
Mosquito Larvicidal Activity and Cytotoxicity of the Extracts of Aromatic Plants from Malaysia


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
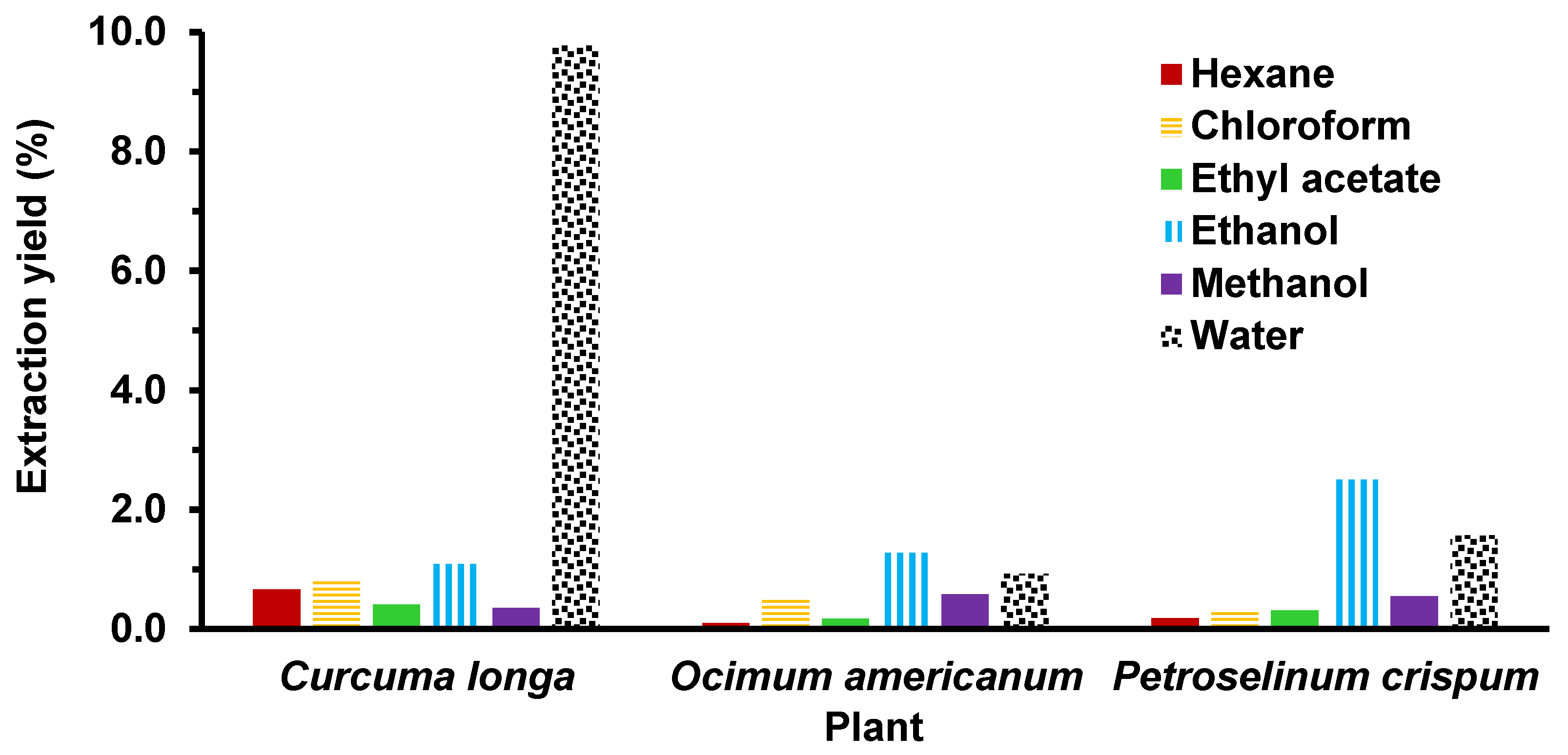
2.2. Preparation of Plant Extracts
2.3. Mosquito Sampling and Larvae Culturing
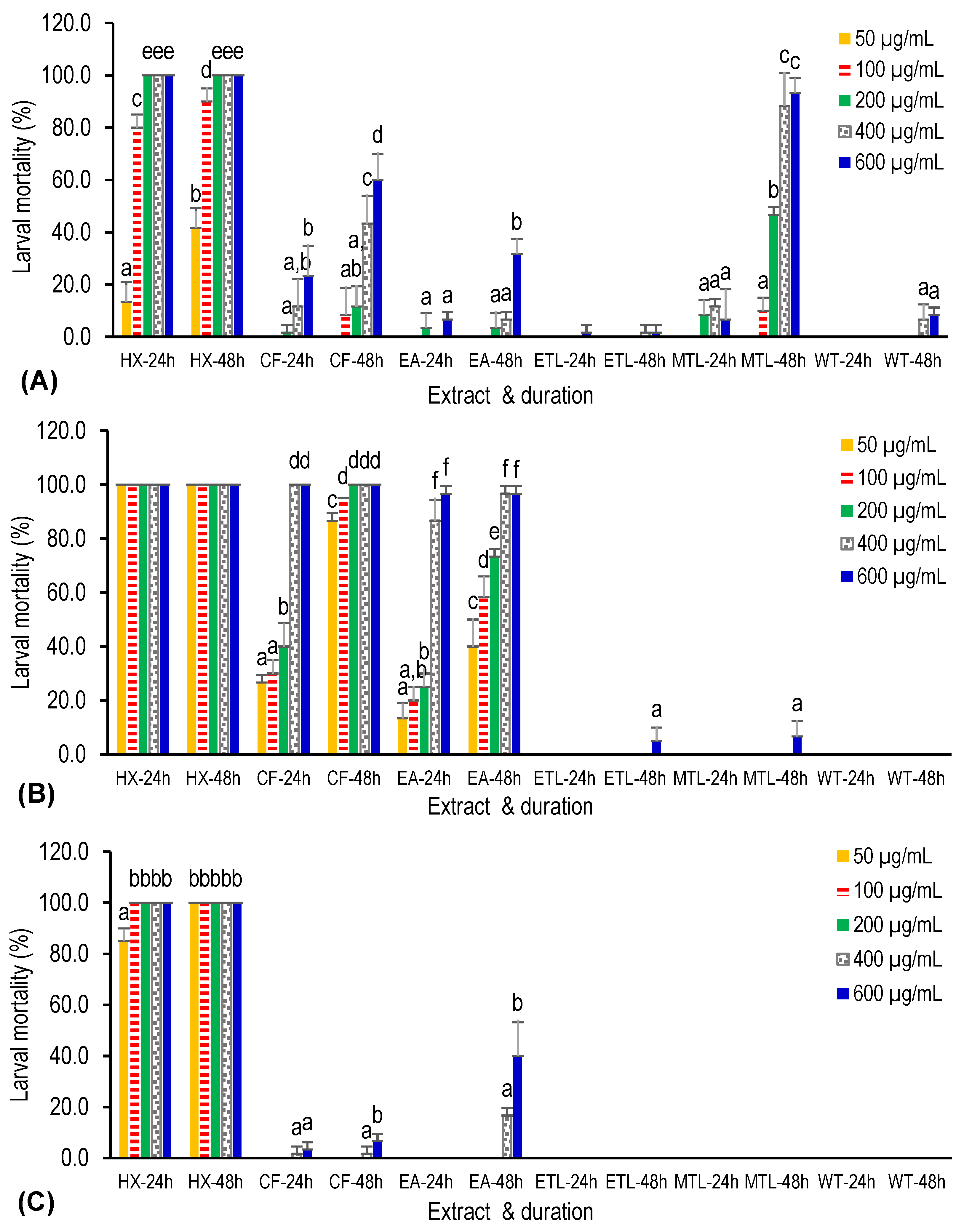
2.4. Larvicidal Bioassay
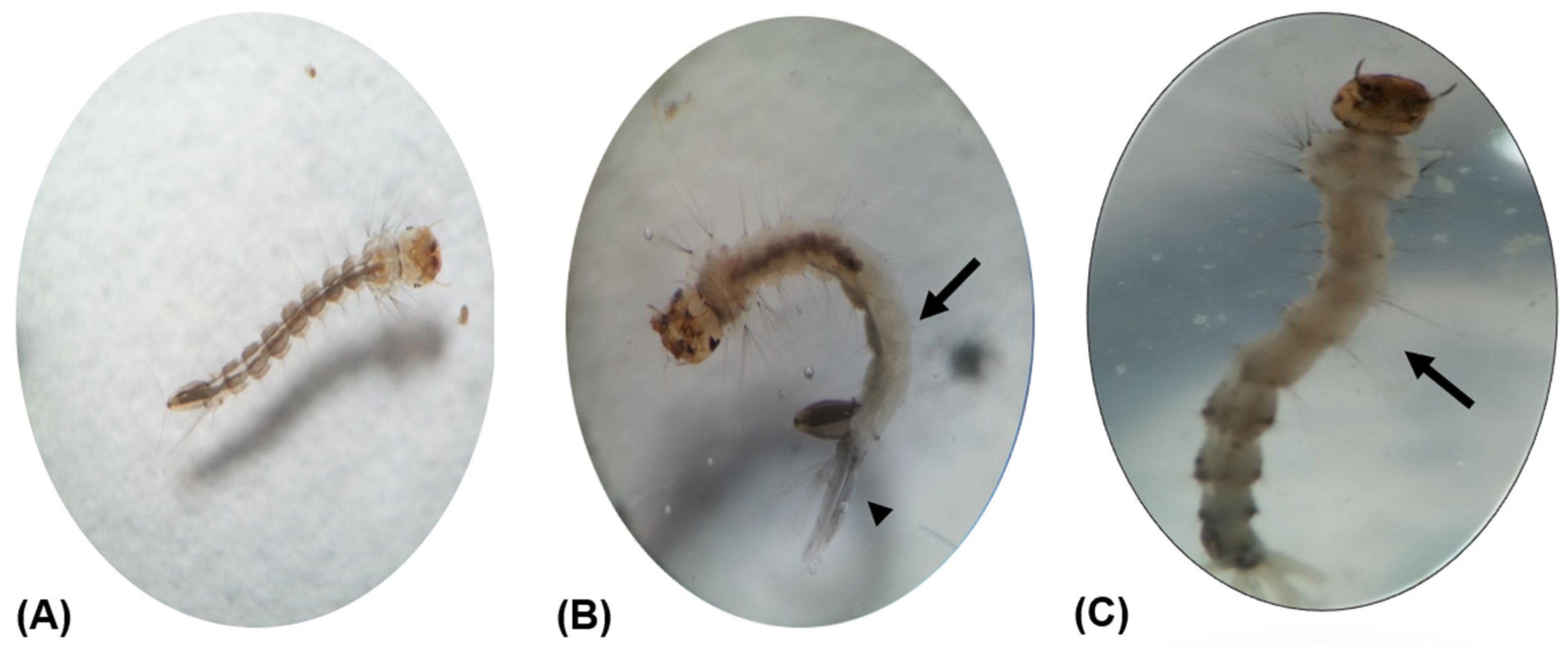
2.5. Larvae Morphological Examination
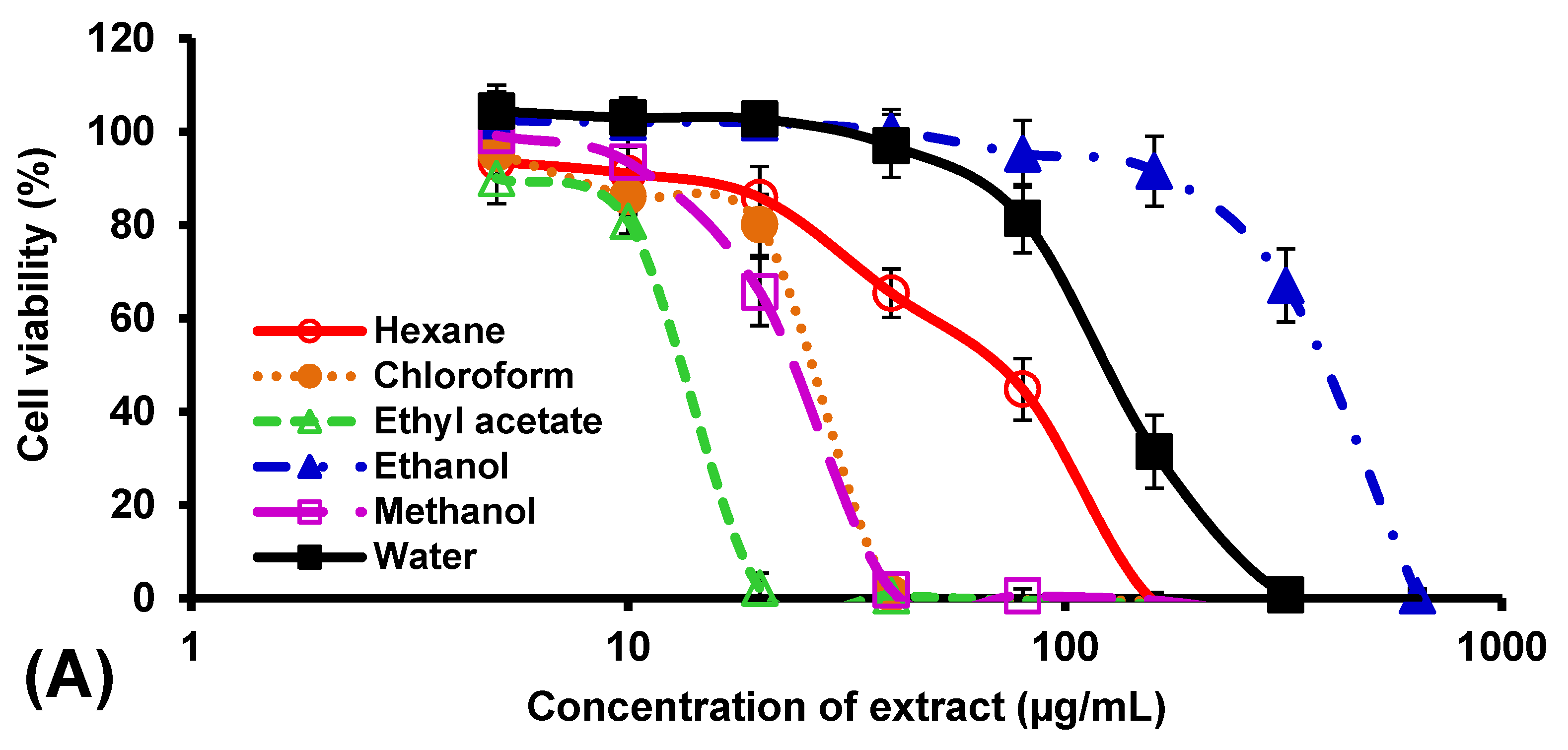
2.6. Cytotoxic Activity
2.7. Profiling of Metabolites Using Gas Chromatography–Mass Spectrometry (GC–MS)
2.8. Data Analysis
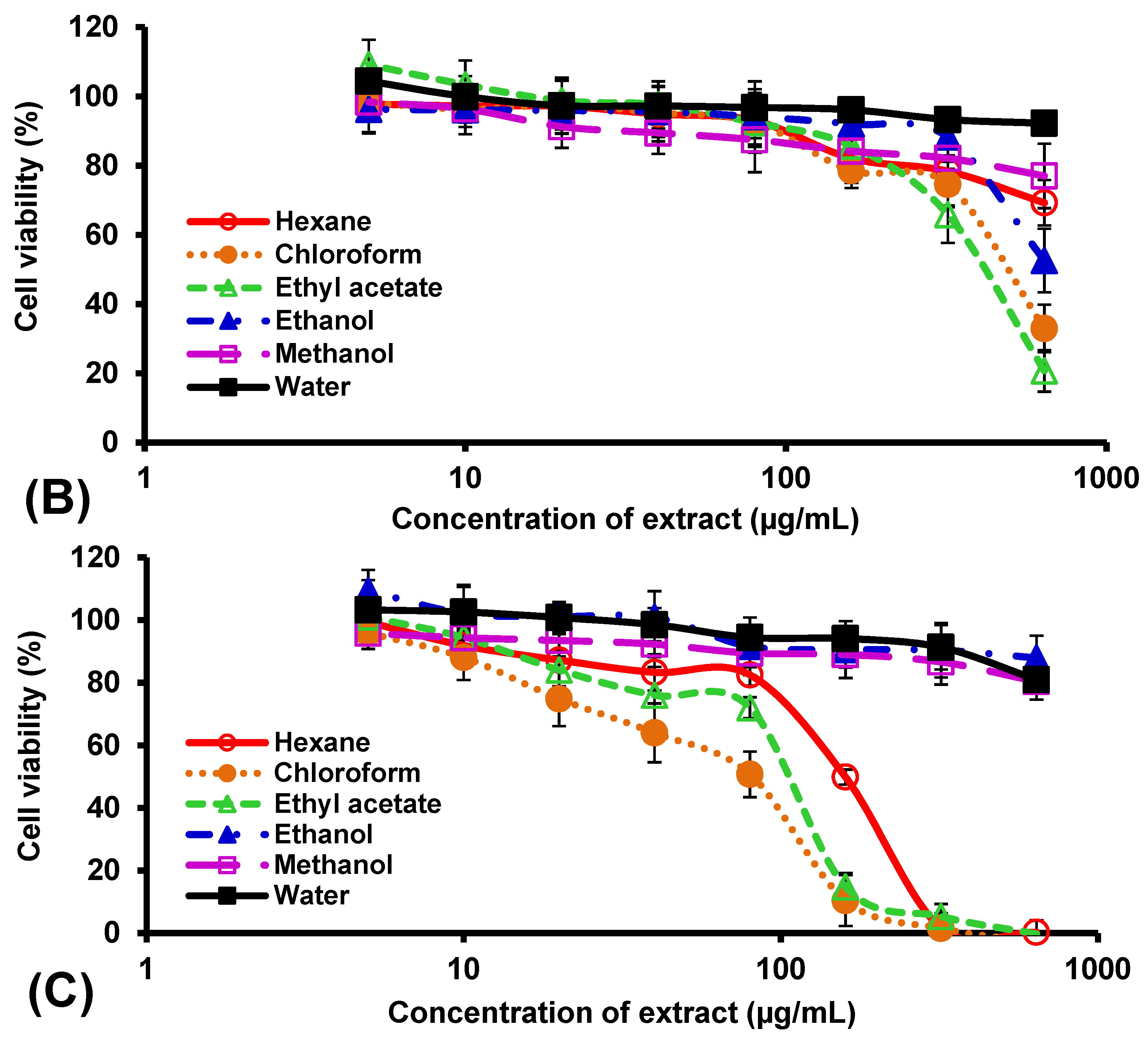
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kraemer, M.U.G.; Reiner, R.C., Jr.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef]
- Lee, H.; Halverson, S.; Ezinwa, N. Mosquito-borne diseases. Prim. Care 2018, 45, 393–407. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Vector-Borne Diseases. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/vector-borne-diseases (accessed on 1 March 2023).
- Silva, N.M.; Santos, N.C.; Martins, I.C. Dengue and Zika viruses: Epidemiological history, potential therapies, and promising vaccines. Trop. Med. Infect. Dis. 2020, 5, 150. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.P.G.; Baptista, S.S.S.G.; Sousa, C.A.G.C.C.; Novo, M.T.L.M.; Ramos, H.C.; Panella, N.A.; Godsey, M.; Simões, M.J.; Anselmo, M.L.; Komar, N.; et al. Bioecology and vectorial capacity of Aedes albopictus (Diptera: Culicidae) in Macao, China, in relation to dengue virus transmission. J. Med. Entomol. 2005, 42, 419–428. [Google Scholar] [CrossRef]
- Gould, E.A.; Higgs, S. Impact of climate change and other factors on emerging arbovirus diseases. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 109–121. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention. Development of Aedes albopictus Risk Maps. 2009. Available online: https://www.ecdc.europa.eu/en/publications-data/development-aedes-albopictus-risk-maps (accessed on 28 February 2023).
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, an arbovirus vector: From the darkness to the light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Wilson, A.L.; Courtenay, O.; Kelly-Hope, L.A.; Scott, T.W.; Takken, W.; Torr, S.J.; Lindsay, S.W. The importance of vector control for the control and elimination of vector-borne diseases. PLoS Negl. Trop. Dis. 2020, 14, e0007831. [Google Scholar] [CrossRef]
- Jones, R.T.; Ant, T.H.; Cameron, M.M.; Logan, J.G. Novel control strategies for mosquito-borne diseases. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, 20190802. [Google Scholar] [CrossRef]
- Protopopoff, N.; Mosha, J.F.; Lukole, E.; Charlwood, J.D.; Wright, A.; Mwalimu, C.D.; Manjurano, A.; Mosha, F.W.; Kisinza, W.; Kleinschmidt, I.; et al. Effectiveness of a long-lasting piperonyl butoxide-treated insecticidal net and indoor residual spray interventions, separately and together, against malaria transmitted by pyrethroid-resistant mosquitoes: A cluster, randomised controlled, two-by-two factorial design trial. Lancet 2018, 391, 1577–1588. [Google Scholar]
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Annu. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Sarma, N.; Patouillard, E.; Cibulskis, R.E.; Arcand, J.L. The economic burden of malaria: Revisiting the evidence. Am. J. Trop. Med. Hyg. 2019, 101, 1405–1415. [Google Scholar] [CrossRef] [PubMed]
- Moyes, C.L.; Vontas, J.; Martins, A.J.; Ng, L.C.; Koou, S.Y.; Dusfour, I.; Raghavendra, K.; Pinto, J.; Corbel, V.; David, J.P.; et al. Contemporary status of insecticide resistance in the major Aedes vectors of arboviruses infecting humans. PLoS Negl. Trop. Dis. 2017, 11, e0005625. [Google Scholar] [CrossRef] [PubMed]
- Weetman, D.; Kamgang, B.; Badolo, A.; Moyes, C.L.; Shearer, F.M.; Coulibaly, M.; Pinto, J.; Lambrechts, L.; McCall, P.J. Aedes mosquitoes and Aedes-borne arboviruses in Africa: Current and future threats. Int. J. Environ. Res. Public Health 2018, 15, 220. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, M.; Fu, W.-J.; Hu, J.-F.; Dai, G.-H. Efficacy of bioactive compounds from Curcuma longa against mosquito larvae. J. Appl. Entomol. 2018, 142, 792–799. [Google Scholar] [CrossRef]
- Ghosh, A.; Chowdhury, N.; Chandra, G. Plant extracts as potential mosquito larvicides. Indian J. Med. Res. 2012, 135, 581–598. [Google Scholar]
- Ajaiyeoba, E.O.; Sama, W.; Essien, E.E.; Olayemi, J.O.; Ekundayo, O.; Walker, T.M.; Setzer, W.N. Larvicidal activity of turmerone-rich essential oils of Curcuma longa leaf and rhizome from Nigeria on Anopheles gambiae. Pharm. Biol. 2008, 46, 279–282. [Google Scholar] [CrossRef]
- Craig, W.J. Health-promoting properties of common herbs. Am. J. Clin. Nutr. 1999, 70, 491S–499S. [Google Scholar] [CrossRef]
- Bansal, R.P.; Bahl, J.R.; Garg, S.N.; Naqvi, A.A.; Kumar, S. Differential chemical compositions of the essential oils of the shoot organs, rhizomes and rhizoids in the turmeric Curcuma longa grown in Indo-Gangetic Plains. Pharm. Biol. 2002, 40, 384–389. [Google Scholar] [CrossRef]
- Chane-Ming, J.; Vera, R.; Chalchat, J.C.; Cabassu, P. Chemical composition of essential oils from rhizomes, leaves and flowers of Curcuma longa L. from Reunion Island. J. Essent. Oil Res. 2002, 14, 249–251. [Google Scholar] [CrossRef]
- Sakuntala, B.; Sahoo, S.; Srivastava, V.K.; Behura, S. Major constituents in leaf essential oils of Curcuma longa L. and Curcuma aromatic Salisb. Curr. Sci. 2002, 83, 1312–1313. [Google Scholar]
- Martins, A.P.; Salgueiro, L.; Gonçalves, M.J.; Cunha, A.P.; Vila, R.; Cañigueral, S.; Mazzoni, V.; Tomi, F.; Casanova, J. Essential oil composition and antimicrobial activity of three Zingiberaceae from Sao Tome and Principe. Planta Med. 2001, 67, 580–584. [Google Scholar] [CrossRef]
- Orellana-Paucar, A.M.; Machado-Orellana, M.G. Pharmacological profile, bioactivities, and safety of turmeric oil. Molecules 2022, 27, 5055. [Google Scholar] [CrossRef] [PubMed]
- Pandian, R.S.; Kathiresan, K. Repellent activity of the rhizome of Curcuma longa Linn. against nocturnal mosquitoes in a rural area. Insect Environ. 2002, 8, 127–129. [Google Scholar]
- Singh, R.; Chandra, R.; Bose, M.; Luthra, P.M. Antibacterial activity of Curcuma longa rhizome extract on pathogenic bacteria. Curr. Sci. 2002, 83, 737–740. [Google Scholar]
- Aggarwal, B.B.; Kumar, A.; Bharti, A.C. Anticancer potential of curcumin: Preclinical and clinical studies. Anticancer Res. 2003, 23, 363–398. [Google Scholar]
- Sacchetti, G.; Maietti, S.; Muzzoli, M.; Scaglianti, M.; Manfredini, S.; Radice, M.; Bruni, R. Comparative evaluation of 11 essential oils of different origins as functional antioxidants, antiradicals and antimicrobials in foods. Food Chem. 2005, 91, 621–632. [Google Scholar] [CrossRef]
- Tripithi, A.K.; Prajapati, V.; Verma, N.; Bahl, J.R.; Bansal, R.P.; Khanuja, S.P.S.; Kumar, S. Bioactivities of the leaf essential oil of Curcuma longa (Var. Ch-66) on three species of stored-product beetles (Coleoptera). J. Econ. Entomol. 2002, 95, 183–189. [Google Scholar] [CrossRef]
- Güneri, N. A review on turmeric (Curcuma longa L.) and usage in seafood. Mar. Sci. Technol. Bull. 2021, 10, 71–84. [Google Scholar] [CrossRef]
- Zhu, J.; Zeng, X.; O’Neal, M.; Schultz, G.; Tucker, B.; Coats, J.; Bartholomay, L.; Xue, R.-D. Mosquito larvicidal activity of botanical-based mosquito repellents. J. Am. Mosq. Control Assoc. 2008, 24, 161–168. [Google Scholar] [CrossRef]
- Das, N.G.; Dhiman, S.; Talukdar, P.K.; Rabha, B.; Goswami, D.; Veer, V. Synergistic mosquito-repellent activity of Curcuma longa, Pogostemon heyneanus and Zanthoxylum limonella essential oils. J. Infect. Public Health 2015, 8, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Singha, S.; Chandra, G. Mosquito larvicidal activity of some common spices and vegetable waste on Culex quinquefasciatus and Anopheles stephensi. Asian Pac. J. Trop. Med. 2011, 4, 288–293. [Google Scholar] [CrossRef]
- Madhu, S.K.; Shaukath, A.K.; Vijayan, V.A. Efficacy of bioactive compounds from Curcuma aromatica against mosquito larvae. Acta Trop. 2010, 113, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Sukumar, K.; Perich, M.J.; Boobar, L.R. Botanical derivatives in mosquito control: A review. J. Am. Mosq. Control Assoc. 1991, 7, 210–237. [Google Scholar]
- Flora and Fauna Web. Ocimum americanum. 2022. Available online: https://www.nparks.gov.sg/florafaunaweb/flora/5/0/5057 (accessed on 1 March 2023).
- Ali, H.M.; Nguta, J.M.; Mapenay, I.O.; Musila, F.M.; Omambia, V.M.; Matara, D.N. Ethnopharmacological uses, biological activities, chemistry and toxicological aspects of Ocimum americanum var. americanum (Lamiaceae). J. Phytopharm. 2021, 10, 56–60. [Google Scholar] [CrossRef]
- Ingole, S.N. Phytochemical analysis of leaf extract of Ocimum americanum L. (Lamiaceae) by GCMS method. World Sci. News 2016, 37, 76–87. [Google Scholar]
- Sutili, F.J.; Velasquez, A.; Pinheiro, C.G.; Heinzmann, B.M.; Gatlin III, D.M.; Baldisserotto, B. Evaluation of Ocimum americanum essential oil as an additive in red drum (Sciaenops ocellatus) diets. Fish Shellfish Immunol. 2016, 56, 155–161. [Google Scholar] [CrossRef]
- Zengin, G.; Ferrante, C.; Gnapi, D.E.; Sinan, K.I.; Orlando, G.; Recinella, L.; Diuzheva, A.; Jekő, J.; Cziáky, Z.; Chiavarol, A.; et al. Comprehensive approaches on the chemical constituents and pharmacological properties of flowers and leaves of American basil (Ocimum americanum L). Food Res. Int. 2019, 125, 108610. [Google Scholar] [CrossRef]
- Vidhya, E.; Vijayakumar, S.; Rajalakshmi, S.; Kalaiselvi, S.; Pandiyan, P. Antimicrobial activity and phytochemical screening of Ocimum americanum L. extracts against pathogenic microorganisms. Acta Ecol. Sin. 2020, 40, 214–220. [Google Scholar]
- Avetisyan, A.; Markosian, A.; Petrosyan, M.; Sahakyan, N.; Babayan, A.; Aloyan, S.; Trchounian, A. Chemical composition and some biological activities of the essential oils from basil Ocimum different cultivars. BMC Complement. Altern. Med. 2017, 17, 60. [Google Scholar] [CrossRef]
- El-Aziz, S.E.A.; Omer, E.A.; Sabra, A.S. Chemical composition of Ocimum americanum essential oil and its biological effects against Agrotis ipsilon (Lepidoptera: Noctuidae). Res. J. Agric. Biol. Sci. 2007, 3, 740–747. [Google Scholar]
- Carović-Stanko, K.; Orlić, S.; Politeo, O.; Strikić, F.; Kolak, I.; Milos, M.; Satovic, Z. Composition and antibacterial activities of essential oils of seven Ocimum taxa. Food Chem. 2010, 119, 196–201. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Vidhya, E.; Anand, G.C.; Nilavukkarasi, M.; Punitha, V.N.; Sakthivel, B. Eco friendly synthesis of TiO2 nanoparticles using aqueous Ocimum americanum L. leaf extracts and their antimicrobial, anti-proliferative and photocatalytic activities. Vegetos 2020, 33, 805–810. [Google Scholar] [CrossRef]
- Chokechaijaroenporn, O.; Bunyapraphatsara, N.; Kongchuensin, S. Mosquito repellent activities of Ocimum volatile oils. Phytomedicine 1994, 1, 135–139. [Google Scholar] [CrossRef]
- Krüger, H.; Wetzel, S.B.; Zeiger, B. The chemical variability of Ocimum species. J. Herbs Spices Med. Plants 2002, 9, 335–344. [Google Scholar] [CrossRef]
- Narayanan, M.; Vijay, A.; Kandasamy, S.; Nasif, O.; Alharbi, S.A.; Srinivasan, R.; Kavitha, R. Phytochemical profile and larvicidal activity of aqueous extract of Ocimum americanum against mosquito vectors. Appl. Nanosci. 2021, 11, 3369–3381. [Google Scholar] [CrossRef]
- Farzaei, M.H.; Abbasabadi, Z.; Ardekani, M.R.S.; Rahimi, R.; Fatemeh, F. Parsley: A review of ethnopharmacology, phytochemistry and biological activities. J. Tradit. Chin. Med. 2013, 33, 815–826. [Google Scholar] [CrossRef]
- Blumenthal, M.; Goldberg, A.; Brinckmann, J. Herbal Medicine: Expanded Commission E Monographs, 1st ed.; American Botanical Council: Austin, TX, USA, 2000; p. 519. [Google Scholar]
- Linde, G.A.; Gazim, Z.C.; Cardoso, B.K.; Jorge, L.F.; Tešević, V.; Glamoćlija, J.; Soković, M.; Colauto, N.B. Antifungal and antibacterial activities of Petroselinum crispum essential oil. Genet. Mol. Res. 2016, 15, gmr.15038538. [Google Scholar] [CrossRef]
- Moazedi, A.A.; Mirzaie, D.N.; Seyyednejad, S.M.; Zadkarami, M.R.; Amirzargar, A. Spasmolytic effect of Petroselinum crispum (Parsley) on rat’s ileum at different calcium chloride concentrations. Pak. J. Biol. Sci. 2007, 10, 4036–4042. [Google Scholar] [CrossRef]
- Behtash, N.; Kargarzadeh, F.; Shafaroudi, H. Analgesic effects of seed extract from Petroselinum crispum (Tagetes minuta) in animal models. Toxicol. Lett. 2008, 180, S127–S128. [Google Scholar] [CrossRef]
- Pápay, Z.E.; Kósa, A.; Boldizsár, I.; Ruszkai, A.; Balogh, E.; Klebovich, I.; Antal, I. Pharmaceutical and formulation aspects of Petroselinum crispum extract. Acta Pharm. Hung. 2012, 82, 3–14. [Google Scholar] [PubMed]
- Aljanaby, A.A.J.J. Antibacterial activity of an aqueous extract of Petroselinum crispum leaves against pathogenic bacteria isolated from patients with burns infections in Al-Najaf Governorate, Iraq. Res. Chem. Intermed. 2013, 39, 3709–3714. [Google Scholar] [CrossRef]
- Agyare, C.; Appiah, T.; Boakye, Y.D.; Apenteng, J.A. Petroselinum crispum: A review. In Medicinal Spices and Vegetables from Africa. Therapeutic Potential against Metabolic, Inflammatory, Infectious and Systemic Diseases; Kuete, V., Ed.; Elsevier: London, UK, 2017; pp. 527–547. [Google Scholar]
- Zhang, H.; Chen, F.; Wang, X.; Yao, H.-Y. Evaluation of antioxidant activity of parsley (Petroselinum crispum) essential oil and identification of its antioxidant constituents. Food Res. Int. 2006, 39, 833–839. [Google Scholar] [CrossRef]
- Sbai, H.; Saad, I.; Ghezal, N.; Greca, M.D.; Haouala, R. Bioactive compounds isolated from Petroselinum crispum L. leaves using bioguided fractionation. Ind. Crops Prod. 2016, 89, 207–214. [Google Scholar] [CrossRef]
- Intirach, J.; Junkum, A.; Lumjuan, N.; Chaithong, U.; Somboon, P.; Jitpakdi, A.; Riyong, D.; Wannasan, A.; Champakaew, D.; Muangmoon, R.; et al. Biochemical effects of Petroselinum crispum (Umbellifereae) essential oil on the pyrethroid resistant strains of Aedes aegypti (Diptera: Culicidae). Insects 2019, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Piplani, M.; Bhagwat, D.P.; Singhvi, G.; Sankaranarayanan, M.; Balana-Fouce, R.; Vats, T.; Chander, S. Plant-based larvicidal agents: An overview from 2000 to 2018. Exp. Parasitol. 2019, 199, 92–103. [Google Scholar] [CrossRef]
- Junkum, A.; Intirach, J.; Chansang, A.; Champakaew, D.; Chaithong, U.; Jitpakdi, A.; Riyong, D.; Somboon, P.; Pitasawat, B. Enhancement of temephos and deltamethrin toxicity by Petroselinum crispum oil and its main constituents against Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2021, 58, 1298–1315. [Google Scholar] [CrossRef]
- Seghier, H.; Tine-Djebbar, F.; Loucif-Ayad, W.; Soltani, N. Insecticidal activity of Petroselinum crispum essential oil on mosquitoes. J. Entomol. Res. 2020, 44, 613–620. [Google Scholar] [CrossRef]
- Knio, K.M.; Usta, J.; Dagher, S.; Zournajian, H.; Kreydiyyeh, S. Larvicidal activity of essential oils extracted from commonly used herbs in Lebanon against the seaside mosquito, Ochlerotatus caspius. Bioresour. Technol. 2008, 99, 763–768. [Google Scholar] [CrossRef]
- World Flora Online Consortium. World Flora Online. 2021. Available online: http://www.worldfloraonline.org (accessed on 4 November 2021).
- Chan, S.M.; Khoo, K.S.; Sekaran, S.D.; Sit, N.W. Mode-dependent antiviral activity of medicinal plant extracts against the mosquito-borne chikungunya virus. Plants 2021, 10, 1658. [Google Scholar] [CrossRef]
- Rueda, L.M. Pictorial keys for the identification of mosquitoes (Diptera: Culicidae) associated with dengue virus transmission. Zootaxa 2004, 589, 1–60. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for Laboratory and Field Testing of Mosquito Larvicides. 2005. Available online: https://apps.who.int/iris/handle/10665/69101 (accessed on 19 August 2014).
- Chan, S.M.; Khoo, K.S.; Sit, N.W. Interactions between plant extracts and cell viability indicators during cytotoxicity testing: Implications for ethnopharmacological studies. Trop. J. Pharm. Res. 2015, 14, 1991–1998. [Google Scholar] [CrossRef]
- Repetto, G.; del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/ cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Snyder, L.R. Classification of the solvent properties of common liquids. J. Chromatogr. 1978, 92, 223–234. [Google Scholar] [CrossRef]
- Kalaivani, K.; Senthil-Nathan, S.; Murugesan, A.G. Biological activity of selected Lamiaceae and Zingiberaceae plant essential oils against the dengue vector Aedes aegypti L. (Diptera: Culicidae). Parasitol. Res. 2012, 110, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Intirach, J.; Junkum, A.; Lumjuan, N.; Chaithong, U.; Jitpakdi, A.; Riyong, D.; Wannasan, A.; Champakaew, D.; Muangmoon, R.; Chansang, A.; et al. Antimosquito property of Petroselinum crispum (Umbellifereae) against the pyrethroid resistant and susceptible strains of Aedes aegypti (Diptera: Culicidae). Environ. Sci. Pollut. Res. 2016, 23, 23994–24008. [Google Scholar] [CrossRef]
- Prajapati, V.; Tripathi, A.K.; Aggarwal, K.K.; Khanuja, S.P.S. Insecticidal, repellent and oviposition-deterrent activity of selected essential oils against Anopheles stephensi, Aedes aegypti and Culex quinquefasciatus. Bioresour. Technol. 2005, 96, 1749–1757. [Google Scholar] [CrossRef]
- Roth, G.N.; Chandra, A.; Nair, M.G. Novel bioactivities of Curcuma longa constituents. J. Nat. Prod. 1998, 61, 542–545. [Google Scholar] [CrossRef]
- Cavalcanti, E.S.; Morais, S.M.; Lima, M.A.; Santana, E.W. Larvicidal activity of essential oils from Brazilian plants against Aedes aegypti L. Mem. Inst. Oswaldo Cruz. 2004, 99, 541–544. [Google Scholar] [CrossRef]
- Prabhavathi, O.; Yuvarajan, R.; Natarajan, D. Mosquitocidal properties of Ocimum canum Sims (Lamiaceae) leaf extracts against dengue vector Aedes aegypti L. (Diptera: Culicidae). J. Entomol. Acarol. Res. 2016, 48, 345–354. [Google Scholar] [CrossRef]
- Yaméogo, F.; Wangrawa, D.W.; Sombié, A.; Sanon, A.; Badolo, A. Insecticidal activity of essential oils from six aromatic plants against Aedes aegypti, dengue vector from two localities of Ouagadougou, Burkina Faso. Arthropod-Plant Interact. 2021, 15, 627–634. [Google Scholar] [CrossRef]
- Pavela, R. Essential oils for the development of eco-friendly mosquito larvicides: A review. Ind. Crops Prod. 2015, 76, 174–187. [Google Scholar] [CrossRef]
- Yu, K.-X.; Wong, C.-L.; Ahmad, R.; Jantan, I. Mosquitocidal and oviposition repellent activities of the extracts of seaweed Bryopsis pennata on Aedes aegypti and Aedes albopictus. Molecules 2015, 20, 14082–14102. [Google Scholar] [CrossRef]
- Hussin, N.H.B.; Ismail, S.B.; Ke-Xin, Y. Larvicidal activity of ethanol extract of Carica papaya seed against Aedes albopictus (Skuse). Int. J. Medical Toxicol. Leg. Med. 2020, 23, 125–128. [Google Scholar] [CrossRef]
- Aziz, E.I.A.; Rahim, N.A.A.; Raduan, S.Z.; Safii, R. A preliminary study on larvicidal efficacy of Piper nigrum L. (Piperaceae) extracts against dengue vector, Aedes albopictus (Diptera: Culicidae). Serangga 2021, 26, 80–94. [Google Scholar]
- Chan, C.A.; Ho, L.Y.; Sit, N.W. Larvicidal activity and phytochemical profiling of sweet basil (Ocimum basilicum L.) leaf extract against Asian Tiger mosquito (Aedes albopictus). Horticulturae 2022, 8, 443. [Google Scholar] [CrossRef]
- David, J.P.; Rey, D.; Pautou, M.P.; Meyran, J.C. Differential toxicity of leaf litter to dipteran larvae of mosquito developmental sites. J. Invertebr. Pathol. 2000, 75, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Soonwera, M.; Phasomkusolsil, S. Effect of Cymbopogon citratus (lemongrass) and Syzygium aromaticum (clove) oils on the morphology and mortality of Aedes aegypti and Anopheles dirus larvae. Parasitol. Res. 2016, 115, 1691–1703. [Google Scholar] [CrossRef] [PubMed]
- Grover, M.; Behl, T.; Sehgal, A.; Singh, S.; Sharma, N.; Virmani, T.; Rachamalla, M.; Farasani, A.; Chigurupati, S.; Alsubayiel, A.M.; et al. In vitro phytochemical screening, cytotoxicity studies of Curcuma longa extracts with isolation and characterisation of their isolated compounds. Molecules 2021, 26, 7509. [Google Scholar] [CrossRef]
- Berrington, D.; Lall, N. Anticancer activity of certain herbs and spices on the cervical epithelial carcinoma (Hela) cell line. Evid. Based Complement. Altern. Med. 2012, 2012, 564927. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Sumner, B.W.; Sumner, L.W. Modern plant metabolomics for the discovery and characterization of natural products and their biosynthetic genes. In Comprehensive Natural Products III Chemistry and Biology; Liu, H.W., Begley, T.P., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands, 2020; Volume 7, pp. 156–188. [Google Scholar]
- Matasyoh, L.G.; Matasyoh, J.C.; Wachira, F.N.; Kinyua, M.G.; Thairu Muigai, A.W.; Mukiama, T.K. Antimicrobial activity of essential oils of Ocimum gratissimum L. from different populations of Kenya. Afr. J. Tradit. Complement. Altern. Med. 2008, 5, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Singh, P.; Tripathi, N.N. Chemistry and bioactivities of essential oils of some Ocimum species: An overview. Asian Pac. J. Trop. Biomed. 2014, 4, 682–694. [Google Scholar] [CrossRef]
- Perumalsamy, H.; Chang, K.S.; Park, C.; Ahn, Y.-J. Larvicidal activity of Asarum heterotropoides root constituents against insecticide-susceptible and-resistant Culex pipiens pallens and Aedes aegypti and Ochlerotatus togoi. J. Agric. Food Chem. 2010, 58, 10001–10006. [Google Scholar] [CrossRef] [PubMed]
- Rants’o, T.A.; Koekemoer, L.L.; van Zyl, R.L. In vitro and in silico analysis of the Anopheles anticholinesterase activity of terpenoids. Parasitol. Int. 2023, 93, 102713. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lin, M.; Jia, M.; Hu, J.; Zhu, L. Chemical composition and larvicidal activity against Aedes mosquitoes of essential oils from Arisaema fargesii. Pest Manag. Sci. 2020, 76, 534–542. [Google Scholar] [CrossRef]
- Cheng, S.-S.; Liu, J.-Y.; Huang, C.-G.; Hsui, Y.R.; Chen, W.-J.; Chang, S.-T. Insecticidal activities of leaf essential oils from Cinnamomum osmophloeum against three mosquito species. Bioresour. Technol. 2009, 100, 457–464. [Google Scholar] [CrossRef]
- Govindarajan, M.; Rajeswary, M.; Hoti, S.L.; Bhattacharyya, A.; Benelli, G. Eugenol, α-pinene and β-caryophyllene from Plectranthus barbatus essential oil as eco-friendly larvicides against malaria, dengue and Japanese encephalitis mosquito vectors. Parasitol. Res. 2016, 115, 807–815. [Google Scholar] [CrossRef]
- Bourdenx, B.; Bernard, A.; Domergue, F.; Pascal, S.; Léger, A.; Roby, D.; Pervent, M.; Vile, D.; Haslam, R.P.; Napier, J.A.; et al. Overexpression of Arabidopsis ECERIFERUM1 promotes wax very-long-chain alkane biosynthesis and influences plant response to biotic and abiotic stresses. Plant Physiol. 2011, 156, 29–45. [Google Scholar] [CrossRef]
- Farag, M.A.; Ezzat, S.M.; Salama, M.M.; Tadros, M.G. Anti-acetylcholinesterase potential and metabolome classification of 4 Ocimum species as determined via UPLC/qTOF/MS and chemometric tools. J. Pharm. Biomed. Anal. 2016, 125, 292–302. [Google Scholar] [CrossRef]
- Perumalsamy, H.; Jang, M.J.; Kim, J.-R.; Kadarkarai, M.; Ahn, Y.-J. Larvicidal activity and possible mode of action of four flavonoids and two fatty acids identified in Millettia pinnata seed toward three mosquito species. Parasites Vectors 2015, 8, 237. [Google Scholar] [CrossRef]
- Wang, Z.-Q.; Perumalsamy, H.; Wang, M.; Shu, S.; Ahn, Y.-J. Larvicidal activity of Magnolia denudata seed hydrodistillate constituents and related compounds and liquid formulations towards two susceptible and two wild mosquito species. Pest Manag. Sci. 2016, 72, 897–906. [Google Scholar] [CrossRef]
- Mohamed Abdoul-Latif, F.; Elmi, A.; Merito, A.; Nour, M.; Risler, A.; Ainane, A.; Bignon, J.; Ainane, T. Essential oils of Ocimum basilicum L. and Ocimum americanum L. from Djibouti: Chemical composition, antimicrobial and cytotoxicity evaluations. Processes 2022, 10, 1785. [Google Scholar] [CrossRef]
- Govindarajan, M.; Benelli, G. α-Humulene and β-elemene from Syzygium zeylanicum (Myrtaceae) essential oil: Highly effective and eco-friendly larvicides against Anopheles subpictus, Aedes albopictus, and Culex tritaeniorhynchus (Diptera: Culicidae). Parasitol. Res. 2016, 115, 2771–2778. [Google Scholar] [CrossRef]
- Cheng, S.-S.; Chua, M.-T.; Chang, E.-H.; Huang, C.-G.; Chen, W.-J.; Chang, S.-T. Variations in insecticidal activity and chemical compositions of leaf essential oils from Cryptomeria japonica at different ages. Bioresour. Technol. 2009, 100, 465–470. [Google Scholar] [CrossRef]
- Pereira Filho, A.A.; Pessoa, G.C.D.; Yamaguchi, L.F.; Stanton, M.A.; Serravite, A.M.; Pereira, R.H.M.; Neves, W.S.; Kato, M.J. Larvicidal activity of essential oils from Piper species against strains of Aedes aegypti (Diptera: Culicidae) resistant to pyrethroids. Front. Plant Sci. 2021, 12, 685864. [Google Scholar] [CrossRef] [PubMed]
- Theochari, I.; Giatropoulos, A.; Papadimitriou, V.; Karras, V.; Balatsos, G.; Papachristos, D.; Michaelakis, A. Physicochemical characteristics of four limonene-based nanoemulsions and their larvicidal properties against two mosquito species, Aedes albopictus and Culex pipiens molestus. Insects 2020, 11, 740. [Google Scholar] [CrossRef] [PubMed]
- Sanei-Dehkordi, A.; Moemenbellah-Fard, M.D.; Saffari, M.; Zarenezhad, E.; Osanloo, M. Nanoliposomes containing limonene and limonene-rich essential oils as novel larvicides against malaria and filariasis mosquito vectors. BMC Complement. Altern. Med. 2022, 22, 140. [Google Scholar] [CrossRef]
- Lozano-Grande, M.A.; Gorinstein, S.; Espitia-Rangel, E.; Dávila-Ortiz, G.; Martínez-Ayala, A.L. Plant sources, extraction methods, and uses of squalene. Int. J. Agron. 2018, 2018, 1829160. [Google Scholar] [CrossRef]
- Lou-Bonafonte, J.M.; Martínez-Beamonte, R.; Sanclemente, T.; Surra, J.C.; Herrera-Marcos, L.V.; Sanchez-Marco, J.; Arnal, C.; Osada, J. Current insights into the biological action of squalene. Mol. Nutr. Food Res. 2018, 62, 1800136. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Exposure Period (h) | LC50 (LCL-UCL) (µg/mL) | LC95 (LCL-UCL) (µg/mL) | Regression Coefficient (±Standard Error) | Chi-Square, X2 | Degree of Freedom, df | p-Value |
|---|---|---|---|---|---|---|---|
| Curcuma longa | |||||||
| Hexane | 24 | 74.89 (68.16–82.21) | 129.90 (113.39–159.84) | 6.878 ± 0.872 | 0.120 | 3 | 0.989 |
| 48 | 54.31 (46.24–61.19) | 115.87 (97.29–157.62) | 4.999 ± 0.820 | 0.185 | 3 | 0.980 | |
| Chloroform | 48 | 483.23 (403.82–613.49) | 2233.57 (1461.64–4464.62) | 2.474 ± 0.319 | 3.796 | 3 | 0.284 |
| Methanol | 48 | 214.32 (189.82–240.98) | 567.50 (475.36–720.79) | 3.889 ± 0.360 | 1.879 | 3 | 0.598 |
| Ocimum americanum | |||||||
| Hexane | 24 | 26.60 (20.63–35.53) | 51.39 (37.72–117.40) | 5.751 ± 0.656 | 15.568 | 5 | 0.008 |
| 48 | 0.90 | 7.75 | 1.761 ± 1.475 | 0.753 | 5 | 0.980 | |
| Chloroform | 24 | 143.04 | 650.19 | 2.501 ± 0.248 | 54.726 | 3 | 0.000 |
| 48 | 18.77 (1.87–32.55) | 85.30 (63.62–156.43) | 2.502 ± 0.813 | 0.817 | 3 | 0.845 | |
| Ethyl acetate | 24 | 198.42 (39.59–1338.13) | 808.17 (334.96–2.46 × 109) | 2.697 ± 0.253 | 27.869 | 3 | 0.000 |
| 48 | 74.78 (56.10–92.78) | 501.23 (365.13–813.97) | 1.991 ± 0.247 | 3.646 | 3 | 0.302 | |
| Petroselinum crispum | |||||||
| Hexane | 24 | 14.35 (0.54–28.62) | 83.66 (57.86–163.92) | 2.148 ± 0.736 | 3.110 | 3 | 0.375 |
| Extract | Median Cytotoxic Concentration, CC50 (µg/mL) | ||
|---|---|---|---|
| Plant | Curcuma longa | Ocimum americanum | Petroselinum crispum |
| Hexane | 72.7 ± 6.7 b | >640 | 159.3 ± 8.0 c |
| Chloroform | 26.7 ± 1.5 a | 509.1 ± 52.8 a | 82.7 ± 8.1 a |
| Ethyl acetate | 12.7 ± 0.6 a | 430.7 ± 40.1 a | 105.3 ± 4.5 b |
| Ethanol | 400.7 ± 33.5 d | >640 | >640 |
| Methanol | 23.3 ± 1.5 a | >640 | >640 |
| Water | 124.0 ± 9.5 c | - | >640 |
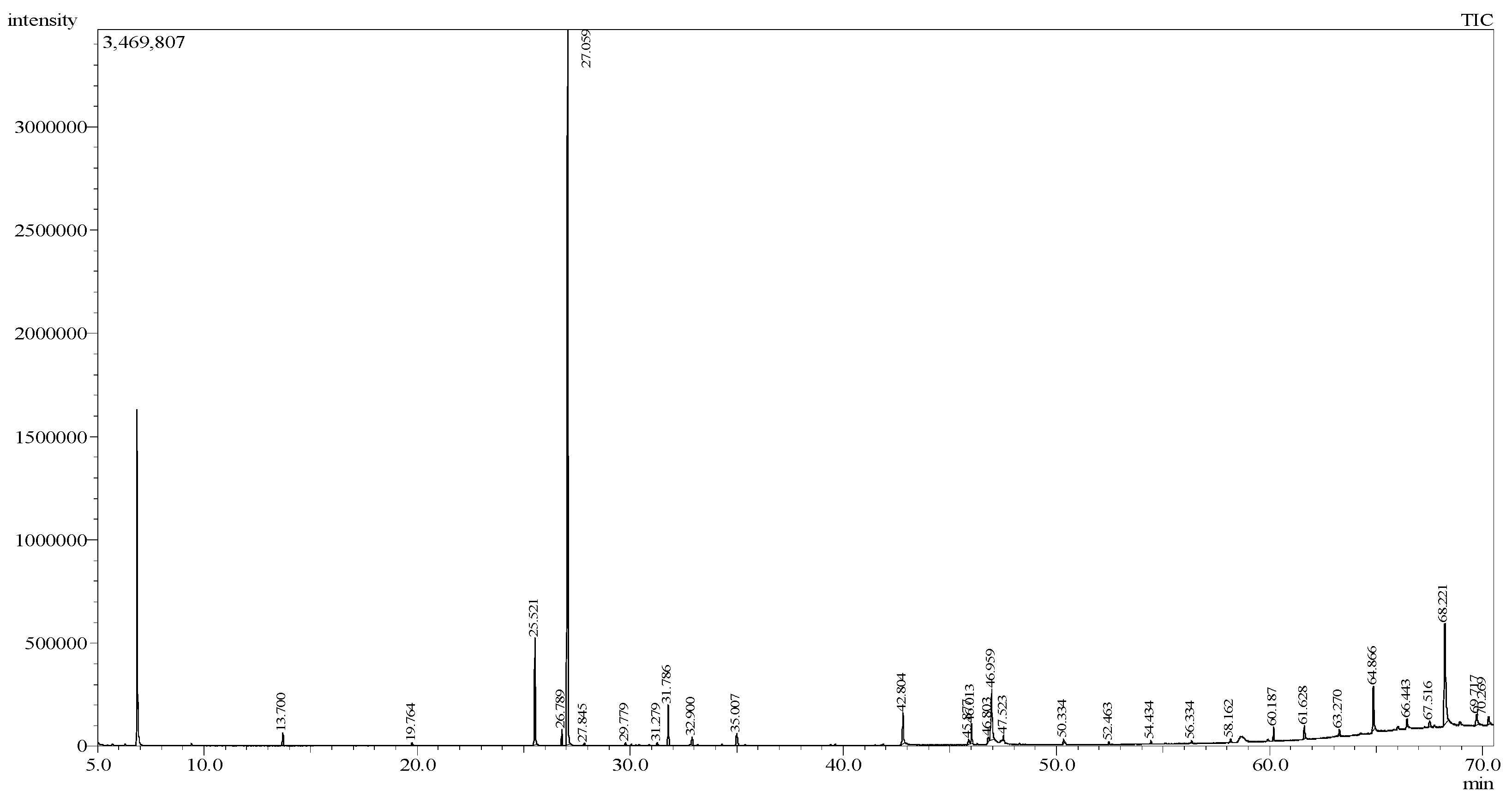
| Peak | Retention Time (min) | Peak Area (%) | Peak Height (%) | Compound Name | Molecular Weight | Chemical Formula | Matching Quality (%) |
|---|---|---|---|---|---|---|---|
| 1 | 13.700 | 0.79 | 1.00 | D-Limonene | 136 | C10H16 | 93 |
| 2 | 19.764 | 0.18 | 0.22 | Diketone alcohol | 116 | C6H12O2 | 89 |
| 3 | 25.521 | 7.77 | 8.43 | Eugenol | 164 | C10H12O2 | 95 |
| 4 | 26.789 | 1.04 | 1.25 | β-Elemene | 204 | C15H24 | 93 |
| 5 | 27.059 | 55.28 | 56.44 | Methyl eugenol | 178 | C11H14O2 | 94 |
| 6 | 27.845 | 0.14 | 0.19 | - | - | - | - |
| 7 | 29.779 | 0.17 | 0.21 | - | - | - | - |
| 8 | 31.279 | 0.20 | 0.25 | - | - | - | - |
| 9 | 31.786 | 2.83 | 3.18 | Elemol | 222 | C15H26O | 93 |
| 10 | 32.900 | 0.59 | 0.65 | - | - | - | - |
| 11 | 35.007 | 1.07 | 0.90 | - | - | - | - |
| 12 | 42.804 | 2.61 | 2.43 | Palmitic acid | 256 | C16H32O2 | 94 |
| 13 | 45.877 | 0.40 | 0.40 | - | - | - | - |
| 14 | 46.013 | 1.32 | 1.58 | Methyl linolenate | 292 | C19H32O2 | 93 |
| 15 | 46.803 | 0.42 | 0.40 | cis, cis-Linoleic acid | 280 | C18H32O2 | 88 |
| 16 | 46.959 | 4.62 | 3.99 | Linolenic acid | 278 | C18H30O2 | 95 |
| 17 | 47.523 | 0.29 | 0.45 | Ethyl linolenate | 306 | C20H34O2 | 86 |
| 18 | 50.334 | 0.25 | 0.36 | - | - | - | - |
| 19 | 52.463 | 0.14 | 0.20 | - | - | - | - |
| 20 | 54.434 | 0.21 | 0.26 | - | - | - | - |
| 21 | 56.334 | 0.15 | 0.21 | - | - | - | - |
| 22 | 58.162 | 0.23 | 0.28 | - | - | - | - |
| 23 | 60.187 | 0.90 | 1.08 | Squalene | 410 | C30H50 | 89 |
| 24 | 61.628 | 0.89 | 1.03 | - | - | - | - |
| 25 | 63.270 | 0.40 | 0.48 | - | - | - | - |
| 26 | 64.866 | 3.25 | 3.49 | Hexatriacontane | 450 | C36H74 | 94 |
| 27 | 66.443 | 0.62 | 0.68 | - | - | - | - |
| 28 | 67.516 | 0.46 | 0.47 | - | - | - | - |
| 29 | 68.221 | 10.77 | 8.01 | Tetrapentacontane | 758 | C54H110 | 94 |
| 30 | 69.717 | 1.10 | 0.82 | - | - | - | - |
| 31 | 70.269 | 0.91 | 0.66 | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, H.; Lee, S.Y.; Ho, L.Y.; Sit, N.W. Mosquito Larvicidal Activity and Cytotoxicity of the Extracts of Aromatic Plants from Malaysia. Insects 2023, 14, 512. https://doi.org/10.3390/insects14060512
Lim H, Lee SY, Ho LY, Sit NW. Mosquito Larvicidal Activity and Cytotoxicity of the Extracts of Aromatic Plants from Malaysia. Insects. 2023; 14(6):512. https://doi.org/10.3390/insects14060512
Chicago/Turabian StyleLim, Huimei, Sook Yee Lee, Lai Yee Ho, and Nam Weng Sit. 2023. "Mosquito Larvicidal Activity and Cytotoxicity of the Extracts of Aromatic Plants from Malaysia" Insects 14, no. 6: 512. https://doi.org/10.3390/insects14060512