; Andrade, F.P.; Vieira, D.J. Cultura dos Algodoeiros Mocó Precoce e Algodão 7MH; Embrapa Algodão: Campina Grande, Brazil, 1999; pp. 6–10. [Google Scholar]
- White, J.H.; Rummel, D.H. Emergence profile of overwintering boll weevils and emergence to cotton. Environ. Entomol. 1978, 7, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Karavina, C.; Mamdumbu, R.; Parwada, C.; Mungunyana, T. Variety and planting date effects on the incidence of bollworms and insect sucking pests of cotton (Gossypium hirsutum L.). J. Agri. Sci. 2012, 3, 607–610. [Google Scholar]
- Pires, C.S.S.; Pimenta, M.; Mata, R.A.; Souza, L.C.; Paula, D.P.; Sujii, E.R.; Fontes, E.M.G. Survival pattern of the boll weevil during cotton fallow in Midwestern Brazil. Pesq. Agrop. Bras. 2017, 52, 149–160. [Google Scholar] [CrossRef] [Green Version]
- Nyabanga, L.; Mandumbu, R.; Rugare, J.T.; Mafuse, N.; Zivenge, E.; Tibugari, H.; Nyamadzawo, G.; Gadzirayi, C.T. Preventing fall armyworm (Spodoptera frugiperda JE Smith) damage in maize by altering planting time and using varied genotypes. SDGs 2021, 2, 47–61. [Google Scholar] [CrossRef]
- Degrande, P.E. Manejo de praga em algodão. Rev. Cult. Grandes Cult. 2002, 42, 14–16. [Google Scholar]
- Jin, Z.; Yu, W.; Zhao, H.; Xian, X.; Jing, K.; Yang, N.; Lu, X.; Liu, W. Potential global distribution of invasive alien species, Anthonomus grandis Boheman, under current and future climate using optimal maxent model. Agriculture 2022, 12, 1759. [Google Scholar] [CrossRef]
- Jeger, M.; Bragard, C.; Caffier, D.; Candresse, T.; Chatzivassiliou, E.; Dehnen-Schmutz, K.; Gilioli, G.; Gregoire, J.-C.; Miret, J.A.J.; Navarro, M.N.; et al. Pest categorisation of Anthonomus grandis. EFSA 2017, 15, 50–74. [Google Scholar] [CrossRef] [Green Version]
- Showler, A.T. Influence of cotton fruit stages as food sources on boll weevil (Coleoptera: Curculionidae) fecundity and oviposition. J. Econ. Entomol. 2004, 97, 1330–1334. [Google Scholar] [CrossRef]
- Showler, A.T. Relationships of different cotton square sizes to boll weevil (Coleoptera: Curculionidae) feeding and oviposition in field conditions. J. Econ. Entomol. 2005, 98, 572–579. [Google Scholar] [CrossRef]
- Busoli, A.C.; Pereira, F.F.; Lopéz, V.A.G.; Soares, J.J.; Melo, R.S.; Almeida, C.A. Preferência alimentar do bicudo-do-algodoeiro por frutos de diferentes cultivares e idades. Pesq. Agrop. Bras. 2004, 39, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.L.A.L.; Silva, C.A.D. Concentração eficiente e econômica de caulim para a proteção de algodoeiro contra o bicudo. Pesq. Agrop. Bras. 2015, 50, 763–768. [Google Scholar] [CrossRef] [Green Version]
- Gondim, D.M.C. Manual de Identificação das Pragas, Doenças, Deficiências Minerais e Injúrias do Algodoeiro no Brasil, 3rd ed.; Coodetec: Cascavel, Brazil, 1999; 120p. [Google Scholar]
- Bastos, C.S.; Pereira, M.J.B.; Takizawa, E.K.; Ohl, G.; Aquino, V.R. Bicudo do Algodoeiro: Identificação, Biologia, Amostragem e Táticas de Controle; Embrapa Algodão: Campina Grande, Brazil, 2005; 31p. [Google Scholar]
- Degrande, P.E. Guia Prático de Controle das Pragas do Algodoeiro; Universidade Federal do Mato Grosso de Sul: Dourados, Brazil, 1998; 60p. [Google Scholar]
- Papa, G.; Celoto, F.J. Controle químico do bicudo-do-algodoeiro Anthonomus grandis Boheman (Coleoptera: Curculionidae). In O Bicudo-do-Algodoeiro (Anthonomus Grandis Boh., 1843) Nos Cerrados Brasileiros: Biologia e Medidas de Controle; Belot, J.L., Ed.; IMAmt: Cuiabá, Brazil, 2015; pp. 140–150. [Google Scholar]
- Torres, J.B.; Rolim, G.G.; Arruda, L.S.; Santos, M.P.; Leite, S.A.; Neves, R.C.S. Insecticides in use and risk of control failure of boll weevil (Coleoptera: Curculionidae) in the brazilian cerrado. Neotrop. Entomol. 2022, 51, 613–627. [Google Scholar] [CrossRef]
- Rolim, G.G.; Coelho, R.R.; Dantonino, J.; Arruda, L.S.; Rodrigues, A.S.; Barros, E.M.; Torres, J.B. Field-evolved resistance to beta-cyfluthrin in the boll weevil: Detection and characterization. Pest Manag. Sci. 2021, 77, 4400–4410. [Google Scholar] [CrossRef]
- Leigh, T.F.; Roach, S.H.; Watson, T.F. Biology and ecology of important insect and mite pests of cotton. In Cotton Insects and Mites: Characterization and Management; King, E.G., Phillips, J.R., Coleman, R.J., Eds.; The Cotton Foundation: Memphis, TN, USA, 1996; pp. 16–86. [Google Scholar]
- Higley, L.G.; Pedigo, L.P. Economic Thresholds for Integrated Pest Management; University of Nebraska Press: Lincoln, NE, USA, 1996; 688p. [Google Scholar]
- Killi, F.; Bolek, Y. Timing of planting is crucial for cotton yield. Acta Agri. Scand. B Soil Plant. Sci. 2006, 56, 155–160. [Google Scholar] [CrossRef]
- Zoneamento Agrícola de Risco Climático (ZARC). Available online: https://indicadores.agricultura.gov.br/zarc/index.htm (accessed on 10 April 2023).
- Paula, D.P.; Claudino, D.; Timbó, R.V.; Miranda, J.E.; Bemquerer, M.P.; Ribeiro, A.C.J.; Sujii, E.R.; Fontes, E.M.G.; Pires, C.S.S. Reproductive dormancy in boll-weevil from populations of the midwest of Brazil. J. Econ. Entomol. 2013, 106, 86–96. [Google Scholar] [CrossRef]
- Pimenta, M.; Mata, R.A.; Venzon, M.; Cunha, D.N.C.; Fontes, E.M.G.; Pires, C.S.S.; Sujii, E.R. Survival and preference of cotton boll weevil adults for alternative food sources. Braz. J. Biol. 2016, 76, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, S.M.; Sparks, A.N., Jr.; Norman, J.W., Jr.; Coleman, R.; Bradford, J.M.; Yang, C.; Sappington, T.W.; Showler, A. Chemical cotton stalk destruction for maintenance of host-free periods for the control of overwintering boll weevil in tropical and subtropical climates. Pest Manag. Sci. 2007, 63, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Neves, R.C.S.; Torres, J.B.; Barros, E.M.; Vivan, L.M. Boll weevil within season and off-season activity monitored using a pheromone-and-glue reusable tube trap. Sci. Agri. 2018, 75, 313–320. [Google Scholar] [CrossRef]
- Arruda, L.S.; Torres, J.B.; Rolim, G.G.; Silva-Torres, C.S.A. Dispersal of boll weevil toward and within the cotton plant and implications for insecticide exposure. Pest Manag. Sci. 2021, 77, 1339–1347. [Google Scholar] [CrossRef]
- Miranda, J.E.; Rodrigues, S.M.M. História do bicudo no Brasil. In O Bicudo-do-Algodoeiro (Anthonomus Grandis Boh., 1843) Nos Cerrados Brasileiros: Biologia e Medidas de Controle; Belot, J.L., Ed.; IMAmt: Cuiabá, Brazil, 2015; pp. 10–44. [Google Scholar]
- Arruda, L.S.; Rolim, G.G.; Barros, E.M.; Pereira, F.F.; Torres, J.B. Colonization and spatial distribution of boll weevil in commercial cotton fields. Neotrop. Entomol. 2020, 49, 901–915. [Google Scholar] [CrossRef]
- Paim, E.A.; Dias, A.M.; Showler, A.T.; Campos, K.L.; Oliveira, A.A.S.; Grillo, P.P.C.; Bastos, C.S. Cotton row spacing for boll weevil management in low-input production systems. Crop Prot. 2021, 145, 105614. [Google Scholar] [CrossRef]
- Showler, A.T.; Cantú, R.V. Intervals between boll weevil (Coleoptera: Curculionidae) oviposition and square abscission, and development to adulthood in lower Rio Grande Valley, Texas, field conditions. Southwest. Entomol. 2005, 30, 161–164. [Google Scholar]
- SAS. The SAS System, version 9.00; SAS Institute: Cary, NC, USA, 2002. [Google Scholar]
- Ramalho, F.S.; Jesus, F.M.M. Avaliação de inseticidas para o controle do bicudo-do-algodoeiro. Pesq. Agrop. Bras. 1989, 24, 273–276. [Google Scholar]
- Walsh, P.J.; Miller, A.J. The relation between degree days and base temperature. Appl. Energy 1983, 13, 241–253. [Google Scholar] [CrossRef]
- Pacheco, R.I.L.; Álvarez, E.M.C.; Ferrer, J.L.R.; Bonilla, H.A.; Gómez-Correa, J.C.; Hernández, M.J.Y.; Artiles, L.P. Accumulation of degree days and their effect on the potential yield of 15 eggplant (Solanum melongena L.) accessions in the Colombian Caribbean. Rev. Fac. Nac. Agron. Medellín 2019, 72, 8917–8926. [Google Scholar] [CrossRef] [Green Version]
- Viator, R.P. Predicting cotton boll maturation period using degree days and other climatic factors. Agron. J. 2005, 97, 494–499. [Google Scholar] [CrossRef] [Green Version]
- Showler, A.T.; Abrigo, V. Common subtropical and tropical nonpollen food sources of the boll weevil (Coleoptera: Curculionidae). Environ. Entomol. 2007, 36, 99–104. [Google Scholar] [CrossRef]
- Greenberg, S.M.; Sappington, T.W.; Spurgeon, D.W.; Sétamou, M. Boll weevil (Coleoptera: Curculionidae) feeding and reproduction as functions of cotton square availability. Environ. Entomol. 2003, 32, 698–704. [Google Scholar] [CrossRef] [Green Version]
- Sterling, W.; Dean, A.; Hartstack, A.; Witz, J. Partitioning boll weevil (Coleoptera: Curculionidae) mortality associated with high temperature: Desiccation or thermal death? Environ. Entomol. 1990, 19, 1457–1462. [Google Scholar] [CrossRef]
- Slosser, J.E. Cultural Control of the Boll Weevil: A Four Season Approach—Texas Rolling Plains. B-1721; Texas A&M University: College Station, TX, USA, 1995; 14p. [Google Scholar]
- Bueno, A.F.; Paula-Moraes, S.V.; Gazzoni, D.L.; Pomari, A.F. Economic thresholds in soybean-integrated pest management: Old concepts, current adoption, and adequacy. Neotrop. Entomol. 2013, 42, 439–447. [Google Scholar] [CrossRef]
- Hill, M.P.; Macfadyen, S.; Nash, M.A. Broad spectrum pesticide application alters natural enemy communities and may facilitate secondary pest outbreaks. PeerJ 2017, 5, e4179. [Google Scholar] [CrossRef] [Green Version]
- Keasar, T.; Wajnberg, E.; Heimpel, G.; Hardy, I.C.W.; Harpaz, L.S.; Gottlieb, D.; van Nouhuys, S. Dynamic economic thresholds for insecticide applications against agricultural pests: Importance of pest and natural enemy migration. J. Econ. Entomol. 2023, 116, 321–330. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reproductive Structures | Planting Dates | Statistics, Fp-Value | ||
|---|---|---|---|---|
| 20 November 2014 | 16 December 2014 | 20 January 2015 | df = 2, 8 | |
| Flower bud | 51.5 ± 1.44 a | 39.5 ± 0.50 b | 54.2 ± 0.75 a | F = 61.650.0001 |
| White flower | 79.7 ± 1.25 a | 67.0 ± 1.00 b | 76.5 ± 0.29 a | F = 44.450.0001 |
| Young boll | 81.7 ± 1.25 a | 69.0 ± 1.00 b | 78.2 ± 0.25 a | F = 44.180.0001 |
| Open boll | 172.5 ± 0.29 a | 149.7 ± 1.44 c | 167.7 ± 0.25 b | F = 232.370.0001 |
| 21 November 2017 | 20 December 2017 | 22 January 2018 | df = 2, 9 | |
| Flower bud | 54.3 ± 0.98 a | 43.3 ± 0.58 b | 57.6 ± 1.02 a | F = 71.500.0001 |
| White flower | 84.4 ± 0.49 a | 79.0 ± 0.76 b | 84.2 ± 1.11 a | F =13.960.0017 |
| Young boll | 92.5 ± 0.80 ab | 89.6 ± 1.06 b | 97.2 ± 1.88 a | F = 8.440.0086 |
| Open Boll | 197.9 ± 1.80 b | 193.9 ± 0.54 b | 206.8 ± 0.65 a | F = 33.160.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, P.J.; Dias, A.M.; Campos, K.L.; Araújo, A.C.A.; Oliveira, A.A.S.; Suinaga, F.A.; Torres, J.B.; Bastos, C.S. Planting Date of Cotton in the Brazilian Cerrado Drives Boll Weevil (Coleoptera: Curculionidae) Infestation. Insects 2023, 14, 599. https://doi.org/10.3390/insects14070599
Santos PJ, Dias AM, Campos KL, Araújo ACA, Oliveira AAS, Suinaga FA, Torres JB, Bastos CS. Planting Date of Cotton in the Brazilian Cerrado Drives Boll Weevil (Coleoptera: Curculionidae) Infestation. Insects. 2023; 14(7):599. https://doi.org/10.3390/insects14070599
Chicago/Turabian StyleSantos, Patrícia J., Antônio M. Dias, Karolayne L. Campos, Ana C. A. Araújo, Andréa A. S. Oliveira, Fábio A. Suinaga, Jorge B. Torres, and Cristina S. Bastos. 2023. "Planting Date of Cotton in the Brazilian Cerrado Drives Boll Weevil (Coleoptera: Curculionidae) Infestation" Insects 14, no. 7: 599. https://doi.org/10.3390/insects14070599