
A New qPCR Assay for the Rapid Diagnosis of Anthonomus grandis Subspecies
 , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
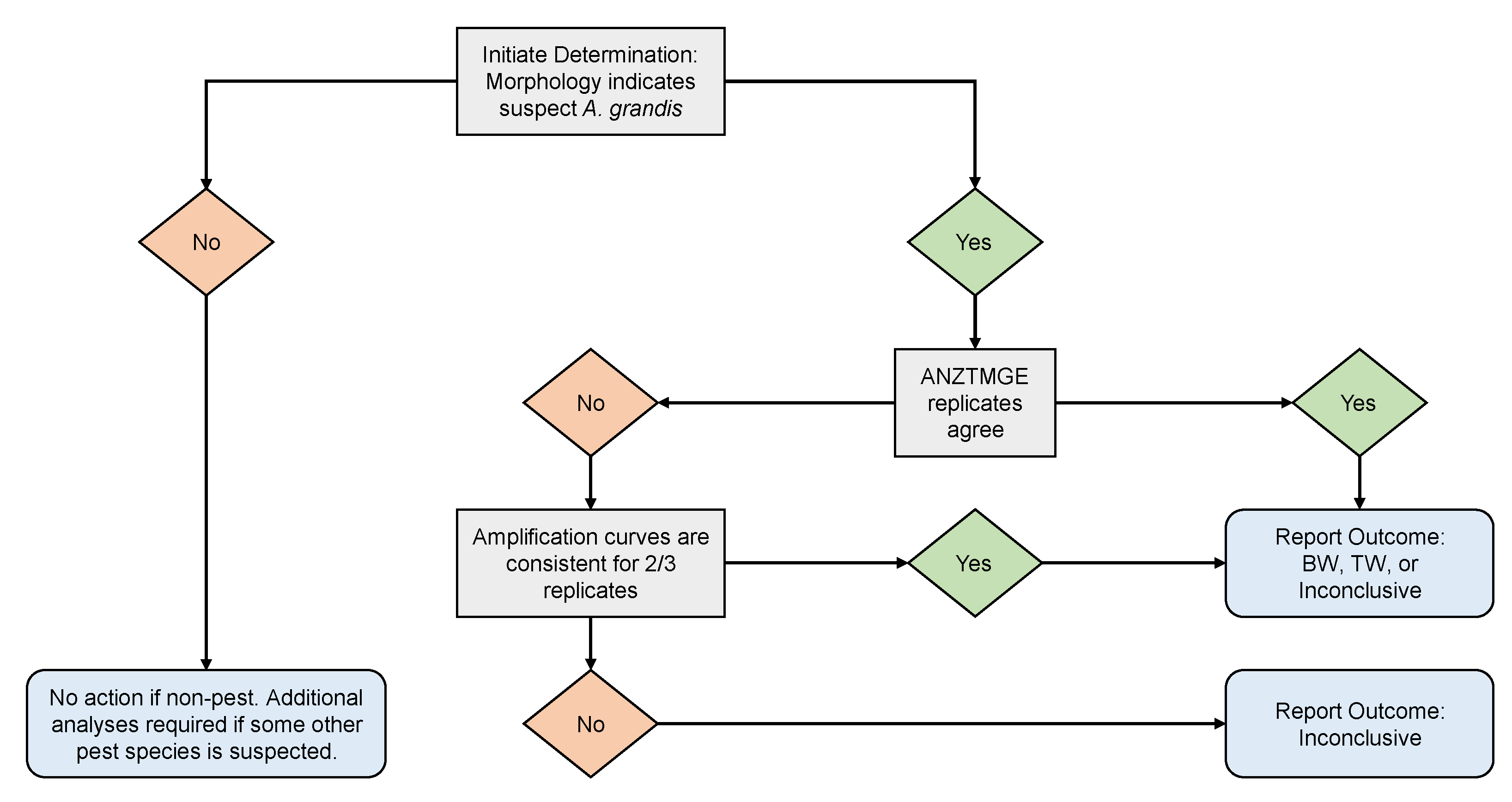
2. Materials and Methods
2.1. Initial Assay Design and Locus Selection
2.2. Validation of the ANZTMGE Assay
2.2.1. Experimental Approach to Assay Validation
2.2.2. Specimen Sampling and DNA Isolation
2.2.3. PCR Setup and Run Conditions at IMMDL and ICCDRU
2.2.4. PCR Setup and Run Conditions at TAMU
2.2.5. Quantification Cycle and Endpoint Analysis and Interpretation for Variant Diagnosis
2.2.6. Sensitivity Test
2.2.7. Non-target Specificity Test
2.2.8. Accuracy Test
3. Results
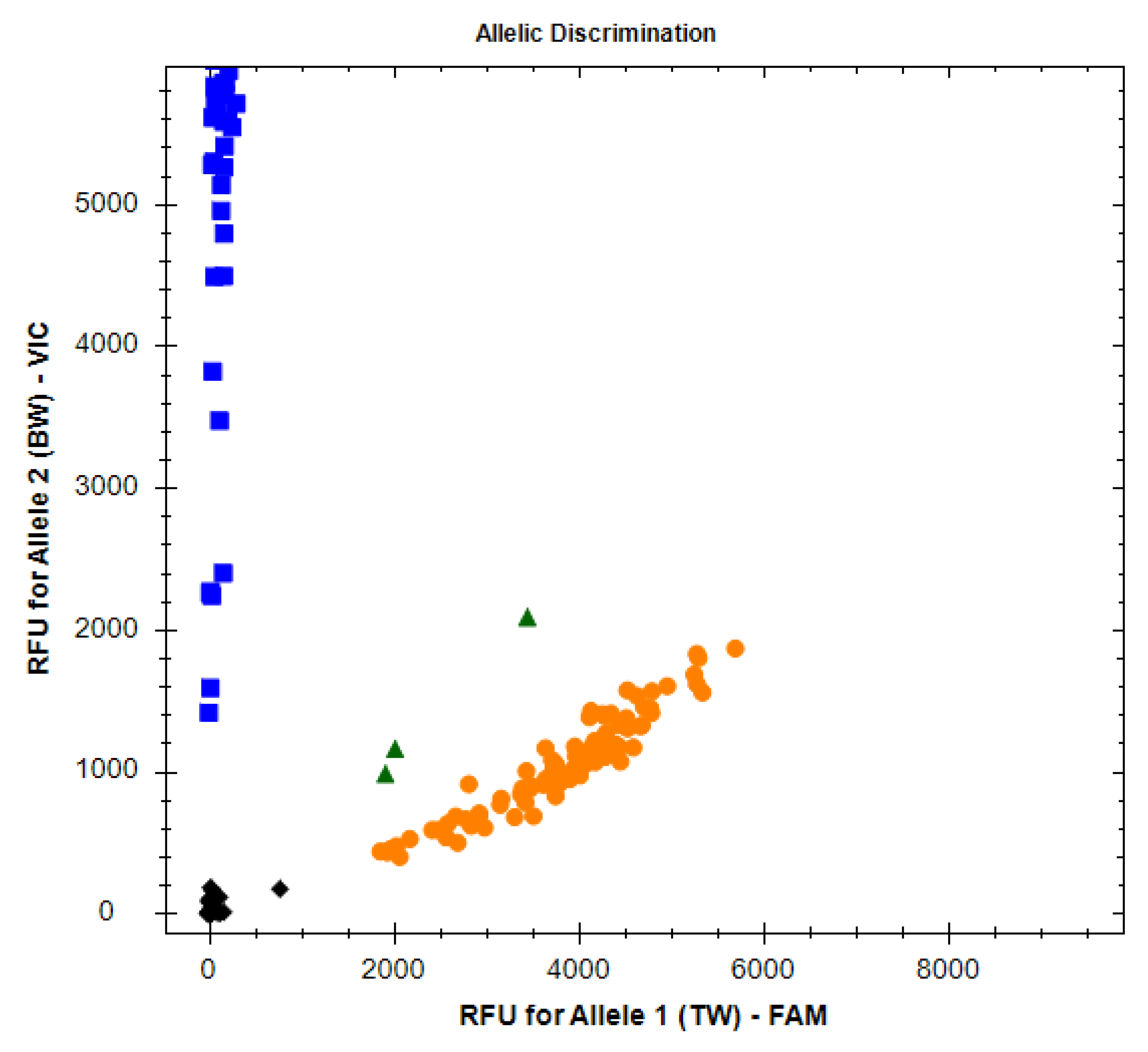
3.1. Quantification Cycle Analysis
3.2. Sensitivity Test
3.3. Non-Target Specificity Test
3.4. Accuracy Test
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burke, H.R.; Clark, W.E.; Cate, J.R.; Fryxell, P.A. Origin and Dispersal of the Boll Weevil. Bull. Entomol. Soc. Am. 1986, 32, 228–238. [Google Scholar] [CrossRef]
- Lange, F.; Olmstead, A.L.; Rhode, P.W. The Impact of the Boll Weevil, 1892–1932. J. Econ. Hist. 2009, 69, 685–718. [Google Scholar] [CrossRef]
- Raszick, T.J. Boll Weevil Eradication: A Success Story of Science in the Service of Policy and Industry. Ann. Entomol. Soc. Am. 2021, 114, 702–708. [Google Scholar] [CrossRef]
- Cross, W.H.; Lukefahr, M.J.; Fryxell, P.A.; Burke, H.R. Host Plants of the Boll Weevil 12. Environ. Entomol. 1975, 4, 19–26. [Google Scholar] [CrossRef]
- Sánchez-Reyes, U.J.; Jones, R.W.; Raszick, T.J.; Ruiz-Arce, R.; Sword, G.A. Potential Distribution of Wild Host Plants of the Boll Weevil (Anthonomus grandis) in the United States and Mexico. Insects 2022, 13, 337. [Google Scholar] [CrossRef]
- Showler, A.T. Short-Range Dispersal and Overwintering Habitats of Boll Weevils (Coleoptera: Curculionidae) during and after Harvest in the Subtropics. J. Econ. Entomol. 2006, 99, 1152–1160. [Google Scholar] [CrossRef]
- Spurgeon, D.W.; Raulston, J.R. Captures of Boll Weevils (Coleoptera: Curculionidae) in Traps Associated with Different Habitats. J. Econ. Entomol. 2006, 99, 752–756. [Google Scholar] [CrossRef]
- Raszick, T.J.; Dickens, C.M.; Perkin, L.C.; Tessnow, A.E.; Suh, C.P.-C.; Ruiz-Arce, R.; Boratynski, T.N.; Falco, M.R.; Johnston, J.S.; Sword, G.A. Population Genomics and Phylogeography of the Boll Weevil, Anthonomus Grandis Boheman (Coleoptera: Curculionidae), in the United States, Northern Mexico, and Argentina. Evol. Appl. 2021, 14, 1778–1793. [Google Scholar] [CrossRef]
- Alvarado, A.; Jones, R.W.; Pedraza-Lara, C.; Alvarado Villanueva, O.; Pfeiler, E. Reassessment of the Phylogeography and Intraspecific Relationships of Western and Eastern Populations of the Boll Weevil, Anthonomus Grandis Boheman (Coleoptera: Curculionidae), in North America. Biol. J. Linn. Soc. 2017, 122, 29–45. [Google Scholar] [CrossRef]
- Warner, R.E. Taxonomy of the Subspecies of Anthonomus Grandis (Coleoptera: Curculionidae). Ann. Entomol. Soc. Am. 1966, 59, 1073–1088. [Google Scholar] [CrossRef]
- Kuester, A.P.; Jones, R.W.; Sappington, T.W.; Kim, K.S.; Barr, N.B.; Roehrdanz, R.L.; Senechal, P.; Nason, J.D. Population Structure and Genetic Diversity of the Boll Weevil (Coleoptera: Curculionidae) on Gossypium in North America. Ann. Entomol. Soc. Am. 2012, 105, 902–916. [Google Scholar] [CrossRef]
- Allen, C.T. Boll Weevil Eradication: An Areawide Pest Management Effort. In Areawide Pest Management: Theory and Implementation; CABI: Wallingford, UK, 2008; pp. 467–559. [Google Scholar] [CrossRef]
- Cross, W.H.; Hardee, D.D.; Nichols, F.; Mitchell, H.C.; Mitchell, E.B.; Huddleston, P.M.; Tumlinson, J.H. Attraction of Female Boll Weevils to Traps Baited with Males or Extracts of Males. J. Econ. Entomol. 1969, 62, 154–161. [Google Scholar] [CrossRef]
- Hardee, D.D.; McKibben, G.H.; Gueldner, R.C.; Mitchell, E.B.; Tumlinson, J.H.; Cross, W.H. Boll Weevils in Nature Respond to Grandlure, a Synthetic Pheromone. J. Econ. Entomol. 1972, 65, 97–100. [Google Scholar] [CrossRef]
- Tumlinson, J.H.; Hardee, D.D.; Gueldner, R.C.; Thompson, A.C.; Hedin, P.A.; Minyard, J.P. Sex Pheromones Produced by Male Boll Weevil: Isolation, Identification, and Synthesis. Science 1969, 166, 1010–1012. [Google Scholar] [CrossRef]
- Jones, R.G.; Williams, M.R. A Field Guide to Boll Weevil Identification. In Technical Bulletin; Mississippi Agricultural and Forestry Experiment Station: Starkville, MS, USA, 2001; Volume 228. [Google Scholar]
- Barr, N.; Ruiz–Arce, R.; Obregón, O.; De Leon, R.; Foster, N.; Reuter, C.; Boratynski, T.; Vacek, D. Molecular Diagnosis of Populational Variants of Anthonomus Grandis (Coleoptera: Curculionidae) in North America. J. Econ. Entomol. 2013, 106, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Roehrdanz, R.L. Genetic Differentiation of Southeastern Boll Weevil and Thurberia Weevil Populations of Anthonomus Grandis (Coleoptera: Curculionidae) Using Mitochondrial DNA. Ann. Entomol. Soc. Am. 2001, 94, 928–935. [Google Scholar] [CrossRef]
- Tessnow, A.E.; Gilligan, T.M.; Burkness, E.; Bortoli, C.P.D.; Jurat-Fuentes, J.L.; Porter, P.; Sekula, D.; Sword, G.A. Novel Real-Time PCR Based Assays for Differentiating Fall Armyworm Strains Using Four Single Nucleotide Polymorphisms. PeerJ 2021, 9, e12195. [Google Scholar] [CrossRef]
- Livak, K.J.; Flood, S.J.; Marmaro, J.; Giusti, W.; Deetz, K. Oligonucleotides with Fluorescent Dyes at Opposite Ends Provide a Quenched Probe System Useful for Detecting PCR Product and Nucleic Acid Hybridization. Genome Res. 1995, 4, 357–362. [Google Scholar] [CrossRef]
- Raszick, T.J.; Perkin, L.C.; Shirley, X.A.; Ruiz-Arce, R.; Kramer, Z.A.; Suh, C.P.-C.; Sword, G.A. Source Tracing of Anthonomus Grandis Captured in Areas of the USA Where the Species Had Previously Been Eradicated. J. Pest. Sci. 2023, 1–11. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Rousset, F. Genepop’007: A Complete Re-Implementation of the Genepop Software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Perkin, L.C.; Oppert, B.; Duke, S.; Suh, C.P.-C. Assessment of DNA Integrity from Trap-Captured Boll Weevil (Coleoptera: Curculionidae) for Use in a New PCR-Based Diagnostic Tool. J. Econ. Entomol. 2021, 114, 1321–1328. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, C.; Sánchez, E.; Olivares, N.; Hinrichsen, P. Multiplex TaqMan Real-Time PCR Assay for Sensitive Detection of Two Weevil Species (Coleoptera: Curculionidae). J. Econ. Entomol. 2021, 114, 90–99. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Total Captures | A. g. grandis | A. g. thurberiae | Undet. A. grandis | Non-A. grandis |
|---|---|---|---|---|
| 1214 | 33 | 1036 | 134 | 11 |
| 3% | 85% | 11% | 1% |
| Forward Primer | 5′-CTGGCACTGTCGCGAATCTAT-3′ |
| Reverse Primer | 5′-ACGGACCGTTAGAAAAATACTTGGT-3′ |
| Reporter 1 Sequence | 5′-AAGCCGATCTTGCTAGTT-3′ |
| Reporter 2 Sequence | 5′-AAAGCCGATCTTACTAGTT-3′ |
| Context Sequence | 5′-GGCACTGTCGCGAATCTATAACTAG[C/T]AAGATCGGCTTTACCAAGTATTTTT-3′ |
| Subspecies | Locality | N | Target (DNA) (ng/µL) | Actual Range (DNA) (ng/µL) |
|---|---|---|---|---|
| A. g. grandis | ||||
| USA, Texas, Knippa | 3 | 0.05 | 0.029–0.252 | |
| USA, Texas La Feria | 2 | 0.05 | 0.018–0.172 | |
| USA, Texas, Knippa | 3 | 1 | 0.064–0.901 | |
| USA, Texas, La Feria | 2 | 1 | 0.538–0.828 | |
| USA, Texas, Knippa | 3 | 5 | 3.32–6.92 | |
| USA, Texas, La Feria | 2 | 5 | 2.76–5.94 | |
| USA, Texas, Knippa | 3 | 25 | 21.8–36.6 | |
| USA, Texas, La Feria | 2 | 25 | 16.6–32.2 | |
| A. g. thurberiae | ||||
| USA, New Mexico | 5 | 0.05 | 0.01–0.127 | |
| USA, New Mexico | 5 | 1 | 0.55–1.68 | |
| USA, New Mexico | 5 | 5 | 2.46–7.18 | |
| USA, New Mexico | 5 | 25 | 11.7–32.2 |
| Species | Common Name | Origin | N |
|---|---|---|---|
| A. eugenii | pepper weevil | USA, Florida | 3 |
| USA, Texas | 2 | ||
| A. fulvus | winecup weevil | USA, Texas | 5 |
| A. hunteri | n/a | Mexico, Campeche | 5 |
| A. musculus | cranberry weevil | USA, New Jersey | 4 |
| A. palmeri | n/a | Mexico, Chiapas | 5 |
| A. peninsularis | n/a | USA, Arizona | 2 |
| USA, California | 4 | ||
| A. texanus | n/a | Mexico, Chihuahua | 2 |
| USA, Texas | 3 | ||
| A. townsendi | n/a | Mexico, Chiapas | 5 |
| Conotrachelus nenuphar | plum curculio | USA, Oklahoma | 2 |
| USA, Vermont | 2 | ||
| USA, Texas | 1 | ||
| Curculio caryae | pecan weevil | USA, Texas | 5 |
| Rhyssomatus lineaticollis | milkweed stem weevil | USA, Wisconsin | 2 |
| USA, Texas | 2 | ||
| USA, Missouri | 1 |
| Subspecies | Country | Locality | Haplotype | |
|---|---|---|---|---|
| A. g. grandis | ||||
| Mexico | ||||
| Chihuahua | - | 2 | ||
| Chihuahua | AN4 | 2 | ||
| Chihuahua | AN11 | 1 | ||
| Chihuahua | AN12 | 25 | ||
| Coahuila | AN4 | 2 | ||
| Coahuila | AN12 | 4 | ||
| Coahuila | AN25 | 1 | ||
| Durango | AN4 | 1 | ||
| Durango | AN12 | 6 | ||
| Tamaulipas | - | 2 | ||
| Tamaulipas | AN1 | 5 | ||
| Tamaulipas | AN28 | 2 | ||
| Tamaulipas | AN30 | 1 | ||
| USA | ||||
| Mississippi | AN1 | 2 | ||
| Mississippi | AN9 | 1 | ||
| Mississippi | AN13 | 2 | ||
| New Mexico | AN1 | 1 | ||
| Texas | AN9 | 1 | ||
| Texas | AN10 | 1 | ||
| Texas | AN13 | 4 | ||
| Texas | AN27 | 7 | ||
| Texas | AN28 | 3 | ||
| Texas | AN31 | 1 | ||
| A. g. thurberiae | ||||
| Mexico | ||||
| Chihuahua | - | 1 | ||
| Chihuahua | AN4 | 2 | ||
| Chihuahua | AN8 | 1 | ||
| Chihuahua | AN14 | 2 | ||
| Chihuahua | AN15 | 1 | ||
| Chihuahua | AN16 | 4 | ||
| Chihuahua | AN17 | 1 | ||
| Chihuahua | AN18 | 1 | ||
| Chihuahua | AN19 | 2 | ||
| Chihuahua | AN20 | 1 | ||
| Chihuahua | AN21 | 1 | ||
| USA | ||||
| Arizona | AN2 | 2 | ||
| Arizona | AN4 | 1 | ||
| Arizona | AN8 | 3 | ||
| Arizona | AN17 | 8 | ||
| Arizona | AN18 | 2 | ||
| Arizona | AN19 | 10 | ||
| Arizona | AN20 | 3 | ||
| Arizona | AN22 | 1 | ||
| Arizona | AN23 | 1 | ||
| Arizona | AN24 | 2 | ||
| Arizona | AN26 | 2 |
| Subspecies | Country | Origin | N |
|---|---|---|---|
| A. g. grandis | |||
| Mexico | |||
| Chihuahua, Delicias | 13 | ||
| Chihuahua, Durango, Torreon | 5 | ||
| Chihuahua, Ojinaga | 8 | ||
| Chihuahua, Los Alamos | 4 | ||
| Coahuila, Ejido Chavez | 5 | ||
| Coahuila, San Pedro | 2 | ||
| Durango, Jimenez | 7 | ||
| Tamaulipas, Valle Hermoso | 10 | ||
| USA | |||
| Mississippi, Meyersville | 5 | ||
| New Mexico, Artesia | 1 | ||
| Texas, Rio Grande Valley | 17 | ||
| A. g. thurberiae | |||
| Mexico | |||
| Chihuahua, Agua Prieta | 17 | ||
| USA | |||
| Arizona, Bisbee | 1 | ||
| Arizona, East of Sasabe | 12 | ||
| Arizona, Kitt Peak | 9 | ||
| Arizona, Phoenix | 5 | ||
| Arizona, Sonoita | 4 | ||
| Arizona, Chiricahua, Mts. | 4 |
| Subspecies and Origin | IMMDL | TAMU | ICCDRU | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | BW | TW | Inc. | BW | TW | Inc. | BW | TW | Inc. | PCR Success | Accuracy | |
| A. g. grandis | ||||||||||||
| Mexico | ||||||||||||
| Chihuahua | 30 | 25 | 0 | 5 | 27 | 0 | 3 | 24 | 0 | 5 | 88.00% | 100% |
| Coahuila | 7 | 5 | 0 | 2 | 7 | 0 | 0 | 5 | 0 | 2 | 79.50% | 100% |
| Durango | 7 | 6 | 0 | 1 | 7 | 0 | 0 | 5 | 0 | 0 | 91.30% | 100% |
| Tamaulipas | 10 | 10 | 0 | 0 | 10 | 0 | 0 | 10 | 0 | 0 | 95.70% | 100% |
| USA | ||||||||||||
| Mississippi | 5 | 3 | 0 | 2 | 4 | 0 | 1 | 3 | 0 | 1 | 68.80% | 100% |
| New, Mexico | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 85.70% | 100% |
| Texas | 17 | 12 | 0 | 5 | 14 | 0 | 3 | 3 | 2 | 5 | 74.30% | 95.50% |
| A. g. thurberiae | ||||||||||||
| Mexico | ||||||||||||
| Chihuahua | 17 | 0 | 14 | 3 | 0 | 16 | 1 | 1 | 14 | 2 | 90.80% | 98.00% |
| USA | ||||||||||||
| Arizona | 35 | 0 | 28 | 7 | 0 | 33 | 2 | 0 | 28 | 7 | 90.30% | 100% |
| Subspecies and Origin | IMMDL | TAMU | ICCDRU | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N | BW | TW | Inc. | BW | TW | Inc. | BW | TW | Inc. | Accuracy | |
| A. g. grandis (BW) | |||||||||||
| Mexico | |||||||||||
| Chihuahua, Delicias | 13 | 9 | 0 | 4 | 10 | 0 | 3 | 10 | 0 | 3 | 100% |
| Chihuahua, Durango, Torreon | 5 | 5 | 0 | 0 | 5 | 0 | 0 | 4 | 0 | 0 | 100% |
| Chihuahua, Ojinaga | 8 | 8 | 0 | 0 | 8 | 0 | 0 | 7 | 0 | 0 | 100% |
| Chihuahua, Los Alamos | 4 | 3 | 0 | 1 | 4 | 0 | 0 | 3 | 0 | 1 | 100% |
| Coahuila, Ejido Chavez | 5 | 3 | 0 | 2 | 5 | 0 | 0 | 3 | 0 | 0 | 100% |
| Coahuila, San Pedro | 2 | 2 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 100% |
| Durango, Jimenez | 7 | 6 | 0 | 1 | 7 | 0 | 0 | 6 | 0 | 0 | 100% |
| Tamaulipas, Valle Hermoso | 10 | 10 | 0 | 0 | 10 | 0 | 0 | 10 | 0 | 0 | 100% |
| USA | |||||||||||
| Mississippi, Meyersville | 5 | 3 | 0 | 2 | 4 | 0 | 1 | 3 | 0 | 1 | 100% |
| New Mexico, Artesia | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 100% |
| Texas, Rio Grande Valley | 17 | 12 | 0 | 5 | 14 | 0 | 3 | 5 | 2 | 3 | 95.50% |
| A. g. thurberiae (TW) | |||||||||||
| Mexico | |||||||||||
| Chihuahua, Agua Prieta | 17 | 14 | 0 | 3 | 16 | 0 | 1 | 14 | 1 | 2 | 98.00% |
| USA | |||||||||||
| Arizona, Bisbee | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 100% |
| Arizona, East of Sasabe | 12 | 12 | 0 | 0 | 12 | 0 | 0 | 12 | 0 | 0 | 100% |
| Arizona, Kitt Peak | 9 | 4 | 0 | 5 | 7 | 0 | 2 | 6 | 0 | 3 | 100% |
| Arizona, Phoenix | 5 | 4 | 0 | 1 | 5 | 0 | 0 | 3 | 0 | 2 | 100% |
| Arizona, Sonoita | 4 | 4 | 0 | 0 | 4 | 0 | 0 | 2 | 0 | 0 | 100% |
| Arizona, Chiricahua, Mts. | 4 | 3 | 0 | 1 | 4 | 0 | 0 | 4 | 0 | 0 | 100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raszick, T.J.; Perkin, L.C.; Godoy, A.; Shirley, X.A.; Wright, K.; Martin, P.T.; Suh, C.P.-C.; Ruiz-Arce, R.; Sword, G.A. A New qPCR Assay for the Rapid Diagnosis of Anthonomus grandis Subspecies. Insects 2023, 14, 845. https://doi.org/10.3390/insects14110845
Raszick TJ, Perkin LC, Godoy A, Shirley XA, Wright K, Martin PT, Suh CP-C, Ruiz-Arce R, Sword GA. A New qPCR Assay for the Rapid Diagnosis of Anthonomus grandis Subspecies. Insects. 2023; 14(11):845. https://doi.org/10.3390/insects14110845
Chicago/Turabian StyleRaszick, Tyler Jay, Lindsey C. Perkin, Alejandra Godoy, Xanthe A. Shirley, Karen Wright, Paxton T. Martin, Charles P. -C. Suh, Raul Ruiz-Arce, and Gregory A. Sword. 2023. "A New qPCR Assay for the Rapid Diagnosis of Anthonomus grandis Subspecies" Insects 14, no. 11: 845. https://doi.org/10.3390/insects14110845