Wolbachia Effect on Drosophila melanogaster Lipid and Carbohydrate Metabolism

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Drosophila Lines
2.2. Body Mass and TAG Content Measurements
2.3. Colorimetric Method for Quantitative Determination of Total Lipid Content Using Sulfophosphovaniline (SPV) Reaction
2.4. Spectrophotometric Method of Evaluation of Carbohydrates Metabolism
2.5. Quantitative Real-Time PCR (qPCR)
2.6. Capillary Feeding Assay
2.7. Starvation Resistance Analysis
2.8. Statistical Analysis
3. Results
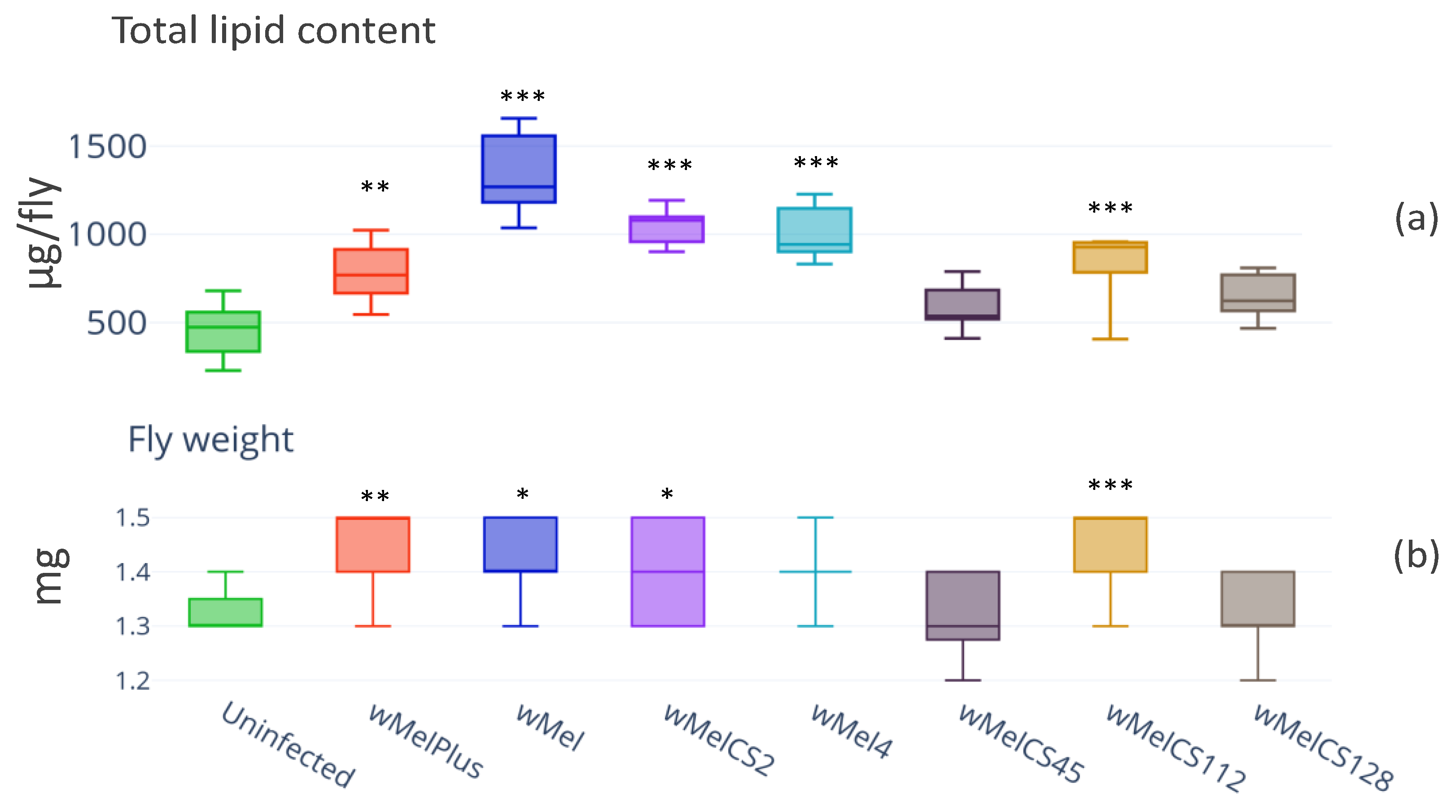
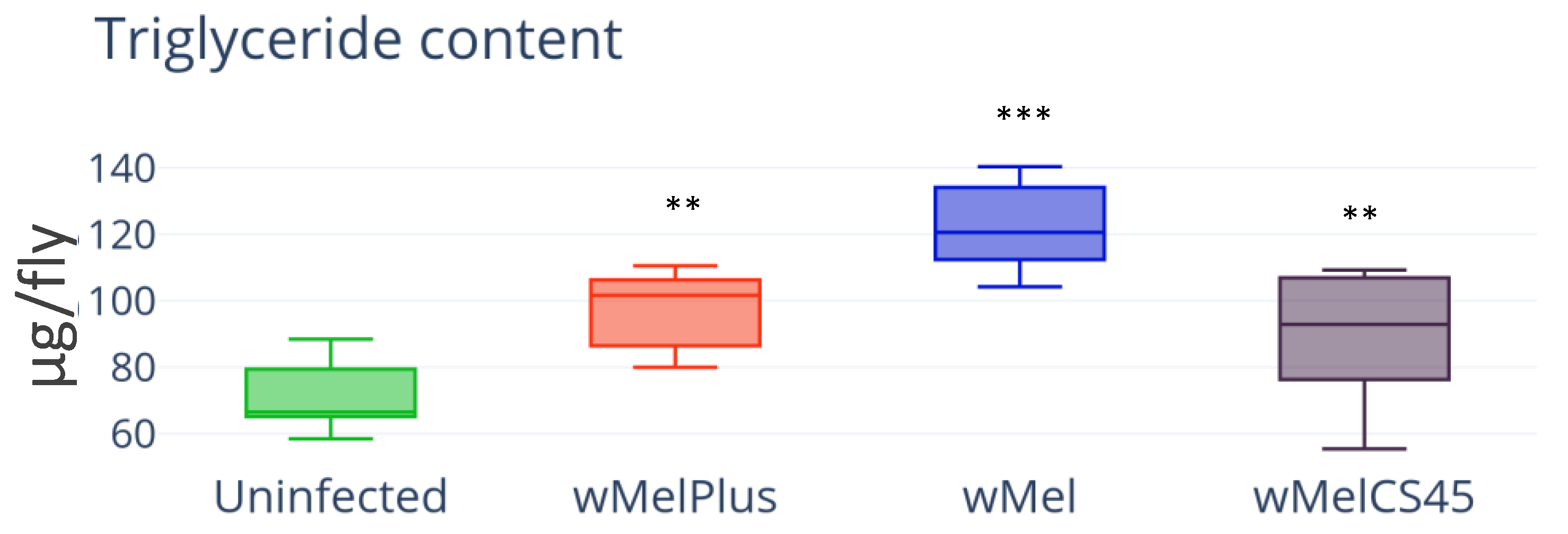
3.1. Lipid Content
3.2. Carbohydrate Content
3.3. Evaluation of the Effect the Wolbachia Infection Had on the Expression of Key Genes for Carbohydrate and Lipid Metabolism Using Quantitative Real-Time PCR
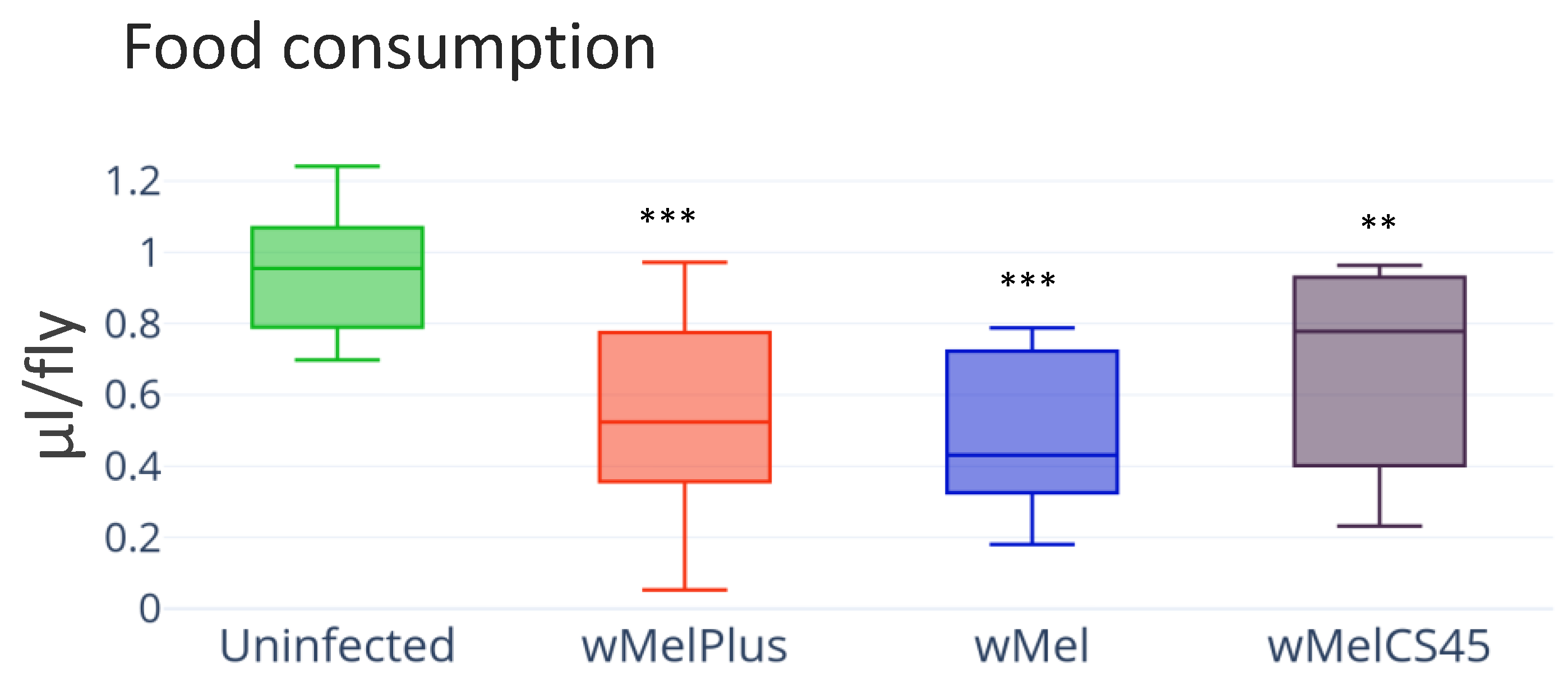
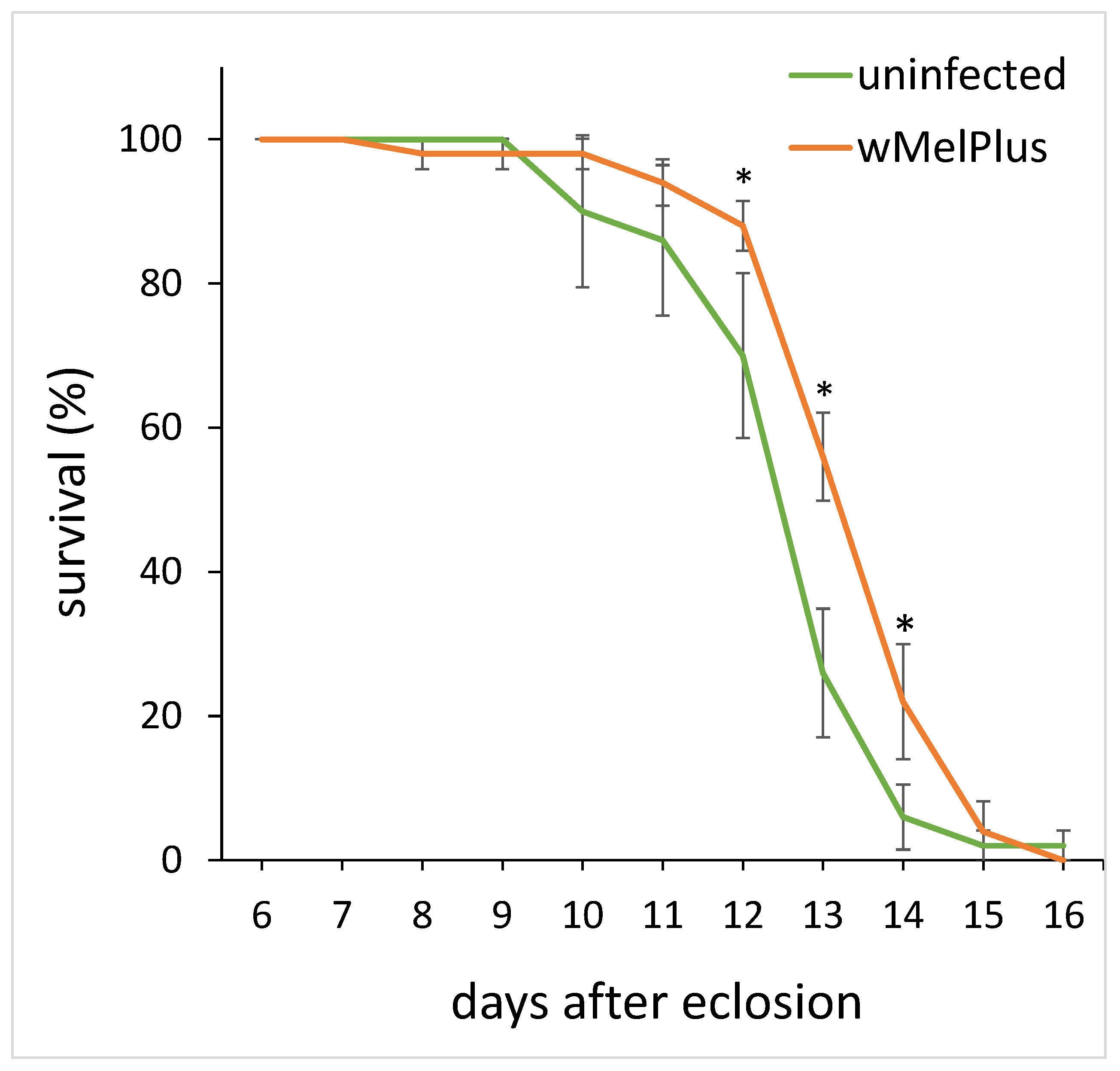
3.4. Feeding Behavior and Starvation Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Burdina, E.V.; Gruntenko, N.E. Physiological Aspects of Wolbachia pipientis–Drosophila melanogaster Relationship. J. Evol. Biochem. Phys. 2022, 58, 303–317. [Google Scholar] [CrossRef]
- Baldo, J.V.; Schwartz, S.; Wilkins, D.; Dronkers, N.F. Role of frontal versus temporal cortex in verbal fluency as revealed by voxel-based lesion-symptom mapping. J. Int. Neuropsychol. Soc. 2006, 12, 896–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, R.; Shropshire, J.D.; Cross, K.L.; Leigh, B.; Mansueto, A.J.; Stewart, V.; Bordenstein, S.R.; Bordenstein, S.R. Living in the endosymbiotic world of Wolbachia: A centennial review. Cell Host Microbe. 2021, 29, 879–893. [Google Scholar] [CrossRef]
- Ghedin, E.; Wang, S.; Spiro, D.; Caler, E.; Zhao, Q.; Crabtree, J.; Allen, J.E.; Delcher, A.L.; Guiliano, D.B.; Miranda-Saavedra, D.; et al. Draft genome of the filarial nematode parasite Brugia malayi. Science 2007, 317, 1756–1760. [Google Scholar] [CrossRef] [Green Version]
- Hosokawa, T.; Koga, R.; Kikuchi, Y.; Meng, X.-Y.; Fukatsu, T. Wolbachia as a bacteriocyte- associated nutritional mutualist. Proc. Natl. Acad. Sci. USA 2010, 107, 769–774. [Google Scholar] [CrossRef] [Green Version]
- Nikoh, N.; Hosokawa, T.; Moriyama, M.; Oshima, K.; Hattori, M.; Fukatsu, T. Evolutionary origin of insect-Wolbachia nutritional mutualism. Proc. Natl. Acad. Sci. USA 2014, 111, 10257–10262. [Google Scholar] [CrossRef] [Green Version]
- Dedeine, F.; Vavre, F.; Fleury, F.; Loppin, B.; Hochberg, M.E.; Bouletreau, M. Removing symbiotic Wolbachia bacteria specifically inhibits oogenesis in a parasitic wasp. Proc. Natl. Acas. Sci. USA 2001, 98, 6247–6252. [Google Scholar] [CrossRef] [Green Version]
- Landmann, F.; Voronin, D.; Sullivan, W.; Taylor, M.J. Anti-filarial activity of antibiotic therapy is due to extensive apoptosis after Wolbachia depletion from filarial nematodes. PLoS Pathog. 2011, 7, e1002351. [Google Scholar] [CrossRef] [Green Version]
- Starr, D.J.; Cline, T.W. A host parasite interaction rescues Drosophila oogenesis defects. Nature 2002, 418, 76–79. [Google Scholar] [CrossRef]
- Guo, Y.; Hoffmann, A.A.; Xu, X.Q.; Zhang, X.; Huang, H.J.; Ju, J.F.; Gong, J.T.; Hong, X.Y. Wolbachia-induced apoptosis associated with increased fecundity in Laodelphax striatellus (Hemiptera: Delphacidae). Insect Mol. Biol. 2018, 27, 796–807. [Google Scholar] [CrossRef]
- Chrostek, E.; Marialva, M.S.; Esteves, S.S.; Weinert, L.A.; Martinez, J.; Jiggins, F.M.; Teixeira, L. Wolbachia variants induce differential protection to viruses in Drosophila melanogaster: A phenotypic and phylogenomic analysis. PLoS Genet. 2013, 9, e1003896. [Google Scholar] [CrossRef] [Green Version]
- Hedges, L.M.; Brownlie, J.C.; O’Neill, S.L.; Johnson, K.N. Wolbachia and virus protection in insects. Science 2008, 322, 702. [Google Scholar] [CrossRef]
- Martinez, J.; Longdon, B.; Bauer, S.; Chan, Y.S.; Miller, W.J.; Bourtzis, K.; Teixeira, L.; Jiggins, F.M. Symbionts Commonly Provide Broad Spectrum Resistance to Viruses in Insects: A Comparative Analysis of Wolbachia Strains. PLoS Pathog. 2014, 10, e1004369. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, L.; Ferreira, Á.; Ashburner, M. The Bacterial Symbiont Wolbachia Induces Resistance to RNA Viral Infections in Drosophila melanogaster. PLoS Biol. 2008, 6, e1000002. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, A.C.; Cesar, C.S.; Martins, M.; Cogni, R. The Antiviral Effects of the Symbiont Bacteria Wolbachia in Insects. Front Immunol. 2021, 11, 626329. [Google Scholar] [CrossRef]
- Heier, C.; Kühnlein, R.P. Triacylglycerol Metabolism in Drosophila melanogaster. Genetics 2018, 210, 1163–1184. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.K.; Rulifson, E.J. Conserved mechanisms of glucose sensing and regulation by Drosophila corpora cardiaca cells. Nature 2004, 431, 316–320. [Google Scholar] [CrossRef]
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17–27. [Google Scholar] [CrossRef]
- Zheng, X.; Yang, Z.; Yue, Z.; Alvarez, J.D.; Sehgal, A. FOXO and insulin signaling regulate sensitivity of the circadian clock to oxidative stress. Proc. Natl. Acad. Sci. USA 2007, 104, 15899–15904. [Google Scholar] [CrossRef] [Green Version]
- Eremina, M.A.; Menshanov, P.N.; Shishkina, O.D.; Gruntenko, N.E. The transcription factor dfoxo controls the expression of insulin pathway genes and lipids content under heat stress in Drosophila melanogaster. Vavilovskii Zhurnal Genet Selektsii. 2021, 25, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Grönke, S.; Mildner, A.; Fellert, S.; Tennagels, N.; Petry, S.; Müller, G.; Jäckle, H.; Kühnlein, R.P. Brummer lipase is an evolutionary conserved fat storage regulator in Drosophila. Cell Metab. 2005, 1, 323–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnikow, E.; Xu, S.; Liu, J.; Bell, A.J.; Ghedin, E.; Unnasch, T.R.; Lustigman, S. A potential role for the interaction of Wolbachia surface proteins with the Brugia malayi glycolytic enzymes and cytoskeleton in maintenance of endosymbiosis. PLoS Negl. Trop. Dis. 2013, 7, e2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponton, F.; Wilson, K.; Holmes, A.; Raubenheimer, D.; Robinson, K.L.; Simpson, S.J. Macronutrients mediate the functional relationship between Drosophila and Wolbachia. Proc. Biol. Sci. 2015, 282, 20142029. [Google Scholar] [CrossRef] [Green Version]
- Gruntenko, N.E.; Ilinsky, Y.Y.; Adonyeva, N.V.; Burdina, E.V.; Bykov, R.A.; Menshanov, P.N.; Rauschenbach, I.Y. Various Wolbachia genotypes differently influence host Drosophila dopamine metabolism and survival under heat stress conditions. BMC Evol. Biol. 2017, 17, 252. [Google Scholar] [CrossRef] [Green Version]
- Burdina, E.V.; Bykov, R.A.; Menshanov, P.N.; Ilinsky, Y.Y.; Gruntenko, N.E. Unique Wolbachia strain wMelPlus increases heat stress resistance in Drosophila melanogaster. Arch. Insect Biochem. Physiol. 2021, 106, e21776. [Google Scholar] [CrossRef]
- Riegler, M.; Sidhu, M.; Miller, W.J.; O’Neill, S.L. Evidence for a global Wolbachia replacement in Drosophila melanogaster. Curr. Biol. 2005, 15, 1428–1433. [Google Scholar] [CrossRef] [Green Version]
- Bykov, R.A.; Yudina, M.A.; Gruntenko, N.E.; Zakharov, I.K.; Voloshina, M.A.; Melashchenko, E.S.; Danilova, M.V.; Mazunin, I.O.; Ilinsky, Y.Y. Prevalence and genetic diversity of Wolbachia endosymbiont and mtDNA in Palearctic populations of Drosophila melanogaster. BMC Evol. Biol. 2019, 19, 48. [Google Scholar] [CrossRef] [Green Version]
- Ilinsky, Y. Coevolution of Drosophila melanogaster mtDNA and Wolbachia genotypes. PLoS ONE 2013, 8, e54373. [Google Scholar] [CrossRef] [Green Version]
- Braig, H.R.; Zhou, W.; Dobson, S.L.; O’Neill, S.L. Cloning and Characterization of a Gene Encoding the Major Surface Protein of the Bacterial Endosymbiont Wolbachia pipientis. J. Bacteriol. 1998, 180, 2373–2378. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, S.L.; Giordano, R.; Colbert, A.M.E.; Karr, T.L.; Robertson, H.M. 16S rRNA phylogenetic analysis of the bacterial endosymbionts associated with cytoplasmic incompatibility in insects. Proc. Natl. Acad. Sci. USA 1992, 89, 2699–2702. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Mishra, M. Biochemical Estimation to Detect the Metabolic Pathways of Drosophila. In Fundamental Approaches to Screen Abnormalities in Drosophila Springer Protocols Handbooks; Mishra, M., Ed.; Springer: New York, NY, USA, 2020; pp. 135–149. ISBN 978-1-4939-9755-8. [Google Scholar]
- Eremina, M.A.; Gruntenko, N.E. Adaptation of the sulfophosphovanillin method of analysis of total lipids for various biological objects as exemplified by Drosophila melanogaster. Vavilovskii Zhurnal Genetiki i Selektsii = Vavilov J. Genet. Breed. 2020, 24, 441–445. [Google Scholar] [CrossRef]
- Musselman, L.P.; Fink, J.L.; Narzinski, K.; Ramachandran, P.V.; Hathiramani, S.S.; Cagan, R.L.; Baranski, T.J. A high-sugar diet produces obesity and insulin resistance in wild-type Drosophila. Dis. Model. Mech. 2011, 4, 842–849. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Williams, M.J.; Wang, Y.; Klockars, A.; Lind, P.M.; Fredriksson, R.; Schiöth, H.B. Exposure to Bisphenol A Affects Lipid Metabolism in Drosophila melanogaster. Basic Clin. Pharmacol. Toxicol. 2014, 114, 414–420. [Google Scholar] [CrossRef]
- Trinh, I.; Boulianne, G.L. Modeling obesity and its associated disorders in Drosophila. Physiology 2013, 28, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Sun, L.V.; Vamathevan, J.; Riegler, M.; Deboy, R.; Brownlie, J.C.; McGraw, E.A.; Martin, W.; Esser, C.; Ahmadinejad, N.; et al. Phylogenomics of the reproductive parasite Wolbachia pipientis wMel: A streamlined genome overrun by mobile genetic elements. PLoS Biol. 2004, 2, E69. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.B.; Cao, Z.; Qiao, J.X.; Zhong, Z.Q.; Pan, C.C.; Liu, C.; Zhang, L.-M.; Wang, Y.-F. Metabolomics provide new insights into mechanisms of Wolbachia-induced paternal defects in Drosophila melanogaster. PLoS Pathog. 2021, 17, e1009859. [Google Scholar] [CrossRef]
- Da Rocha Fernandes, M.; Martins, R.; Pessoa Costa, E.; Casagrande Pacidônio, E.; Araujo de Abreu, L.; Moreira, L.A.; da Fonseca, R.N.; Logullo, C. The Modulation of the Symbiont/Host Interaction between Wolbachia pipientis and Aedes fluviatilis Embryos by Glycogen Metabolism. PLoS ONE 2014, 9, e98966. [Google Scholar] [CrossRef] [Green Version]
- Pasco, M.Y.; Léopold, P. High Sugar-Induced Insulin Resistance in Drosophila Relies on the Lipocalin Neural Lazarillo. PLoS ONE 2012, 7, e36583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ugrankar, R.; Berglund, E.; Akdemir, F.; Tran, C.; Kim, M.S.; Noh, J.; Schneider, R.; Ebert, B.; Graff, J.M. Drosophila glucome screening identifies Ck1alpha as a regulator of mammalian glucose metabolism. Nat. Commun. 2015, 6, 7102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, M.; Matsuda, H.; Kubo, H.; Nishimura, T. Molecular characterization of Tps1 and Treh genes in Drosophila and their role in body water homeostasis. Sci. Rep. 2016, 6, 30582. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, R.; Hosono, C.; Saigo, K.; Samakovlis, C. The Intersection of the Extrinsic Hedgehog and WNT/Wingless Signals with the Intrinsic Hox Code Underpins Branching Pattern and Tube Shape Diversity in the Drosophila Airways. PLoS Genet. 2015, 11, e1004929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broughton, S.; Alic, N.; Slack, C.; Bass, T.; Ikeya, T.; Vinti, G.; Tommasi, A.M.; Driege, Y.; Hafen, E.; Partridge, L. Reduction of DILP2 in Drosophila triages a metabolic phenotype from lifespan revealing redundancy and compensation among DILPs. PLoS ONE 2008, 3, e3721. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Lu, Y.; Zhang, P.; Zhang, X.; Cao, G.; Xue, R.; Gong, C. Effect of inhibiting the expression of insulin-like peptide gene BBX-B8 on development and reproduction of silkworm, Bombyx mori. Afr. J. Biotechnol. 2012, 11, 2548–2554. [Google Scholar] [CrossRef] [Green Version]
- Iwami, M. Bombyxin: An Insect Brain Peptide That Belongs to the Insulin Family. Zoolog. Sci. 2000, 17, 1035–1044. [Google Scholar] [CrossRef]
- Satake, M.; MacKenzie, L.; Yasumoto, T. Identification of Protoceratium reticulatum as the biogenetic origin of yessotoxin. Nat. Toxins 1997, 5, 164–167. [Google Scholar] [CrossRef]
- Grönke, S.; Clarke, D.F.; Broughton, S.; Andrews, T.D.; Partridge, L. Molecular evolution and functional characterization of Drosophila insulin-like peptides. PLoS Genet. 2010, 6, e1000857. [Google Scholar] [CrossRef] [Green Version]
- Haselton, A.T.; Fridell, Y.W. Adult Drosophila melanogaster as a model for the study of glucose homeostasis. Aging 2010, 2, 523–526. [Google Scholar] [CrossRef] [Green Version]
- Caragata, E.P.; Pais, F.S.; Baton, L.A.; Silva, J.B.; Sorgine, M.H.; Moreira, L.A. The transcriptome of the mosquito Aedes fluviatilis (Diptera: Culicidae), and transcriptional changes associated with its native Wolbachia infection. BMC Genomics 2017, 18, 6. [Google Scholar] [CrossRef] [Green Version]
- Rancès, E.; Ye, Y.H.; Woolfit, M.; McGraw, E.A.; O’Neill, S.L. The relative importance of innate immune priming in Wolbachia-mediated dengue interference. PLoS Pathog. 2012, 8, e1002548. [Google Scholar] [CrossRef] [Green Version]
- Turley, A.P.; Moreira, L.A.; O’Neill, S.L.; McGraw, E.A. Wolbachia infection reduces blood-feeding success in the dengue fever mosquito, Aedes aegypti. PLoS Negl. Trop. Dis. 2009, 3, e516. [Google Scholar] [CrossRef]
- He, Z.; Zhang, H.B.; Li, S.T.; Yu, W.J.; Biwot, J.; Yu, X.Q.; Peng, Y.; Wang, Y.-F. Effects of Wolbachia infection on the postmating response in Drosophila melanogaster. Behav. Ecol. Sociobiol. 2018, 72, 146. [Google Scholar] [CrossRef]
- Ilinsky, Y.Y.; Zakharov, I.K. Cytoplasmic incompatibility in Drosophila melanogaster is caused by different Wolbachia genotypes. Russ. J. Genet. Appl. Res. 2011, 1, 458. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
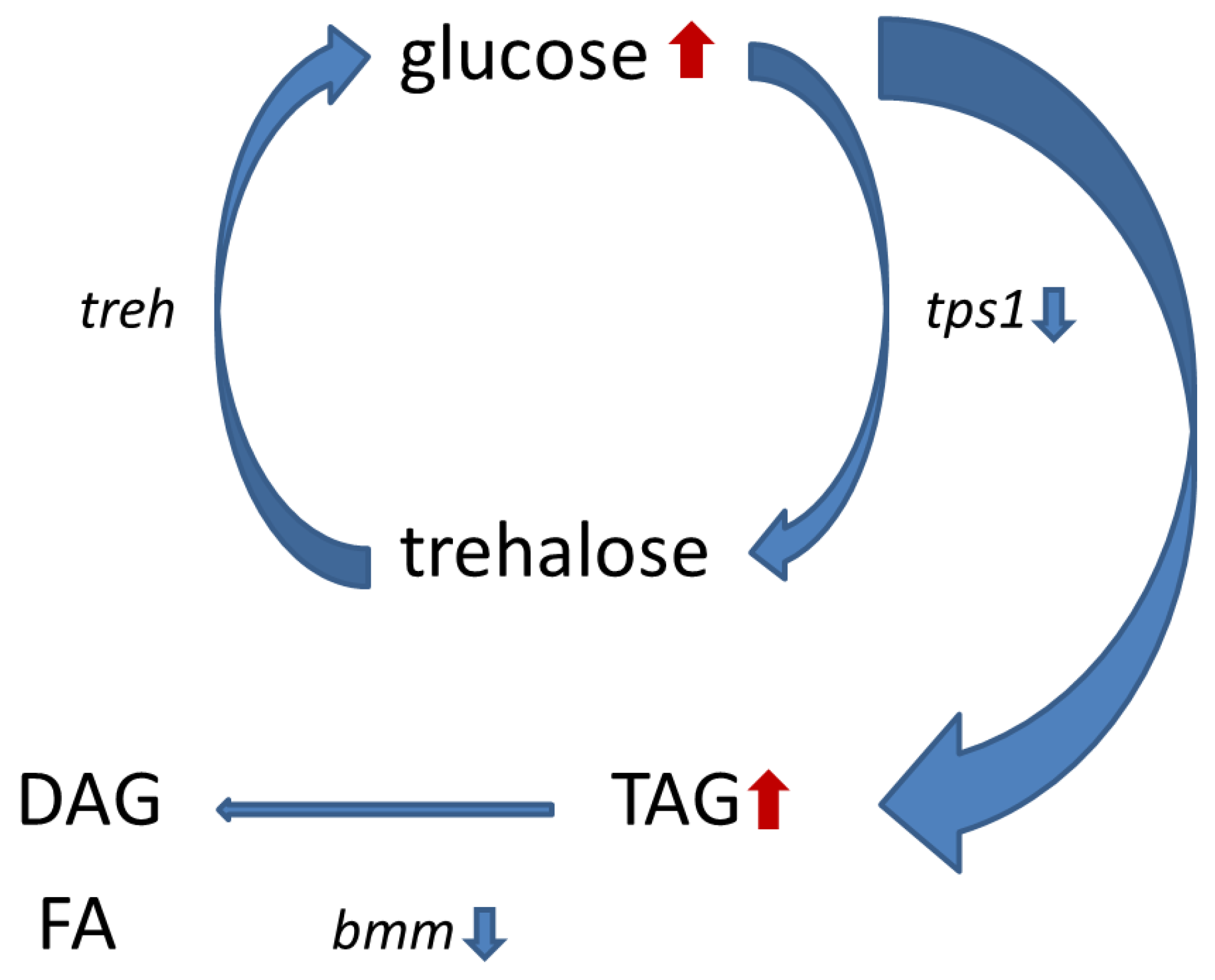{kind=link}
| Drosophila Line | Wolbachia Infection | Line-Donor of Cytoplasm | Origin of Donor Line |
|---|---|---|---|
| Bi90T | --- | Bi90, tetracycline treated | Kyrgyzstan, 2004 |
| Bi90wMel | wMel | Bi90 | Kyrgyzstan, 2004 |
| Bi90wMelPlus | wMelPlus | w153 | Uzbekistan, 1989 |
| Bi90wMelCS128 | wMelCS | 1-128 | Australia, 1986 |
| Bi90wMelCS45 | wMelCS | 45 | Sankt-Peterburg, Russia, 1995 |
| Bi90wMelCS112 | wMelCS | 3-112 | Bloomington, USA, 2010 |
| Bi90wMelCS2 | wMelCS2 | 93220 | Biysk, Altai, Russia, 1993 |
| Bi90wMel4 | wMel4 | w304 | Sinai Peninsula, Egypt, 2010 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karpova, E.K.; Bobrovskikh, M.A.; Deryuzhenko, M.A.; Shishkina, O.D.; Gruntenko, N.E. Wolbachia Effect on Drosophila melanogaster Lipid and Carbohydrate Metabolism. Insects 2023, 14, 357. https://doi.org/10.3390/insects14040357
Karpova EK, Bobrovskikh MA, Deryuzhenko MA, Shishkina OD, Gruntenko NE. Wolbachia Effect on Drosophila melanogaster Lipid and Carbohydrate Metabolism. Insects. 2023; 14(4):357. https://doi.org/10.3390/insects14040357
Chicago/Turabian StyleKarpova, Evgenia K., Margarita A. Bobrovskikh, Maksim A. Deryuzhenko, Olga D. Shishkina, and Nataly E. Gruntenko. 2023. "Wolbachia Effect on Drosophila melanogaster Lipid and Carbohydrate Metabolism" Insects 14, no. 4: 357. https://doi.org/10.3390/insects14040357