
Up and down from North to South: Latitudinal Distribution of Flea Beetle Genera in Continental Africa (Coleoptera, Chrysomelidae, Galerucinae, Alticini)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
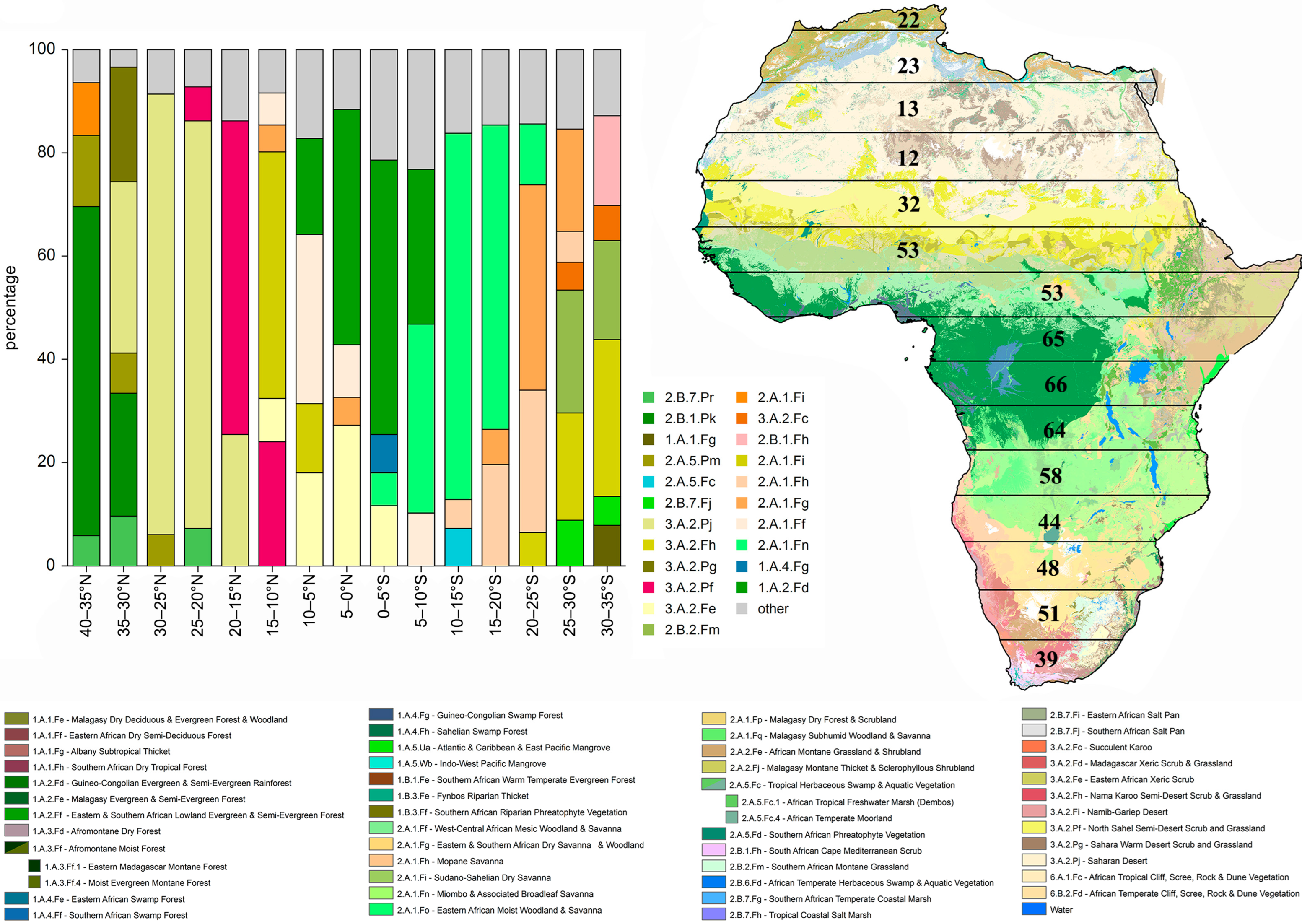
2.1. Study Area, Dataset, Vegetation Formations, and Bioclimatic Variables
2.2. Statistical and Spatial Analyses
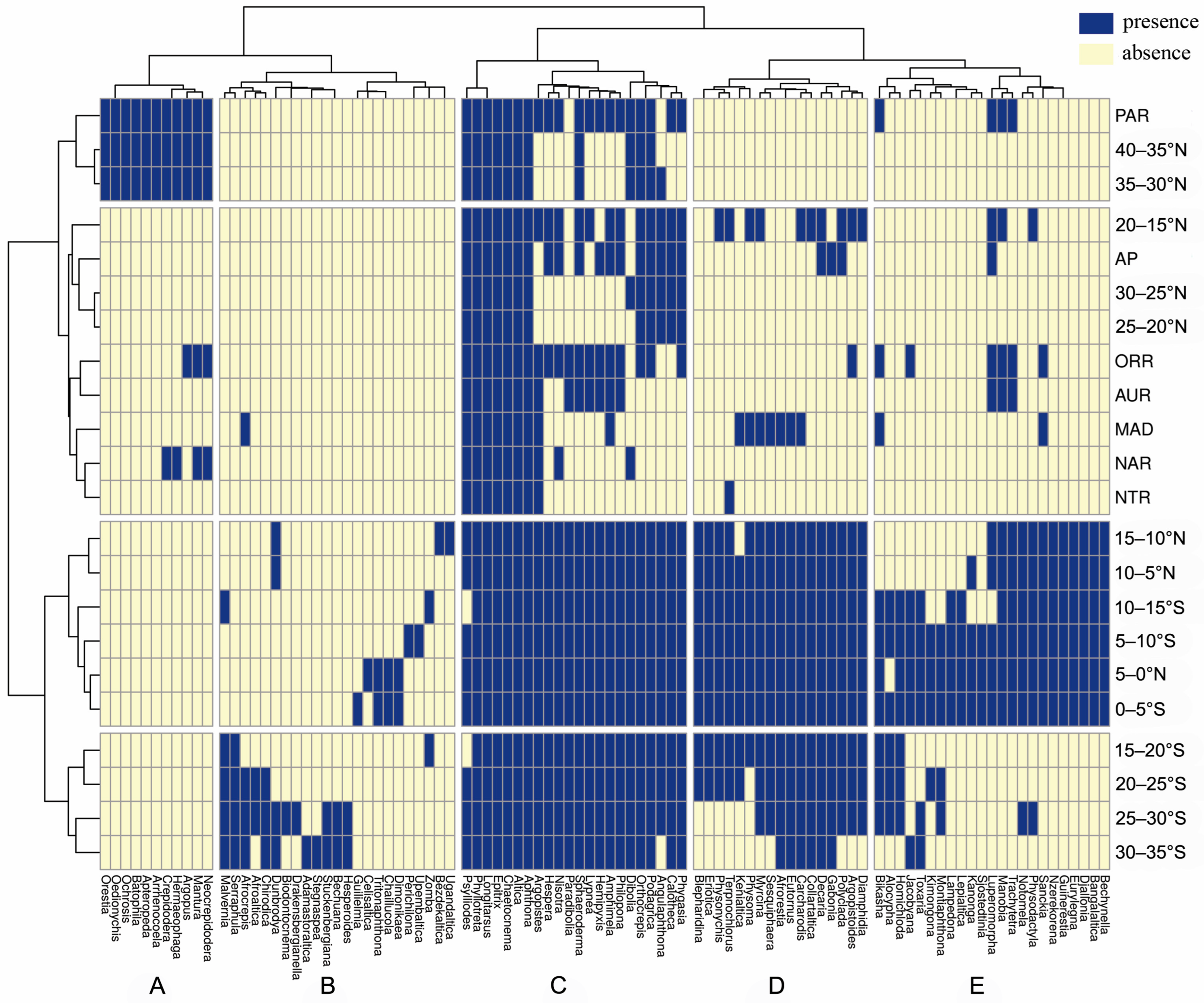
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaston, K.J. Global Patterns in Biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Pollock, L.J.; O’Connor, L.M.J.; Mokany, K.; Rosauer, D.F.; Talluto, M.V.; Thuiller, W. Protecting Biodiversity (in All Its Complexity): New Models and Methods. Trends Ecol. Evol. 2020, 35, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Biondi, M.; D’Alessandro, P.; De Simone, W.; Iannella, M. DBSCAN and GIE, Two Density-Based “Grid-Free” Methods for Finding Areas of Endemism: A Case Study of Flea Beetles (Coleoptera, Chrysomelidae) in the Afrotropical Region. Insects 2021, 12, 1115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, Y.-G.; Zhang, C.-Y.; Wang, T.-R.; Su, T.-H.; Huang, P.-H.; Meng, H.-H.; Li, J. Latitudinal Diversity Gradient in the Changing World: Retrospectives and Perspectives. Diversity 2022, 14, 334. [Google Scholar] [CrossRef]
- Jetz, W.; Thomas, G.H.; Joy, J.B.; Hartmann, K.; Mooers, A.O. The Global Diversity of Birds in Space and Time. Nature 2012, 491, 444–448. [Google Scholar] [CrossRef]
- Pyron, R.A.; Wiens, J.J. Large-Scale Phylogenetic Analyses Reveal the Causes of High Tropical Amphibian Diversity. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131622. [Google Scholar] [CrossRef]
- Kerkhoff, A.J.; Moriarty, P.E.; Weiser, M.D. The Latitudinal Species Richness Gradient in New World Woody Angiosperms Is Consistent with the Tropical Conservatism Hypothesis. Proc. Natl. Acad. Sci. USA 2014, 111, 8125–8130. [Google Scholar] [CrossRef]
- Rolland, J.; Condamine, F.L.; Jiguet, F.; Morlon, H. Faster Speciation and Reduced Extinction in the Tropics Contribute to the Mammalian Latitudinal Diversity Gradient. PLoS Biol. 2014, 12, e1001775. [Google Scholar] [CrossRef]
- Scherson, R.A.; Thornhill, A.H.; Urbina-Casanova, R.; Freyman, W.A.; Pliscoff, P.A.; Mishler, B.D. Spatial Phylogenetics of the Vascular Flora of Chile. Mol. Phylogenet. Evol. 2017, 112, 88–95. [Google Scholar] [CrossRef]
- Dagallier, L.M.J.; Janssens, S.B.; Dauby, G.; Blach-Overgaard, A.; Mackinder, B.A.; Droissart, V.; Svenning, J.; Sosef, M.S.M.; Stévart, T.; Harris, D.J.; et al. Cradles and Museums of Generic Plant Diversity across Tropical Africa. New Phytol. 2020, 225, 2196–2213. [Google Scholar] [CrossRef]
- McKenna, D.D.; Farrell, B.D. Tropical Forests Are Both Evolutionary Cradles and Museums of Leaf Beetle Diversity. Proc. Natl. Acad. Sci. USA 2006, 103, 10947–10951. [Google Scholar] [CrossRef]
- Condamine, F.L.; Sperling, F.A.H.; Wahlberg, N.; Rasplus, J.-Y.; Kergoat, G.J. What Causes Latitudinal Gradients in Species Diversity? Evolutionary Processes and Ecological Constraints on Swallowtail Biodiversity: Phylogeny and Latitudinal Diversity Gradient. Ecol. Lett. 2012, 15, 267–277. [Google Scholar] [CrossRef]
- Jablonski, D.; Huang, S.; Roy, K.; Valentine, J.W. Shaping the Latitudinal Diversity Gradient: New Perspectives from a Synthesis of Paleobiology and Biogeography. Am. Nat. 2017, 189, 1–12. [Google Scholar] [CrossRef]
- Owens, H.L.; Lewis, D.S.; Dupuis, J.R.; Clamens, A.; Sperling, F.A.H.; Kawahara, A.Y.; Guralnick, R.P.; Condamine, F.L.; Kerr, J. The Latitudinal Diversity Gradient in New World Swallowtail Butterflies Is Caused by Contrasting Patterns of Out-of- and Into-the-tropics Dispersal. Glob. Ecol. Biogeogr. 2017, 26, 1447–1458. [Google Scholar] [CrossRef]
- D’Alessandro, P.; Iannella, M.; Frasca, R.; Biondi, M. Distribution Patterns and Habitat Preference for the Genera-Group Blepharida s.l. in Sub-Saharan Africa (Coleoptera: Chrysomelidae: Galerucinae: Alticini). Zool. Anz. 2018, 277, 23–32. [Google Scholar] [CrossRef]
- Economo, E.P.; Narula, N.; Friedman, N.R.; Weiser, M.D.; Guénard, B. Macroecology and Macroevolution of the Latitudinal Diversity Gradient in Ants. Nat. Commun. 2018, 9, 1778. [Google Scholar] [CrossRef]
- Iannella, M.; D’Alessandro, P.; De Simone, W.; Biondi, M. Habitat Specificity, Host Plants and Areas of Endemism for the Genera-Group Blepharida s.l. in the Afrotropical Region (Coleoptera, Chrysomelidae, Galerucinae, Alticini). Insects 2021, 12, 299. [Google Scholar] [CrossRef]
- Nadein, K.S.; Bezdêk, J. Galerucinae Latreille 1802. In Coleoptera, Beetles. Vol. 3. Morphology and Systematics (Phytophaga); Leschen, R.A.B., Beutel, R.G., Eds.; De Gruyter: Berlin, Germany, 2014; pp. 251–259. [Google Scholar]
- Bieńkowski, A.O.; Orlova-Bienkowskaja, M.J. Alien Leaf Beetles (Coleoptera, Chrysomelidae) of European Russia and Some General Tendencies of Leaf Beetle Invasions. PLoS ONE 2018, 13, e0203561. [Google Scholar] [CrossRef]
- Iannella, M.; D’Alessandro, P.; Longo, S.; Biondi, M. New Records and Potential Distribution by Ecological Niche Modeling of Monoxia obesula in the Mediterranean Area. Bull. Insectology 2019, 72, 135–142. [Google Scholar]
- Iannella, M.; D’Alessandro, P.; Biondi, M. Forecasting the Spread Associated with Climate Change in Eastern Europe of the Invasive Asiatic Flea Beetle, Luperomorpha xanthodera (Coleoptera: Chrysomelidae). Eur. J. Entomol. 2020, 117, 130–138. [Google Scholar] [CrossRef]
- Iannella, M.; De Simone, W.; D’Alessandro, P.; Biondi, M. Climate Change Favours Connectivity between Virus-Bearing Pest and Rice Cultivations in Sub-Saharan Africa, Depressing Local Economies. PeerJ 2021, 9, e12387. [Google Scholar] [CrossRef] [PubMed]
- De Simone, W.; Iannella, M.; D’Alessandro, P.; Biondi, M. Assessing Influence in Biofuel Production and Ecosystem Services When Environmental Changes Affect Plant–Pest Relationships. GCB Bioenergy 2020, 12, 864–877. [Google Scholar] [CrossRef]
- Douglas, H.B.; Konstantinov, A.S.; Brunke, A.J.; Moseyko, A.G.; Chapados, J.T.; Eyres, J.; Richter, R.; Savard, K.; Sears, E.; Prathapan, K.D.; et al. Phylogeny of the Flea Beetles (Galerucinae: Alticini) and the Position of Aulacothorax Elucidated through Anchored Phylogenomics (Coleoptera: Chrysomelidae: Alticini). Syst. Entomol. 2023, 1–23. [Google Scholar] [CrossRef]
- Jolivet, P.; Verma, K.K. Biology of Leaf Beetles; Intercept: Hampshire, UK, 2002; ISBN 978-1-898298-86-1. [Google Scholar]
- Biondi, M.; Urbani, F.; D’Alessandro, P. Relationships between the Geographic Distribution of Phytophagous Insects and Different Types of Vegetation: A Case Study of the Flea Beetle Genus Chaetocnema (Coleoptera: Chrysomelidae) in the Afrotropical Region. Eur. J. Entomol. 2015, 112, 311–327. [Google Scholar] [CrossRef]
- McGeoch, M.A. The Selection, Testing and Application of Terrestrial Insects as Bioindicators. Biol. Rev. Camb. Philos. Soc. 1998, 73, 181–201. [Google Scholar] [CrossRef]
- Biondi, M.; D’Alessandro, P. The Genus Chabria Jacoby: First Records in the Afrotropical Region with Description of Three New Species from Madagascar and Annotated Worldwide Species Catalogue (Coleoptera, Chrysomelidae, Galerucinae, Alticini). Zool. Anz.-J. Comp. Zool. 2013, 252, 88–100. [Google Scholar] [CrossRef]
- Biondi, M.; D’Alessandro, P. Ntaolaltica and Pseudophygasia, Two New Flea Beetle Genera from Madagascar (Coleoptera: Chrysomelidae: Galerucinae: Alticini). Insect Syst. Evol. 2013, 44, 93–106. [Google Scholar] [CrossRef]
- D’Alessandro, P.; Urbani, F.; Biondi, M. Biodiversity and Biogeography in Madagascar: Revision of the Endemic Flea Beetle Genus Neodera Duvivier, 1891 with Description of 19 New Species (Coleoptera, Chrysomelidae, Galerucinae, Alticini). Syst. Entomol. 2014, 39, 710–748. [Google Scholar] [CrossRef]
- Biondi, M.; D’Alessandro, P. Revision of Diphaulacosoma Jacoby, an Endemic Flea Beetle Genus from Madagascar, with Description of Three New Species (Coleoptera: Chrysomelidae, Galerucinae, Alticini). Fragm. Entomol. 2016, 48, 143. [Google Scholar] [CrossRef]
- Iannella, M.; D’Alessandro, P.; Biondi, M. Entomological Knowledge in Madagascar by GBIF Datasets: Estimates on the Coverage and Possible Biases (Insecta). Fragm. Entomol. 2019, 51, 1–10. [Google Scholar] [CrossRef]
- ESRI, Inc. ArcGIS Pro 3.1 2023, Environmental Systems Research Institute: Redlands, CA, USA.
- Biondi, M. Hesperoides, a New “Hairy” Flea Beetle Genus from Southern Africa (Coleoptera: Chrysomelidae, Galerucinae, Alticini). Fragm. Entomol. 2017, 49, 151–158. [Google Scholar] [CrossRef]
- Biondi, M.; D’Alessandro, P. Genus-Group Names of Afrotropical Flea Beetles (Coleoptera: Chrysomelidae: Alticinae): Annotated Catalogue and Biogeographical Notes. Eur. J. Entomol. 2010, 107, 401–424. [Google Scholar] [CrossRef]
- Biondi, M.; D’Alessandro, P. Afrotropical Flea Beetle Genera: A Key to Their Identification, Updated Catalogue and Biogeographical Analysis (Coleoptera, Chrysomelidae, Galerucinae, Alticini). ZooKeys 2012, 253, 1–158. [Google Scholar] [CrossRef]
- Biondi, M.; D’Alessandro, P. Revision of the Afrotropical Genus Notomela Jacoby, 1899 with Description N. joliveti sp. n. from Principe Island (Coleoptera, Chrysomelidae, Galerucinae, Alticini). ZooKeys 2015, 547, 63–74. [Google Scholar] [CrossRef]
- Biondi, M.; D’Alessandro, P. Guilielmia Weise, a Little Known Afrotropical Flea Beetle Genus: Systematic Affinities and Description of a Second New Species from Central Africa (Coleoptera, Chrysomelidae, Galerucinae, Alticini). Zootaxa 2017, 4323, 572. [Google Scholar] [CrossRef]
- Biondi, M.; D’Alessandro, P. Taxonomic Revision of the Genus Angulaphthona (Coleoptera: Chrysomelidae: Galerucinae: Alticini). Eur. J. Entomol. 2018, 115, 30–44. [Google Scholar] [CrossRef]
- Biondi, M.; Frasca, R.; Grobbelaar, E.; D’Alessandro, P. Supraspecific Taxonomy of the Flea Beetle Genus Blepharida Chevrolat, 1836 (Coleoptera: Chrysomelidae) in the Afrotropical Region and Description of Afroblepharida Subgen. Nov. Insect Syst. Evol. 2017, 48, 97–155. [Google Scholar] [CrossRef]
- Biondi, M.; Iannella, M.; D’Alessandro, P. Adamastoraltica humicola, New Genus and New Species: The First Example of Possible Moss-Inhabiting Flea Beetle Genus from Sub-Saharan Africa (Coleoptera, Chrysomelidae, Galerucinae). Zootaxa 2020, 4763, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Biondi, M.; D’Alessandro, P.; Cerasoli, F.; De Simone, W.; Iannella, M. Taxonomy, Habitat Preference, and Niche Overlap of Two Arrow-Poison Flea Beetle Species of the Genus Polyclada in Sub-Saharan Africa (Coleoptera, Chrysomelidae). Insects 2022, 13, 668. [Google Scholar] [CrossRef]
- D’Alessandro, P.; Biondi, M. Ugandaltica Gen. n., a Tiny Flea Beetle from the Forest Canopy in Central Africa (Coleoptera, Chrysomelidae, Galerucinae, Alticini). ZooKeys 2018, 746, 123–136. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, P.; Frasca, R.; Grobbelaar, E.; Iannella, M.; Biondi, M. Systematics and Biogeography of the Afrotropical Flea Beetle Subgenus Blepharidina (Afroblepharida) Biondi & D’Alessandro, with Description of Seven New Species (Coleoptera, Chrysomelidae, Galerucinae, Alticini). Insect Syst. Evol. 2018, 49, 443–480. [Google Scholar] [CrossRef]
- D’Alessandro, P.; Iannella, M.; Biondi, M. Revision of the Afrotropical Flea Beetle Subgenus Blepharidina s. str. Bechyné (Coleoptera, Chrysomelidae). Zootaxa 2019, 4545, 32. [Google Scholar] [CrossRef]
- Döberl, M. Alticinae. In Catalogue of Palaearctic Coleoptera; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2010; Volume 6-Chrysomeloidea; pp. 491–563. ISBN 978-87-88757-71-2. [Google Scholar]
- Freeman, B.G.; Pennell, M.W. The Latitudinal Taxonomy Gradient. Trends Ecol. Evol. 2021, 36, 778–786. [Google Scholar] [CrossRef]
- Nie, R.-E.; Breeschoten, T.; Timmermans, M.J.T.N.; Nadein, K.; Xue, H.-J.; Bai, M.; Huang, Y.; Yang, X.-K.; Vogler, A.P. The Phylogeny of Galerucinae (Coleoptera: Chrysomelidae) and the Performance of Mitochondrial Genomes in Phylogenetic Inference Compared to Nuclear RRNA Genes. Cladistics 2018, 34, 113–130. [Google Scholar] [CrossRef]
- Sayre, R. A New Map of Standardized Terrestrial Ecosystems of Africa; Association of American Geographers: Washington, DC, USA, 2013; ISBN 978-0-89291-275-9. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- NCSS Statistical Software 2023. NCSS, LLC.: Kaysville, UT, USA. Available online: ncss.com/software/ncss (accessed on 1 March 2023).
- Metsalu, T.; Vilo, J. ClustVis: A Web Tool for Visualizing Clustering of Multivariate Data Using Principal Component Analysis and Heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Novotny, V.; Drozd, P.; Miller, S.E.; Kulfan, M.; Janda, M.; Basset, Y.; Weiblen, G.D. Why Are There So Many Species of Herbivorous Insects in Tropical Rainforests? Science 2006, 313, 1115–1118. [Google Scholar] [CrossRef]
- Norton, D.A.; Didham, R.K. Comment on “Why Are There So Many Species of Herbivorous Insects in Tropical Rainforests”? Science 2007, 315, 1666. [Google Scholar] [CrossRef]
- Freedman, A.H.; Harrigan, R.J.; Zhen, Y.; Hamilton, A.M.; Smith, T.B. Evidence for Ecotone Speciation across an African Rainforest-savanna Gradient. Mol. Ecol. 2023, 00, 1–14. [Google Scholar] [CrossRef]
- Silva-Pereira, I.; Meira-Neto, J.A.A.; Rezende, V.L.; Eisenlohr, P.V. Biogeographic Transitions as a Source of High Biological Diversity: Phylogenetic Lessons from a Comprehensive Ecotone of South America. Perspect. Plant Ecol. Evol. Syst. 2020, 44, 125528. [Google Scholar] [CrossRef]
- Kotze, D.J.; Samways, M.J. No General Edge Effects for Invertebrates at Afromontane Forest/Grassland Ecotones. Biodivers. Conserv. 2001, 10, 443–466. [Google Scholar] [CrossRef]
- Benton, M.J. The Red Queen and the Court Jester: Species Diversity and the Role of Biotic and Abiotic Factors Through Time. Science 2009, 323, 728–732. [Google Scholar] [CrossRef] [PubMed]
- Fraser, D.; Soul, L.C.; Tóth, A.B.; Balk, M.A.; Eronen, J.T.; Pineda-Munoz, S.; Shupinski, A.B.; Villaseñor, A.; Barr, W.A.; Behrensmeyer, A.K.; et al. Investigating Biotic Interactions in Deep Time. Trends Ecol. Evol. 2021, 36, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Winkler, I.S.; Mitter, C.; Scheffer, S.J. Repeated Climate-Linked Host Shifts Have Promoted Diversification in a Temperate Clade of Leaf-Mining Flies. Proc. Natl. Acad. Sci. USA 2009, 106, 18103–18108. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.C.; Crouch, N.M.A.; Ferguson, A.W.; Bates, J.M. Climatic Refugia and Reduced Extinction Correlate with Underdispersion in Mammals and Birds in Africa. Ecol. Evol. 2022, 12, e8752. [Google Scholar] [CrossRef]
- Li, J.; Li, Q.; Wu, Y.; Ye, L.; Liu, H.; Wei, J.; Huang, X. Mountains Act as Museums and Cradles for Hemipteran Insects in China: Evidence from Patterns of Richness and Phylogenetic Structure. Glob. Ecol. Biogeogr. 2021, 30, 1070–1085. [Google Scholar] [CrossRef]
- Sonne, J.; Dalsgaard, B.; Borregaard, M.K.; Kennedy, J.; Fjeldså, J.; Rahbek, C. Biodiversity Cradles and Museums Segregating within Hotspots of Endemism. Proc. R. Soc. B Biol. Sci. 2022, 289, 20221102. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genera | No. of Species in Continental Africa | Distribution |
|---|---|---|
| Adamastoraltica Biondi, Iannella and D’Alessandro, 2020 | 1 | SSA |
| Afroaltica Biondi and D’Alessandro, 2007 | 2 | SSA |
| Afrocrepis Bechyné, 1954 | 3 | SSA-MAD |
| Afrorestia Bechyné, 1959 | ≈20 | SSA-MAD |
| Alocypha Weise, 1911 | 1 | SSA |
| Altica Geoffroy, 1762 | >30 | World |
| Amphimela Chapuis, 1875 | >30 | SSA-AP-MAD-PAR-ORR-AUR |
| Angulaphthona Bechyné, 1960 | 7 | AFR-AP |
| Aphthona Chevrolat, 1836 | >30 | World |
| Apteropeda Motschulsky, 1860 | 1 | MAF-PAR |
| Argopistes Motschulsky, 1860 | ≈10 | MAF-PAR |
| Argopistoides Jacoby, 1892 | 4 | SSA-ORR |
| Argopus Fischer von Waldheim, 1824 | 1 | MAF-PAR-ORR |
| Arrhenocoela Foudras, 1861 | 1 | MAF-PAR |
| Bangalaltica Bechyné, 1960 | 1 | SSA |
| Batophila Foudras, 1860 | 1 | MAF-PAR |
| Bechuana Scherer, 1970 | 2 | SSA |
| Bechynella Biondi and D’Alessandro, 2010 | 3 | SSA |
| Bezdekaltica Döberl, 2012 | 1 | SSA |
| Bikasha Maulik, 1931 | ≈6 | SSA-MAD-PAR-ORR |
| Biodontocnema Biondi, 2000 | 1 | SSA |
| Blepharidina Bechyné, 1968 | >30 | SSA |
| Calotheca Heyden, 1887 | >30 | SSA-AP-PAR |
| Carcharodis Weise, 1910 | 7 | SSA-MAD |
| Celisaltica Biondi, 2001 | 1 | SSA |
| Chaetocnema Stephens, 1831 | >30 | World |
| Chaillucola Bechyné, 1968 | 1 | SSA |
| Chirodica Germar, 1834 | 8 | SSA |
| Collartaltica Bechyné, 1959 | 6 | SSA |
| Crepidodera Chevrolat, 1836 | 3 | MAF-PAR-NAR |
| Decaria Weise, 1895 | ≈20 | SSA-AP |
| Diamphidia Gerstaecker, 1855 | 17 | SSA |
| Dibolia Latreille, 1829 | ≈20 | AFR-PAR-NAR |
| Dimonikaea Bechyné, 1968 | 1 | SSA |
| Djallonia Bechyné, 1955 | 1 | SSA |
| Drakensbergianella Biondi and D’Alessandro, 2003 | 1 | SSA |
| Dunbrodya Jacoby, 1906 | 2 | SSA |
| Epitrix Foudras, 1860 | ≈12 | World |
| Eriotica Harold, 1877 | 7 | SSA |
| Eurylegna Weise, 1910 | 6 | SSA |
| Eutornus Clark, 1860 | ≈7 | SSA-MAD |
| Gabonia Jacoby, 1893 | >30 | SSA-AP |
| Guilielmia Weise, 1924 | 2 | SSA |
| Guinerestia Scherer, 1959 | 3 | SSA |
| Hemipyxis Chevrolat, 1836 | >30 | SSA-AP-PAR-ORR-AUR |
| Hermaeophaga Foudras, 1860 | 1 | MAF-PAR-NAR |
| Hespera Weise, 1889 | >30 | SSA-AP-PAR-ORR |
| Hesperoides Biondi, 2017 | 1 | SSA |
| Homichloda Weise, 1902 | 3 | SSA |
| Jacobyana Maulik, 1926 | 3 | SSA-ORR |
| Kanonga Bechyné, 1960 | 1 | SSA |
| Kenialtica Bechyné, 1960 | 7 | SSA-MAD |
| Kimongona Bechyné, 1959 | 3 | SSA |
| Lampedona Weise, 1907 | 3 | SSA |
| Lepialtica Scherer, 1962 | 4 | SSA |
| Longitarsus Berthold, 1827 | >30 | World |
| Luperomorpha Weise, 1887 | 2 | SSA-AP-PAR-ORR-AUR |
| Lypnea Baly, 1876 | ≈10 | SSA-PAR-ORR-AUR |
| Malvernia Jacoby, 1899 | 2 | SSA |
| Manobia Jacoby, 1885 | ≈15 | SSA-PAR-ORR-AUR |
| Mantura Stephens, 1831 | 4 | MAF-PAR-NAR-ORR |
| Montiaphthona Scherer, 1961 | 6 | SSA |
| Myrcina Chapuis, 1875 | ≈16 | SSA-MAD |
| Neocrepidodera Heikertinger, 1911 | 5 | MAF-PAR-NAR-ORR |
| Nisotra Baly, 1864 | >30 | SSA-AP-PAR-ORR-AUR |
| Notomela Jacoby, 1899 | 3 | SSA |
| Nzerekorena Bechyné, 1955 | 9 | SSA |
| Ochrosis Foudras, 1861 | 1 | MAF-PAR |
| Oedionychis Latreille, 1829 | 2 | MAF-PAR |
| Orestia Chevrolat, 1836 | 3 | MAF-PAR |
| Orthocrepis Weise, 1888 | >30 | AFR-AP-PAR-ORR |
| Paradibolia Baly, 1875 | 3 | SSA-ORR-AUR |
| Perichilona Weise, 1919 | 2 | SSA |
| Philopona Weise, 1903 | ≈20 | SSA-AP-PAR-ORR-AUR |
| Phygasia Chevrolat, 1836 | >30 | SSA-AP-PAR-ORR |
| Phyllotreta Chevrolat, 1836 | >30 | World |
| Physodactyla Chapuis, 1875 | 6 | SSA |
| Physoma Clark, 1863 | 2 | SSA-MAD |
| Physonychis Clark, 1860 | >30 | SSA |
| Podagrica Chevrolat, 1836 | >30 | SSA-AP-PAR-ORR |
| Polyclada Chevrolat, 1836 | 16 | SSA-AP |
| Psylliodes Berthold, 1827 | 9 | World |
| Sanckia Duvivier, 1891 | 4 | SSA-MAD-ORR |
| Serraphula Jacoby, 1897 | 19 | SSA |
| Sesquiphaera Bechyné, 1958 | ≈10 | SSA-MAD |
| Sjostedtinia Weise, 1910 | 2 | SSA |
| Sphaeroderma Stephens, 1831 | >30 | SSA-AP-PAR-ORR-AUR |
| Stegnaspea Baly, 1877 | 6 | SSA |
| Stuckenbergiana Scherer, 1963 | 1 | SSA |
| Terpnochlorus Fairmaire, 1904 | 2 | SSA-NTR |
| Toxaria Weise, 1903 | 5 | SSA |
| Trachytetra Sharp, 1886 | 5 | SSA-PAR-ORR-AUR |
| Tritonaphthona Bechyné, 1960 | 1 | SSA |
| Ugandaltica D’Alessandro and Biondi, 2018 | 1 | SSA |
| Upembaltica Bechyné, 1960 | 1 | SSA |
| Zomba Bryant, 1922 | 1 | SSA |
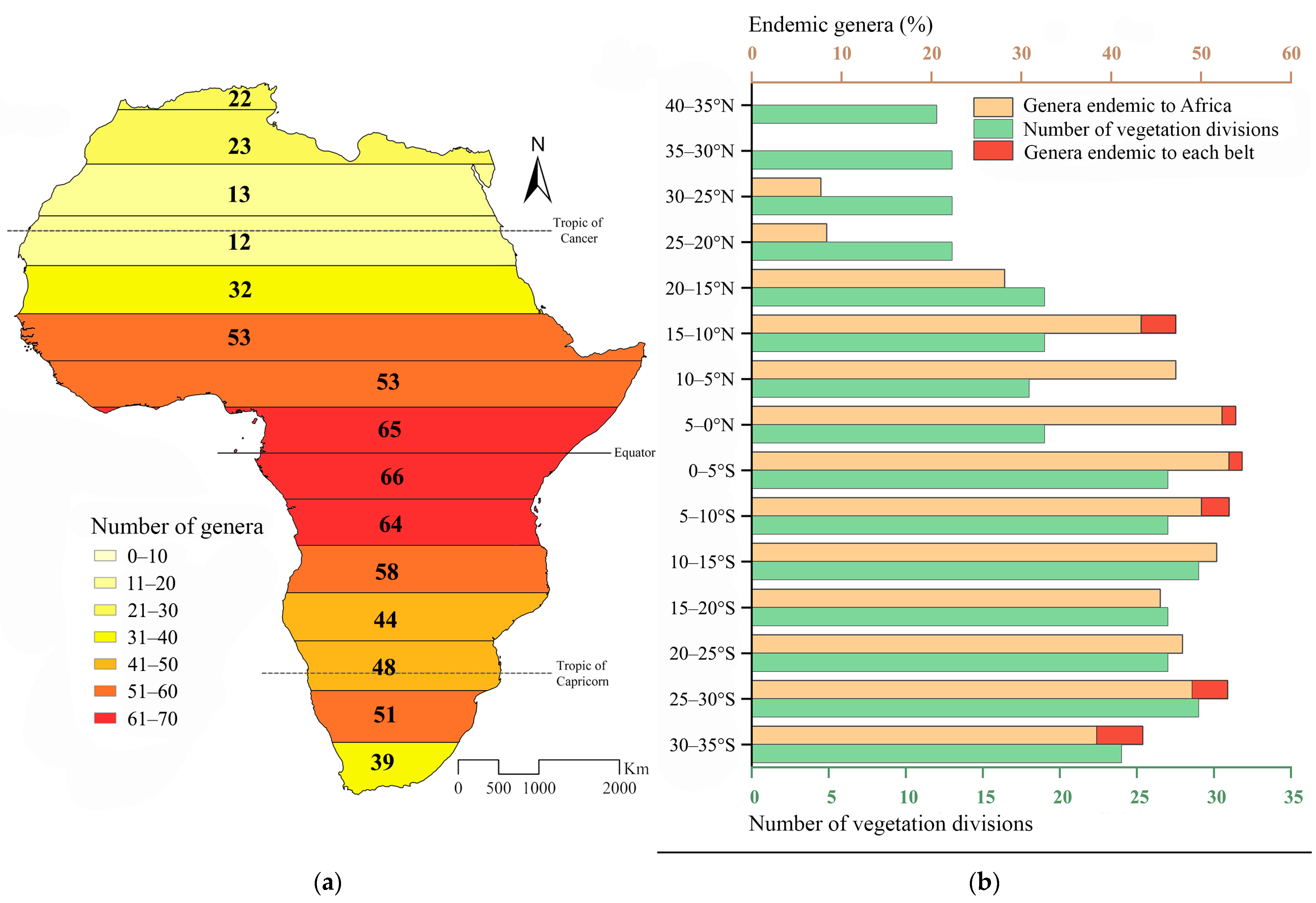
| Latitudinal Range | Total Area (km2) | Total Number of Flea Beetle Genera | African Endemic Genera (%) | Flea Beetle Genera Endemic to a Single Latitudinal Range (%) | Number of Vegetation Divisions |
|---|---|---|---|---|---|
| 40–35° N | 219,406 | 22 | 0 | 0 | 12 |
| 35–30° N | 1,441,210 | 23 | 0 | 0 | 13 |
| 30–25° N | 2,567,593 | 13 | 7.69 | 0 | 13 |
| 25–20° N | 2,990,429 | 12 | 8.33 | 0 | 13 |
| 20–15° | 3,228,301 | 32 | 28.13 | 0 | 19 |
| 15–10° N | 3,620,333 | 53 | 47.17 | 3.77 | 19 |
| 10–5° N | 3,673,971 | 53 | 47.17 | 0 | 18 |
| 5–0° N | 2,269,300 | 65 | 53.85 | 1.54 | 19 |
| 0–5° S | 1,888,226 | 66 | 54.55 | 1.51 | 27 |
| 5–10° S | 1,617,538 | 64 | 53.12 | 3.12 | 27 |
| 10–15° S | 1,648,111 | 58 | 51.72 | 0 | 29 |
| 15–20° S | 1,520,157 | 44 | 45.45 | 0 | 27 |
| 20–25° S | 1,201,049 | 48 | 47.92 | 0 | 27 |
| 25–30° S | 915,547 | 51 | 52.94 | 3.92 | 29 |
| 30–35° S | 478,961 | 39 | 43.49 | 5.13 | 24 |
| Latitudinal Range | BIO2 (Mean) | BIO3 (Mean) | BIO8 (Mean) | BIO9 (Mean) | BIO13 (Mean) | BIO14 (Mean) | BIO15 (Mean) | BIO18 (Mean) | BIO19 (Mean) |
|---|---|---|---|---|---|---|---|---|---|
| 40–35° N | 11.39 | 38.27 | 11.01 | 24.99 | 72.54 | 5.323 | 51.29 | 38.31 | 192.3 |
| 35–30° N | 12.94 | 40.44 | 13.78 | 28.04 | 25.4 | 1.265 | 60.87 | 9.296 | 60.28 |
| 30–25° N | 14.43 | 43.22 | 17.55 | 27.78 | 4.716 | 0.06 | 49.47 | 1.775 | 9.149 |
| 25–20° N | 14.9 | 46.42 | 29.45 | 21.57 | 6.837 | 0.122 | 67.47 | 10.19 | 1.752 |
| 20–15° | 15.11 | 52.89 | 31.09 | 22.97 | 44.36 | 0.137 | 139 | 29.21 | 1.894 |
| 15–10° N | 14.05 | 59.63 | 26.5 | 25.05 | 192.7 | 0.814 | 127.1 | 88.07 | 105.8 |
| 10–5° N | 11.94 | 68.53 | 24.58 | 25.03 | 223.7 | 6.895 | 84.81 | 195.3 | 422.1 |
| 5–0° N | 10.8 | 78.37 | 24.64 | 24.71 | 197.5 | 29.09 | 62.61 | 295.3 | 344.7 |
| 0–5° S | 9.799 | 78.05 | 23.63 | 22.66 | 215.4 | 27.19 | 58.34 | 403.6 | 155.5 |
| 5–10° S | 11.18 | 69.54 | 23.43 | 21.63 | 213 | 3.525 | 83.16 | 333.9 | 65.69 |
| 10–15° S | 12.74 | 61.17 | 22.47 | 18.63 | 237.1 | 0.716 | 104.3 | 288 | 7.666 |
| 15–20° S | 14.16 | 58.75 | 24.16 | 18.39 | 166.1 | 2.265 | 109.2 | 234.3 | 15.84 |
| 20–25° S | 15.27 | 59.7 | 24.83 | 16.41 | 91.84 | 2.446 | 95.73 | 215.6 | 11.29 |
| 25–30° S | 15.54 | 55.71 | 22.77 | 12.61 | 78.24 | 5.45 | 74.59 | 202.1 | 22.39 |
| 30–35° S | 14.41 | 54.25 | 18.41 | 13.32 | 61.6 | 13.83 | 46.66 | 134.9 | 70.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biondi, M.; D’Alessandro, P.; Iannella, M. Up and down from North to South: Latitudinal Distribution of Flea Beetle Genera in Continental Africa (Coleoptera, Chrysomelidae, Galerucinae, Alticini). Insects 2023, 14, 394. https://doi.org/10.3390/insects14040394
Biondi M, D’Alessandro P, Iannella M. Up and down from North to South: Latitudinal Distribution of Flea Beetle Genera in Continental Africa (Coleoptera, Chrysomelidae, Galerucinae, Alticini). Insects. 2023; 14(4):394. https://doi.org/10.3390/insects14040394
Chicago/Turabian StyleBiondi, Maurizio, Paola D’Alessandro, and Mattia Iannella. 2023. "Up and down from North to South: Latitudinal Distribution of Flea Beetle Genera in Continental Africa (Coleoptera, Chrysomelidae, Galerucinae, Alticini)" Insects 14, no. 4: 394. https://doi.org/10.3390/insects14040394