
Effects of Irradiation on Biology and Mating Behaviour of Wild Males of Brown Marmorated Stink Bug Using a 6 MV Medical Linear Accelerator


 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Rearing
2.2. Irradiation
2.3. Male Dose Response, Year 1
- (a)
- 0 Gy male × 0 Gy female (zero-dose control);
- (b)
- 16 Gy male × 0 Gy female;
- (c)
- 24 Gy male × 0 Gy female.
- (a)
- 0 Gy male × 0 Gy female (zero-dose control);
- (b)
- 16 Gy male × 0 Gy female;
- (c)
- 24 Gy male × 0 Gy female.
2.4. Male Dose Response, Year 2
- (a)
- 0 Gy male × 0 Gy female (zero-dose Cell control);
- (b)
- 0 Gy male × 0 Gy female (zero-dose control);
- (c)
- 16 Gy male × 0 Gy female;
- (d)
- 24 Gy male × 0 Gy female;
- (e)
- 32 Gy male × 0 Gy female;
- (f)
- 40 Gy male × 0 Gy female.
2.5. Cumulative Mortality in F1
2.6. Mating-Behaviour Bioassays
- (a)
- 0 Gy male × 0 Gy female (zero-dose control);
- (b)
- 32 Gy male × 0 Gy female;
- (c)
- 40 Gy male × 0 Gy female;
2.7. Statistical Methods
- (a)
- The mated status was ascertained by comparing the spermathecal widths of overwintering versus mated one-week-old females. A one-tailed Welch t-test [27], which considered the two means equal, was performed to test the null hypothesis.
- (b)
- The fecundity of the females was compared using a two-sample Brown–Mood median test [28] followed by a contingency table (2 × 6) χ2. The number of egg masses produced per female and the number of eggs per egg mass were compared with the Kruskal–Wallis test.
- (c)
- The fertility of males and the longevities of both males and females for the irradiated (16, 24, 32 and 40 Gy) and two unirradiated control groups (0 Gy Cell and 0 Gy) were compared through a Kruskal–Wallis test. In cases of significance, a Mann–Whitney pairwise test with the Bonferroni correction was then performed as a post hoc test.
- (d)
- To evaluate possible cumulative mortality effects in the F1 generation among the two highest dosages of male irradiation (32 and 40 Gy) and a 0 Gy control, a χ2 test in a contingency table (2 × 3) was conducted for each stage (nymph from second to fifth instar and adult). Ryan’s test for multiple comparisons of proportions [29] was performed as a post hoc test.
- (e)
- The numbers of matings, the mating durations and the total mating times were compared across treatments with the Kruskal–Wallis test, which was followed by the Mann–Whitney pairwise test with the Bonferroni correction as a post hoc test in cases of significance.
3. Results
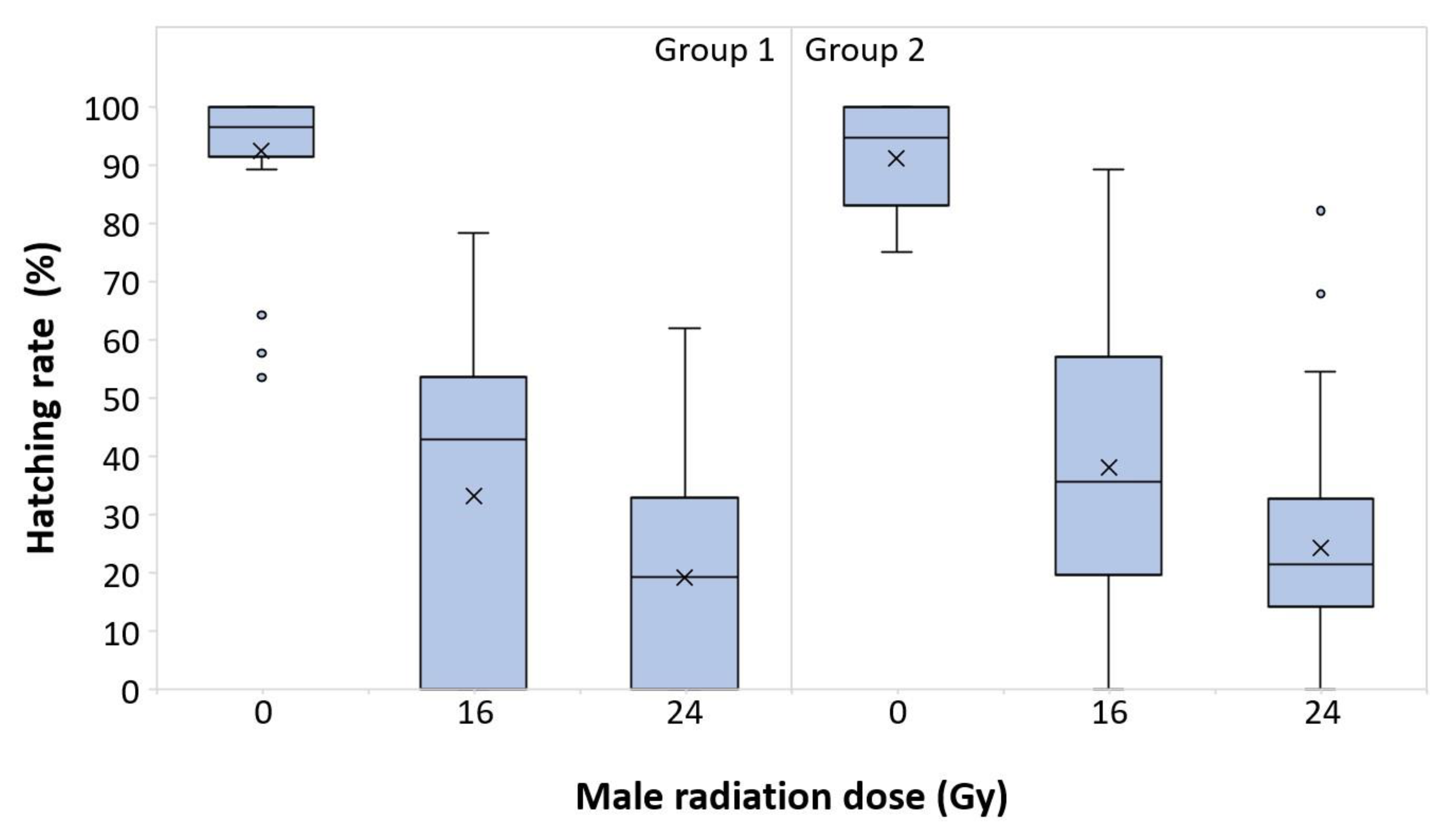
3.1. Male Dose Response, Year 1
3.2. Male Dose Response, Year 2
3.3. Cumulative Mortality in F1
3.4. Mating Behavioural Bioassays
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, D.H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the Biology, Ecology, and Management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Entomol. 2013, 42, 627–641. [Google Scholar] [CrossRef]
- Costi, E.; Haye, T.; Maistrello, L. Biological Parameters of the Invasive Brown Marmorated Stink Bug, Halyomorpha halys, in Southern Europe. J. Pest Sci. 2017, 90, 1059–1067. [Google Scholar] [CrossRef]
- Leskey, T.C.; Nielsen, A.L. Impact of the invasive brown marmorated stink bug in North America and Europe: History, biology, ecology, and management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kriticos, D.J.; Kean, J.M.; Phillips, C.B.; Senay, S.D.; Acosta, H.; Haye, T. The Potential Global Distribution of the Brown Marmorated Stink Bug, Halyomorpha halys, a Critical Threat to Plant Biosecurity. J. Pest Sci. 2017, 90, 1033–1043. [Google Scholar] [CrossRef]
- Inkley, D.B. Characteristics of Home Invasion by the Brown Marmorated Stink Bug (Hemiptera: Pentatomidae). J. Entomol. Sci. 2012, 47, 125–130. [Google Scholar] [CrossRef]
- Rice, K.B.; Bergh, C.J.; Bergmann, E.J.; Biddinger, D.J.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Haye, T.; et al. Biology, Ecology, and Management of Brown Marmorated Stink Bug (Hemiptera: Pentatomidae). J. Integr. Pest Manag. 2014, 5, A1–A13. [Google Scholar] [CrossRef]
- Malek, R.; Tattoni, C.; Ciolli, M.; Corradini, S.; Andreis, D.; Ibrahim, A.; Mazzoni, V.; Eriksson, A.; Anfora, G. Coupling Traditional Monitoring and Citizen Science to Disentangle the Invasion of Halyomorpha halys. ISPRS Int. J. Geo-Inf. 2018, 7, 171. [Google Scholar] [CrossRef] [Green Version]
- Kuhar, T.; Wallingford, A.; Philips, C.; Aigner, J.; Kamminga, K. E72 Evaluation of Foliar Insecticides for the Control of Brown Marmorated Stink Bugs in Tomatoes in Virginia, 2011. Arthropod Manag. Tests 2012, 37, 2–3. [Google Scholar] [CrossRef] [Green Version]
- Kuhar, T.P.; Kamminga, K. Review of the Chemical Control Research on Halyomorpha halys in the USA. J. Pest Sci. 2017, 90, 1021–1031. [Google Scholar] [CrossRef]
- Leskey, T.C.; Short, B.D.; Lee, D.H. Efficacy of Insecticide Residues on Adult Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) Mortality and Injury in Apple and Peach Orchards. Pest Manag. Sci. 2014, 70, 1097–1104. [Google Scholar] [CrossRef]
- Abram, P.K.; Hoelmer, K.A.; Acebes-Doria, A.; Andrews, H.; Beers, E.H.; Bergh, J.C.; Bessin, R.; Biddinger, D.; Botch, P.; Buffington, M.L.; et al. Indigenous Arthropod Natural Enemies of the Invasive Brown Marmorated Stink Bug in North America and Europe. J. Pest Sci. 2017, 90, 1009–1020. [Google Scholar] [CrossRef] [Green Version]
- Conti, E.; Avila, G.; Barratt, B.; Cingolani, F.; Colazza, S.; Guarino, S.; Hoelmer, K.; Laumann, R.A.; Maistrello, L.; Martel, G.; et al. Biological Control of Invasive Stink Bugs: Review of Global State and Future Prospects. Entomol. Exp. Appl. 2021, 169, 28–51. [Google Scholar] [CrossRef]
- Klassen, W.; Curtis, C.F. History of the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 3–44. Available online: https://www.taylorfrancis.com/chapters/oa-edit/10.1201/9781003035572-1/history-sterile-insect-technique-klassen-curtis-hendrichs (accessed on 20 February 2023). [CrossRef]
- Lance, D.R.; McInnis, D.O. Biological Basis of the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 113–142. [Google Scholar] [CrossRef]
- Enkerlin, W.R.; Gutiérrez Ruelas, J.M.; Pantaleon, R.; Soto Litera, C.; Villaseñor Cortés, A.; Zavala López, J.L.; Orozco Dávila, D.; Montoya Gerardo, P.; Silva Villarreal, L.; Cotoc Roldán, E.; et al. The Moscamed Regional Programme: Review of a Success Story of Area-Wide Sterile Insect Technique Application. Entomol. Exp. Appl. 2017, 164, 188–203. [Google Scholar] [CrossRef] [Green Version]
- Vreysen, M.J.B.; Saleh, K.M.; Ali, M.Y.; Abdulla, A.M.; Zhu, Z.-R.; Juma, K.G.; Dyck, V.A.; Msangi, A.R.; Mkonyi, P.A.; Udo Feldmann, A.H. Glossina austeni (Diptera: Glossinidae) Eradicated on the Island of Unguja, Zanzibar, Using the Sterile Insect Technique. J. Econ. Entomol. 2000, 93, 123–135. [Google Scholar] [CrossRef]
- Lees, R.S.; Caravalho, D.O.; Bouyer, J. Potential Impact of Integrating the Sterile Insect Technique into the Fight Against Disease-Transmitting Mosquitoes. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 1081–1118. Available online: https://www.taylorfrancis.com/chapters/oa-edit/10.1201/9781003035572-33/potential-impact-integrating-sterile-insect-technique-fight-disease-transmitting-mosquitoes-lees-carvalho-bouyer (accessed on 20 February 2023). [CrossRef]
- Welsh, T.J.; Stringer, L.D.; Caldwell, R.; Carpenter, J.E.; Suckling, D.M. Irradiation Biology of Male Brown Marmorated Stink Bugs: Is There Scope for the Sterile Insect Technique? Int. J. Radiat. Biol. 2017, 93, 1357–1363. [Google Scholar] [CrossRef] [PubMed]
- Suckling, D.M.; Cristofaro, M.; Roselli, G.; Levy, M.C.; Cemmi, A.; Mazzoni, V.; Stringer, L.D.; Zeni, V.; Ioriatti, C.; Anfora, G. The Competitive Mating of Irradiated Brown Marmorated Stink Bugs, Halyomorpha halys, for the Sterile Insect Technique. Insects 2019, 10, 411. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.N.; Stringer, L.D.; Hong, K.J. Influence of Irradiation on the Biology of the Brown Marmorated Stink Bug (Hemiptera: Pentatomidae). J. Econ. Entomol. 2021, 114, 1211–1218. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Fleischer, S.; Hamilton, G.C.; Hancock, T.; Krawczyk, G.; Lee, J.C.; Ogburn, E.; Pote, J.M.; Raudenbush, A.; Rucker, A.; et al. Phenology of Brown Marmorated Stink Bug Described Using Female Reproductive Development. Ecol. Evol. 2017, 7, 6680–6690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, C.M.; Coffey, P.L.; Hamby, K.A.; Dively, G.P. Laboratory Rearing of Halyomorpha halys: Methods to Optimize Survival and Fitness of Adults during and after Diapause. J. Pest Sci. 2017, 90, 1069–1077. [Google Scholar] [CrossRef]
- Koutsogeorgiou, E.I.; Kouloussis, N.A.; Koveos, D.S.; Andreadis, S.S. Females of Halyomorpha halys (Hemiptera: Pentatomidae) Experience a Facultative Reproductive Diapause in Northern Greece. Insects 2022, 13, 866. [Google Scholar] [CrossRef]
- Suckling, D.M.; Levy, M.C.; Roselli, G.; Mazzoni, V.; Ioriatti, C.; Deromedi, M.; Cristofaro, M.; Anfora, G. Live Traps for Adult Brown Marmorated Stink Bugs. Insects 2019, 10, 376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horrocks, K.J.; Welsh, T.; Carpenter, J.E.; Suckling, D.M. EGG Sterilisation of Irradiated Nezara viridula (Hemiptera: Pentatomidae). Insects 2020, 11, 564. [Google Scholar] [CrossRef] [PubMed]
- Cristofaro, M.; Sforza, R.F.H.; Roselli, G.; Paolini, A.; Cemmi, A.; Musmeci, S.; Anfora, G.; Mazzoni, V.; Grodowitz, M. Effects of Gamma Irradiation on the Fecundity, Fertility, and Longevity of the Invasive Stink Bug Pest Bagrada hilaris (Burmeister) (Hemiptera: Pentatomidae). Insects 2022, 13, 787. [Google Scholar] [CrossRef] [PubMed]
- Welch, B.L. The Generalization of ‘Student’s‘ Problem When Several Different Population Variances are Involved. Biometrika 1947, 34, 28–35. [Google Scholar] [CrossRef]
- Brown, G.W.; Mood, A.M. On median tests for linear hypotheses. In Proceedings of the Second Berkeley Symposium on Mathematical Statistics and Probability, Los Angeles, CA, USA, 31 July–12 August 1950; University of California Press: Berkeley, CA, USA, 1951; Volume 2, pp. 159–166. [Google Scholar]
- Ryan, T.H. Significance Tests for Multiple Comparison of Proportions, Variances, and Other Statistics. Psychol. Bull. 1960, 57, 318–328. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.r-project.org/index.html (accessed on 20 February 2023).
- De Boeck, P.; Bakker, M.; Zwitser, R.; Nivard, M.; Hofman, A.; Tuerlinckx, F.; Partchev, I. The Estimation of Item Response Models with the Lmer Function from the Lme4 Package in R. J. Stat. Softw. 2011, 39, 33. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. Available online: https://www.nhm.uio.no/english/research/resources/past/ (accessed on 28 February 2023).
- Vreysen, M.J.; Hendrichs, J.; Enkerlin, W.R. The sterile insect technique as a component of sustainable Area-Wide integrated pest management of selected horticultural insect pests. J. Fruit Ornam. Plant Res. 2006, 14, 107. [Google Scholar]
- Hendrichs, J.; Robinson, A.S.; Cayol, J.P.; Enkerlin, W. Medfly Areawide Sterile Insect Technique Programmes for Prevention, Suppression or Eradication: The Importance of Mating Behavior Studies. Fla. Entomol. 2002, 85, 1–13. [Google Scholar] [CrossRef]
- De Beer, C.J.; Moyaba, P.; Boikanyo, S.N.B.; Majatladi, D.; Venter, G.J.; Vreysen, M.J.B. Gamma Irradiation and Male Glossina austeni Mating Performance. Insects 2020, 11, 522. [Google Scholar] [CrossRef]
- Ilboudo, K.; Camara, K.; Salou, E.W.; Gimonneau, G. Quality Control and Mating Performance of Irradiated Glossina palpalis gambiensis Males. Insects 2022, 13, 476. [Google Scholar] [CrossRef]
- Berryman, A. Mathematical Description of the Sterile Male Principle. Can. Entomol. 1967, 99, 858–865. [Google Scholar] [CrossRef]
- Zouros, E. On the Role of Female Monogamy in the Sterile Male Technique of Insect Control. Annales de l’Institut Phytopathol. Benaki 1969, 9, 20–29. [Google Scholar]
- Kawada, H.; Kitamura, C. The reproductive behavior of the brown marmorated stink bug, Halyomorpha mista Uhler (Heteroptera: Pentatomidae) I. Observation of mating behavior and multiple copulation. Appl. Ent. Zool. 1983, 18, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Klassen, W.; Creech, J.F. Suppression of Pest Population with Sterile Male Insects; Miscellaneous Publication, USDA/ARS: Washington, DC, USA, 1971; p. 1182. [Google Scholar]
- Nielsen, A.L.; Hamilton, G.C.; Matadha, D. Developmental Rate Estimation and Life Table Analysis for Halyomorpha halys (Hemiptera: Pentatomidae). Environ. Entomol. 2008, 37, 348–355. [Google Scholar] [CrossRef]
- Haye, T.; Abdallah, S.; Gariepy, T.; Wyniger, D. Phenology, Life Table Analysis and Temperature Requirements of the Invasive Brown Marmorated stink bug, Halyomorpha halys, in Europe. J. Pest Sci. 2014, 87, 407–418. [Google Scholar] [CrossRef]
- Lara, J.R.; Kamiyama, M.T.; Hernandez, G.; Nay, J.; Hoddle, M.S. Assessment of Feeding Acceptance and Injury of Kerman Pistachios, Pistacia vera, by Brown Marmorated Stink Bug (Hemiptera: Pentatomidae). J. Insect Sci. 2017, 17, 106. [Google Scholar] [CrossRef]
- Knipling, E.F. The Basic Principles of Insect Population Suppression and Management. In Agriculture Handbook; USDA/SEA: Washington, DC, USA, 1979; p. 512. [Google Scholar]
- Liebhold, A.M.; Berec, L.; Brockerhoff, E.G.; Epanchin-Niell, R.S.; Hastings, A.; Herms, D.A.; Kean, J.M.; McCullough, D.G.; Suckling, D.M.; Tobin, P.C.; et al. Eradication of Invading Insect Populations: From Concepts to Applications. Annu. Rev. Entomol. 2016, 61, 335–352. [Google Scholar] [CrossRef] [Green Version]
- Kutsaev, S.V.; Agustsson, R.; Berry, R.; Boucher, S.; Smirnov, A.Y. Electron Accelerator for Replacement of Radioactive Sources in Insect Sterilization Facilities. Phys. At. Nucl. 2021, 84, 1743–1747. [Google Scholar] [CrossRef]
- Wang, L.M.; Li, N.; Ren, C.P.; Peng, Z.Y.; Lu, H.Z.; Li, D.; Deng, S.Q. Sterility of Aedes albopictus by X-ray Irradiation as an Alternative to γ-ray Irradiation for the Sterile Insect Technique. Pathogens 2023, 12, 102. [Google Scholar] [CrossRef]
- Balestrino, F.; Mathis, A.; Lang, S.; Veronesi, E. Sterilization of Hulecoeteomyia japonica japonica (= Aedes japonicus japonicus) (Theobald, 1901) by High-Energy Photon Irradiation: Implications for a Sterile Insect Technique Approach in Europe. Med. Vet. Entomol. 2016, 30, 278–285. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coefficient | Estimate | Std. Error | t-Value | Pr(>|t|) |
|---|---|---|---|---|
| Intercept | −0.07863 | 0.02680 | −2.934 | 0.00377 ** 1 |
| Group 2 | −0.01489 | 0.05438 | −0.274 | 0.78447 |
| Radiation, 16 Gy | −1.02482 | 0.12472 | −8.217 | 3.39 × 10−14 *** 1 |
| Radiation, 24 Gy | −1.57294 | 0.18732 | −8.397 | 1.12 × 10−14 *** 1 |
| Group 2; Radiation, 16 Gy | 0.15456 | 0.15784 | 0.979 | 0.32874 |
| Group 2; Radiation, 24 Gy | 0.25251 | 0.22973 | 1.099 | 0.27311 |
| Male Dose (Gy) | n | n1 | No. of Egg Masses | Total Eggs Collected | % Egg-Laying Females | Eggs/Egg Mass | Egg Masses/Egg-Laying Females | Eggs/Female |
|---|---|---|---|---|---|---|---|---|
| 0 Cell 2 | 40 | 35 | 78 | 2052 | 60 | 26.31 | 3.75 | 56 |
| 0 | 40 | 36 | 79 | 2018 | 80.56 | 25.54 | 2.72 | 50 |
| 16 | 40 | 35 | 75 | 1954 | 57.14 | 26.05 | 3.45 | 41.5 |
| 24 | 40 | 32 | 46 | 1185 | 43.75 | 25.76 | 3.29 | 27.5 |
| 32 | 40 | 35 | 98 | 2513 | 62.86 | 25.64 | 4.45 | 95.5 |
| 40 | 40 | 33 | 71 | 1827 | 48.48 | 25.73 | 4.60 | 96 |
| p | 0.9 1 | 0.09 1 | 0.16 1 |
| Male Dose (Gy) | n | No. of Matings | Mating Time (min) | Total Mating Time (min) |
|---|---|---|---|---|
| 0 | 12 | 6 | 12 a | 72 |
| 32 | 11 | 5 | 13 a | 64 |
| 40 | 12 | 5 | 13.5 b | 66 |
| p | 0.15 1 | 5.997 × 10−6 1 | 0.55 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roselli, G.; Anfora, G.; Suckling, D.M.; Mazzoni, V.; Vanoni, V.; Menegotti, L.; Fellin, L.; Rossi Stacconi, M.V.; Ioriatti, C.; Cristofaro, M. Effects of Irradiation on Biology and Mating Behaviour of Wild Males of Brown Marmorated Stink Bug Using a 6 MV Medical Linear Accelerator. Insects 2023, 14, 460. https://doi.org/10.3390/insects14050460
Roselli G, Anfora G, Suckling DM, Mazzoni V, Vanoni V, Menegotti L, Fellin L, Rossi Stacconi MV, Ioriatti C, Cristofaro M. Effects of Irradiation on Biology and Mating Behaviour of Wild Males of Brown Marmorated Stink Bug Using a 6 MV Medical Linear Accelerator. Insects. 2023; 14(5):460. https://doi.org/10.3390/insects14050460
Chicago/Turabian StyleRoselli, Gerardo, Gianfranco Anfora, David Maxwell Suckling, Valerio Mazzoni, Valentina Vanoni, Loris Menegotti, Lorenzo Fellin, Marco Valerio Rossi Stacconi, Claudio Ioriatti, and Massimo Cristofaro. 2023. "Effects of Irradiation on Biology and Mating Behaviour of Wild Males of Brown Marmorated Stink Bug Using a 6 MV Medical Linear Accelerator" Insects 14, no. 5: 460. https://doi.org/10.3390/insects14050460