
Phenological Features of the Spongy Moth, Lymantria dispar (L.) (Lepidoptera: Erebidae), in the Northernmost Portions of Its Eurasian Range
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods

3. Results
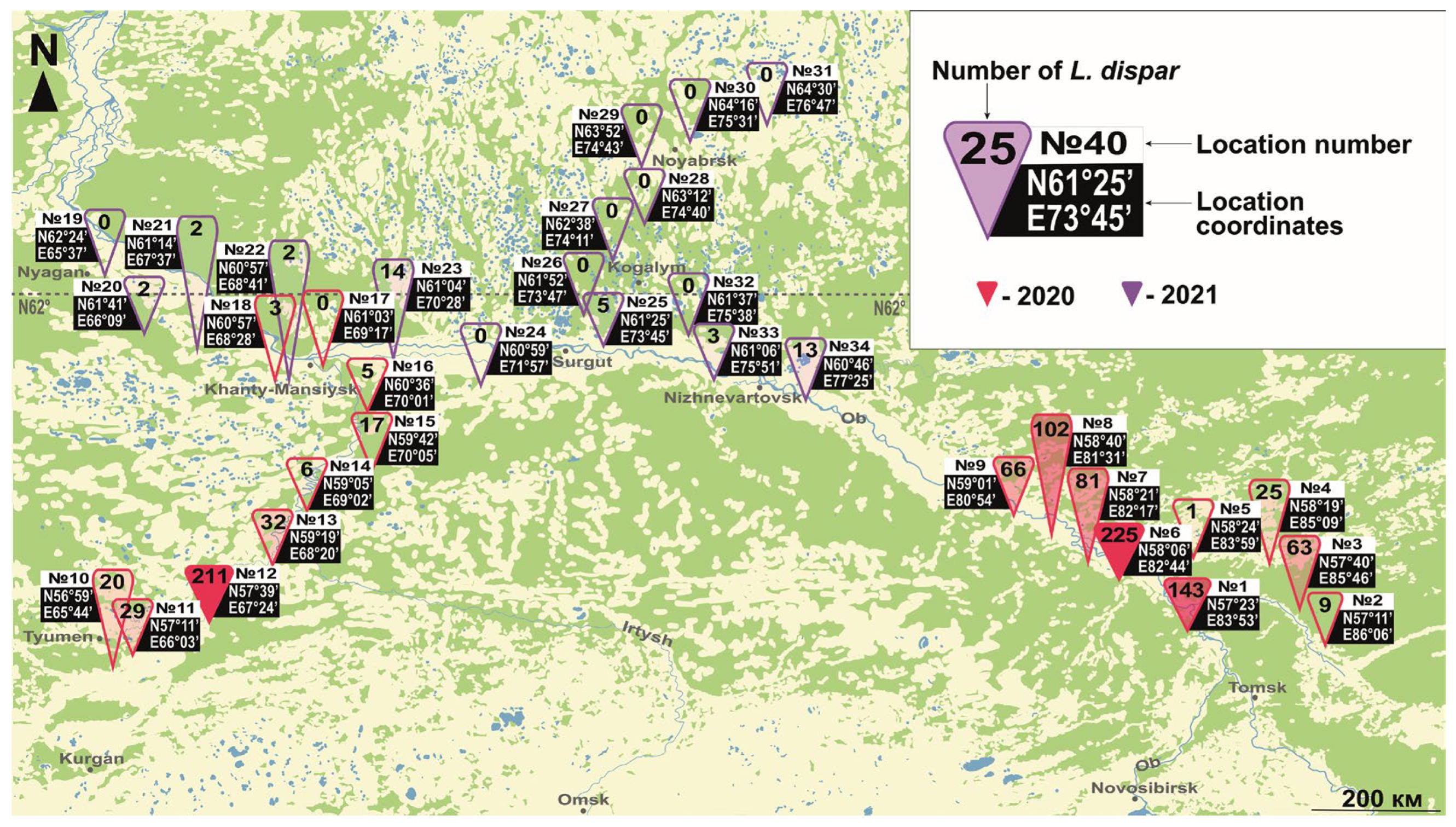
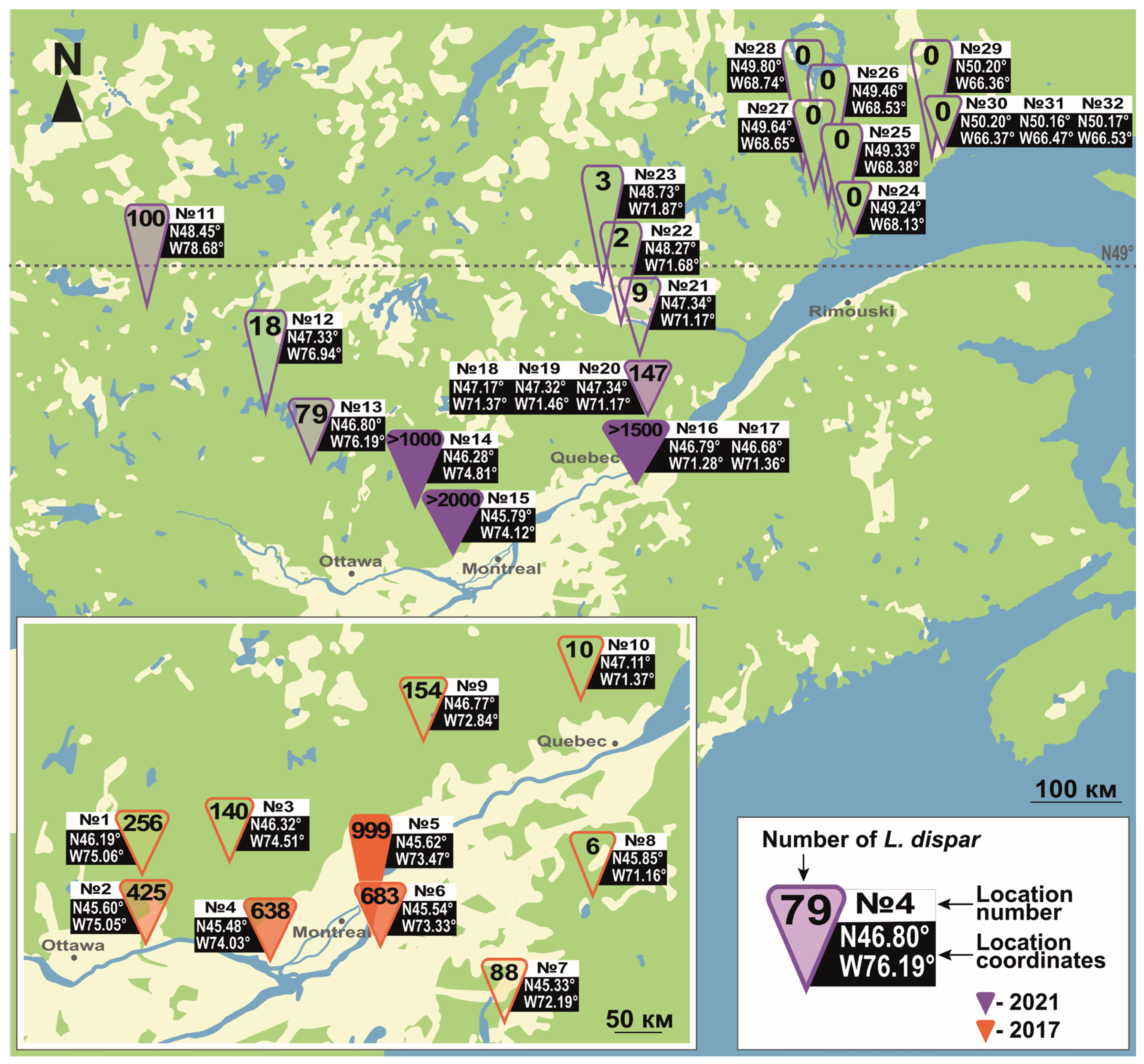
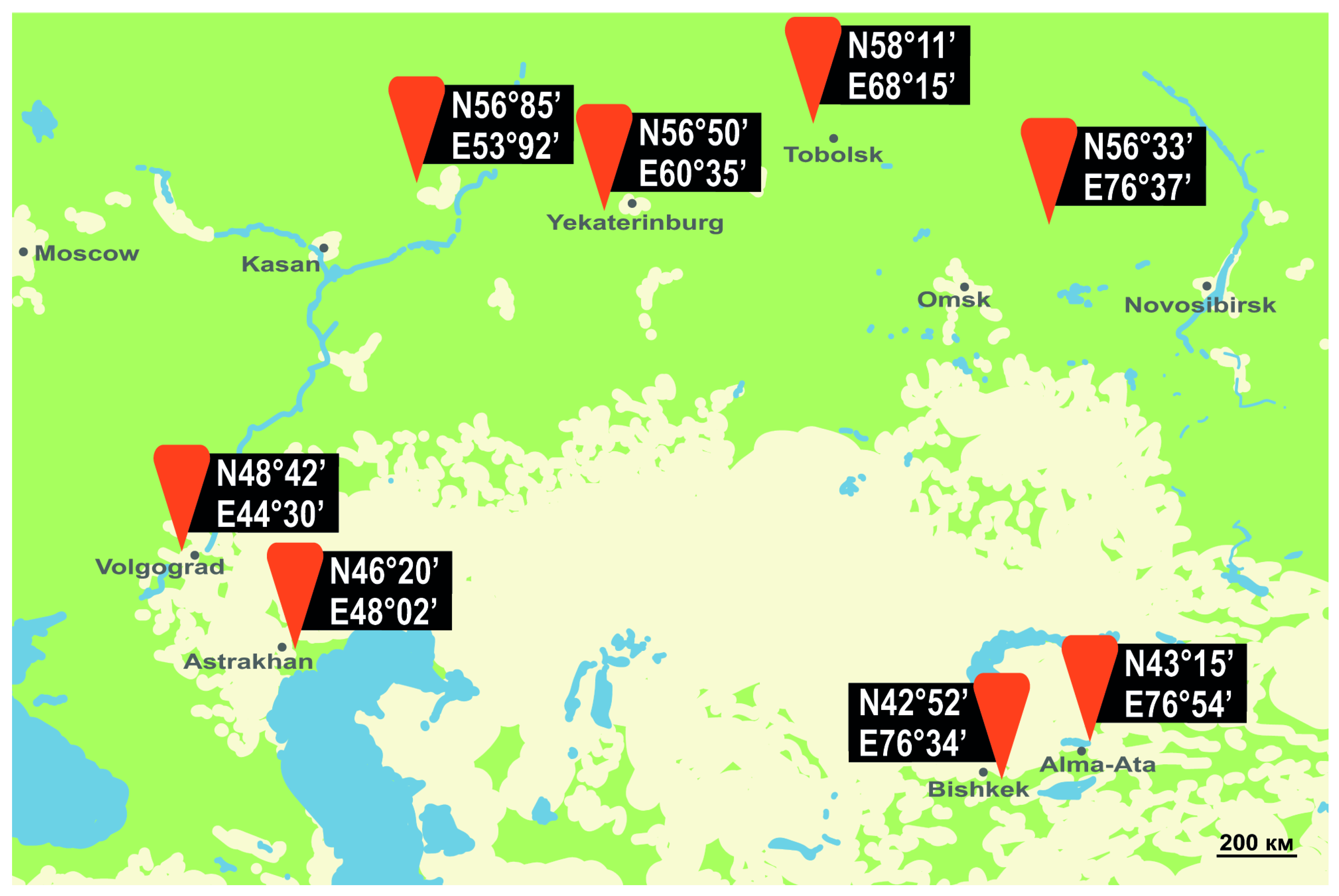
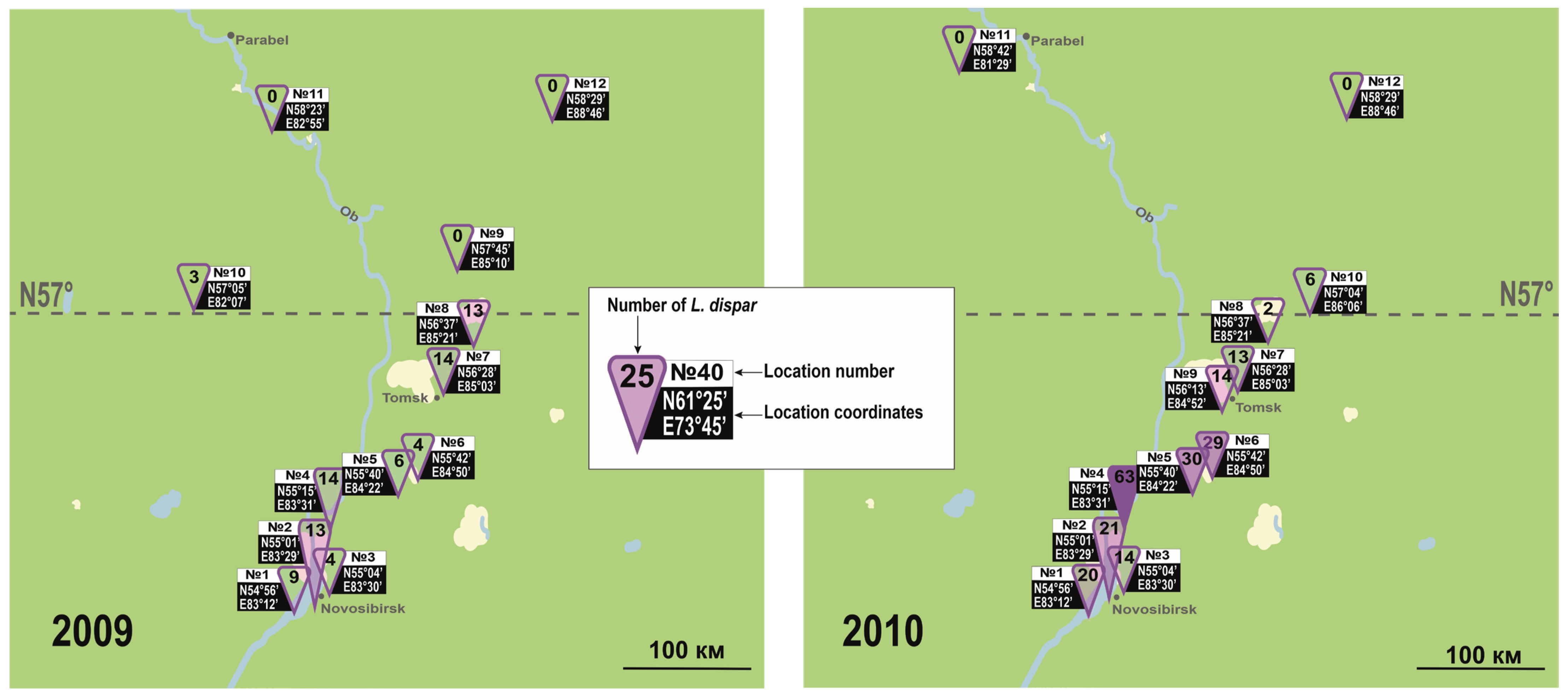
3.1. Northern Limit of L. dispar in Eurasia and North America
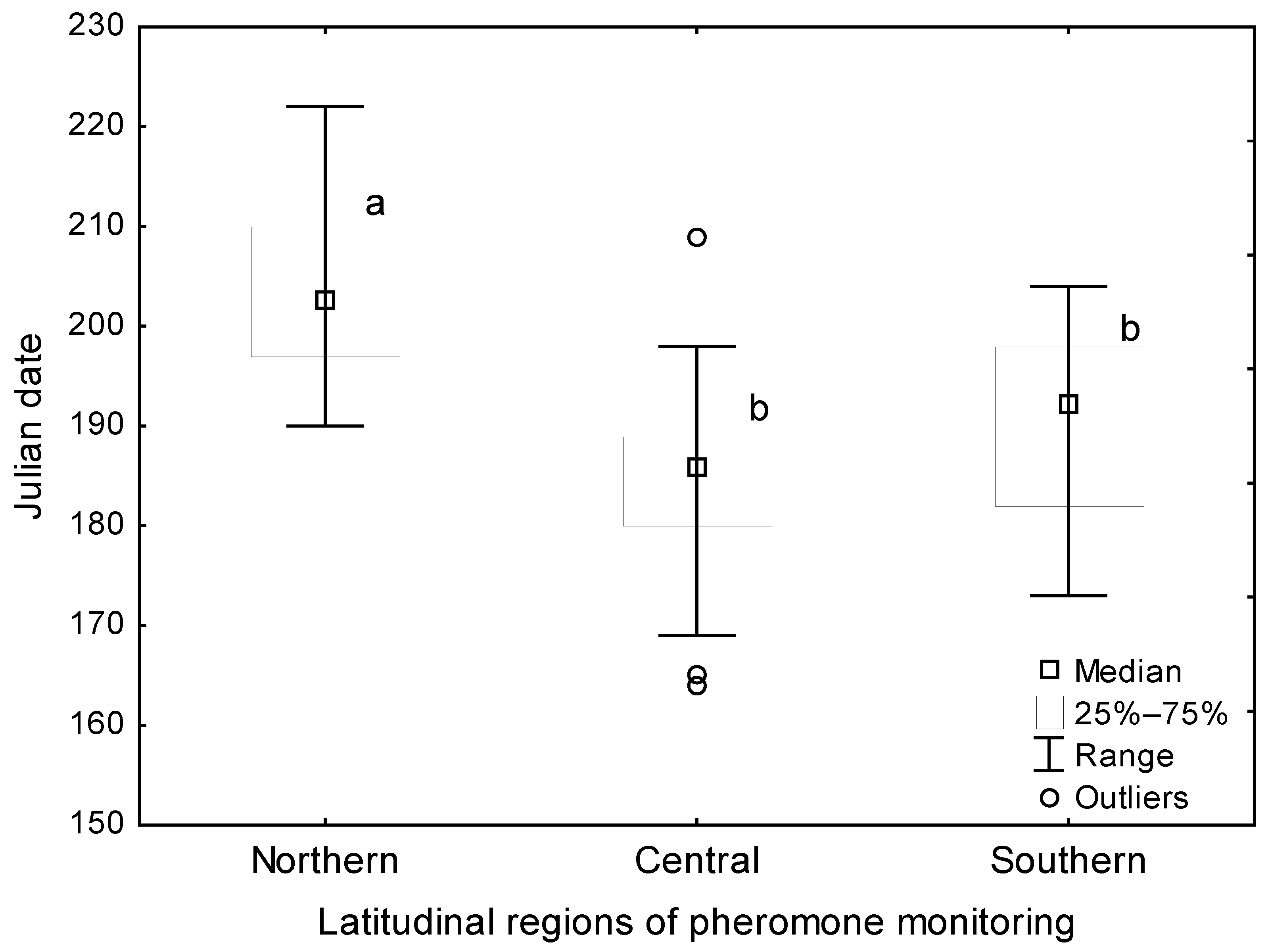
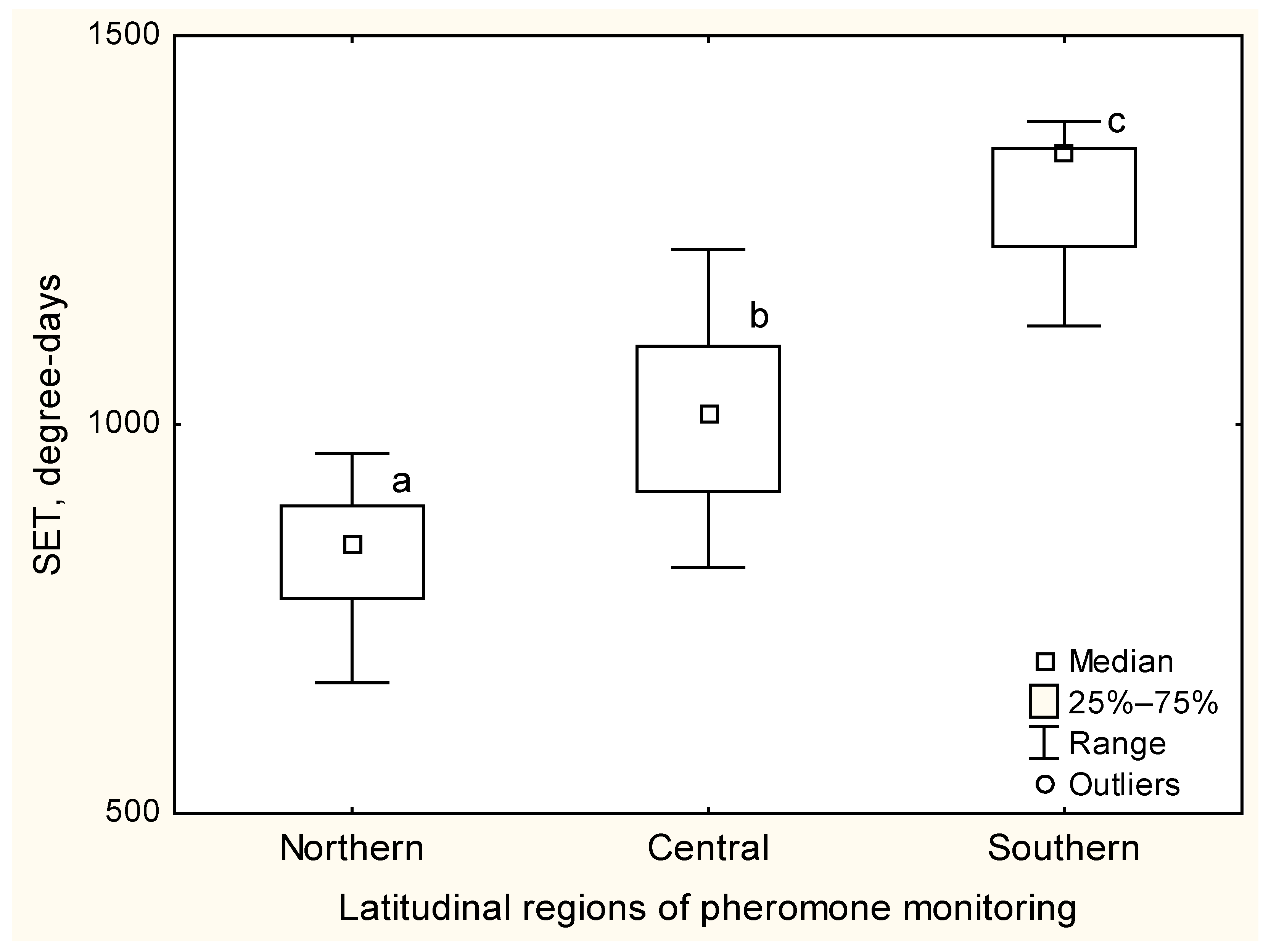
3.2. Flight Phenology, SET and Heat Availability
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leonard, D.E. Bioecology of the gypsy moth. In Gypsy Moth Research towards Integrated Pest Management; Doane, C.C., McManus, M.L., Eds.; U.S. Department of Agriculture: Washington, DC, USA, 1981; pp. 9–29. [Google Scholar]
- Myers, H.S. Synchrony in outbreaks of forest lepidoptera: A possible example of the Moran effect. Ecology 1998, 79, 1111–1117. [Google Scholar] [CrossRef]
- Johnson, D.M.; Liebhold, A.M.; Bjørnstad, O.N.; Mcmanus, M.L. Circumpolar variation in periodicity and synchrony among gypsy moth populations. J. Anim. Ecol. 2005, 74, 882–892. [Google Scholar] [CrossRef]
- Soukhovolsky, V.G.; Ponomarev, V.I.; Sokolov, G.I.; Tarasova, O.V.; Krasnoperova, P.A. Gypsy moth Lymantria dispar L. in the South Urals: Patterns in population dynamics and modeling. Biol. Bull. Rev. 2016, 6, 57–69. [Google Scholar] [CrossRef]
- Martemyanov, V.; Bykov, R.; Demenkova, M.; Gninenko, Y.; Romancev, S.; Bolonin, I.; Mazunin, I.; Belousova, I.; Akhanaev, Y.; Pavlushin, S.; et al. Genetic evidence of broad spreading of Lymantria dispar in the West Siberian Plain. PLoS ONE 2019, 14, e0220954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keena, M.A.; Richards, J.Y. Comparison of survival and development of gypsy moth Lymantria dispar L. (Lepidoptera: Erebidae) populations from different geographic areas on North American conifers. Insects 2020, 11, 260. [Google Scholar] [CrossRef] [Green Version]
- Lowe, S.; Browne, M.; Boudjelas, S.; De Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; IUCN Species Survival Commission, Invasive Species Specialist Group: Auckland, New Zealand, 2001; Available online: http://www.iucngisd.org/gisd/pdf/100English.pdf (accessed on 10 January 2022).
- Liebhold, A.; Mastro, V.; Schaefer, P.W. Learning from the legacy of Leopold Trouvelot. Bull. Entomol. Soc. Am. 1989, 35, 20–22. [Google Scholar] [CrossRef]
- Giese, R.L.; Schneider, M.L. Cartographic comparisons of Eurasian gypsy moth distribution (Lymantria dispar L. Lepidoptera: Lymantriidae). Entomol. News 1979, 90, 1–16. [Google Scholar]
- Mecellem, D.; Chakali, G. Biological and ecological characteristics of Gypsy Moth, Lymantria dispar L. (Lep. Lymantriidae) in the gradation phase’s in Blidéen Atlas Forest (Algeria). Adv. Environ. Biol. 2016, 10, 192–199. [Google Scholar]
- Gninenko, Y.I.; Kavosi, M.R. Lymantria dispar (Lepidoptera, Erebidae) outbreak in north Iran. For. Inf. 2016, 2, 81–89. (In Russian) [Google Scholar]
- Ashimov, K.S. Dendrophilous Insects of Walnut-Fruit Forests of the Southwestern Tien Shan; Kyrgyz National Agrarian University: Bishkek, Kyrgyzstan, 2005. (In Russian) [Google Scholar]
- Klapwijk, M.J.; Csóka, G.; Hirka, A.; Björkman, C. Forest insects and climate change: Long-term trends in herbivore damage. Ecol. Evol. 2013, 3, 4183–4196. [Google Scholar] [CrossRef] [Green Version]
- Kurenshchikov, D.K.; Ilyinykh, A.V. Gypsy moth (Lymantria dispar) male flight features for the south of the Russian Far East. Mod. Probl. Sci. Educ. 2015, 4, 443–451. (In Russian) [Google Scholar]
- Meshkova, V.L. Seasonal Development of Phyllophagous Insects; Planeta-Print: Kharkiv, Ukraine, 2009. (In Russian) [Google Scholar]
- Ilyinskiy, A.I. Supervision, Accounting and Forecast of Mass Propagation of Needle- and Leaf-Eating Insects in the Forests of the USSR; Forest Industry: Moscow, Russia, 1965. (In Russian) [Google Scholar]
- Gray, D.R. Age-dependent postdiapause development in the gypsy moth (Lepidoptera: Lymantriidae) life stage model. Environ. Entomol. 2009, 38, 18–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauffman, B.W.; Clatterbuck, W.K.; Liebhold, A.M.; Coyle, D.R. Gypsy Moth in the Southeastern U.S.: Biology, Ecology and Forest Management Strategies; SREF-FH-008; Southern Regional Extension Forestry: Athens, GA, USA, 2017; 10p. [Google Scholar]
- Gray, D.R. The gypsy moth life stage model: Landscapewide estimates of gypsy moth establishment using a multigenerational phenology model. Ecol. Model. 2004, 176, 155–171. [Google Scholar] [CrossRef]
- Régnière, J.; Sharov, A.A. Simulating temperature-dependent ecological processes at the sub-continental scale: Male gypsy moth flight phenology as an example. Int. J. Biometeorol. 1999, 42, 146–152. [Google Scholar] [CrossRef]
- Tobin, P.C.; Cremers, K.T.; Hunt, L.; Parry, D. All quiet on the western front? Using phenological inference to detect the presence of a latent gypsy moth invasion in Northern Minnesota. Biol. Invasions 2016, 18, 3561–3573. [Google Scholar] [CrossRef]
- Djoumad, A.; Nisole, A.; Zahiri, R.; Freschi, L.; Picq, S.; Gundersen-Rindal, D.E.; Sparks, M.; Dewar, K.; Stewart, D.; Maaroufi, D.; et al. Comparative analysis of mitochondrial genomes of geographic variants of the gypsy moth, Lymantria dispar, reveals a previously undescribed genotypic entity. Sci. Rep. 2017, 7, 14245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picq, S.; Yunke, W.; Martemyanov, V.V.; Pouliot, E.; Pfister, S.E.; Hamelin, R.; Cusson, M. Range-wide population genomics of the spongy moth, Lymantria dispar (Erebidae): Implications for biosurveillance, subspecies classification and phylogeography of a destructive moth. Evol. Appl. 2023, 1–19. [Google Scholar] [CrossRef]
- Ponomarev, V.I.; Klobukov, G.I.; Napalkova, V.V. The dependence of the morphological and physiological indices of Lymantria dispar (L.) (Lepidoptera: Lymantriidae) postembryonic stages from temperature conditions during embryogenesis. Tomsk State Univ. J. Biol. 2016, 3, 107–127. (In Russian) [Google Scholar] [CrossRef] [Green Version]
- Ermolaev, I.V.; Ponomarev, V.I.; Vasil’ev, A.A.; Kumaeva, M.S. Phytophagous insects associated with the pedunculate oak (Quercus robur) in the northeast of its distribution area. Entomol. Rev. 2021, 101, 437–449. [Google Scholar] [CrossRef]
- Yasyukevich, V.V.; Titkina, C.N.; Davidovich, E.A.; Yasyukevich, N.V. Changes in boundaries of gypsy moth and nun moth (Lymantria dispar and Lymantria monacha, Lymantriidae, Lepidoptera) ranges due to the global warming: A model approach. Zool. Zh. 2013, 92, 1377–1382. [Google Scholar] [CrossRef]
- Ilyinykh, A.V.; Krivets, S.A. The results of gypsy moth Lymantria dispar (L.) (Lepidoptera, Lymantriidae) pheromone monitoring in southeast of West Siberia. Bull. SPbFTU 2011, 196, 45–53. (In Russian) [Google Scholar]
- Shapiro, M.; Bell, A.; Owens, C.D. In vivo mass production of gypsy moth nucleopolyhedrosis virus. In Gypsy Moth Research towards Integrated Pest Management; Doane, C.C., McManus, M.L., Eds.; U.S. Department of Agriculture: Washington, DC, USA, 1981; pp. 633–657. [Google Scholar]
- Ponomarev, V.I.; Klobukov, G.I.; Napalkova, V.V.; Tyurin, M.V.; Martemyanov, V.V. Influence of biotic and abiotic factors on the duration of development of the spongy moth Lymantria dispar (L.) (Lepidoptera: Erebidae) in the West Siberian population of different latitudinal origin. Contemp. Probl. Ecol. 2023, 16, 166–172. [Google Scholar]
- Mastro, V.C. Evaluation of disparlure-baited traps. In Gypsy Moth Research towards Integrated Pest Management; Doane, C.C., McManus, M.L., Eds.; U.S. Department of Agriculture: Washington, DC, USA, 1981; pp. 549–553. [Google Scholar]
- Frank, K.L.; Tobin, P.C.; Thistle, H.W.; Kalkstein, L.S. Interpretation of gypsy moth frontal advance using meteorology in a conditional algorithm. Int. J. Biometeorol. 2012, 57, 459–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Régnière, J.; Sharov, A.A. Phenology of Lymantria dispar (Lepidoptera: Lymantriidae), male flight and the effect of moth dispersal in heterogeneous landscapes. Int. J. Biometeorol. 1998, 41, 161–168. [Google Scholar] [CrossRef]
- Johnson, P.C.; Mason, D.P.; Radke, S.L.; Tracewski, K.T. Gypsy moth, Lymantria dispar (L.) (Lepidoptera: Lymantriidae), egg eclosion: Degree-day accumulations. Environ. Entomol. 1983, 12, 929–932. [Google Scholar] [CrossRef]
- Pantyukhov, G.A. The influence of positive temperatures on various geographical populations of Euproctis chrysorrhoea L. and gypsy moth Lymantria dispar L. (Lepidoptera, Orgyidae). Entomol. Rev. 1962, 41, 274–284. [Google Scholar]
- Carter, M.R.; Ravlin, F.W.; McManus, M.L. Effect of defoliation on gypsy moth phenology and capture of male moths in pheromone-baited traps. Environ. Entomol. 1992, 21, 1308–1318. [Google Scholar] [CrossRef]
- Vanhanen, H.; Veteli, T.O.; Päivinen, S.; Kellomäki, S.; Niemelä, P. Climate change and range shifts in two insect defoliators: Gypsy moth and nun moth—A model study. Silva Fenn. 2007, 41, 621–638. [Google Scholar] [CrossRef] [Green Version]
- Limbu, S.; Keena, M.; Chen, F.; Cook, G.; Nadel, H.; Hoover, K. Effects of temperature on development of Lymantria dispar asiatica and Lymantria dispar japonica (Lepidoptera: Erebidae). Environ. Entomol. 2017, 46, 1012–1023. [Google Scholar] [CrossRef]
- Kozhanchikov, I.V. Fauna of the USSR. Lepidoptera Insects. Tussock Moths (Orgyidae); Academy of Sciences USSR: Moscow, Russia, 1950. (In Russian) [Google Scholar]
- Gray, D.R.; Ravlin, F.W.; Braine, J.A. Diapause in the gypsy moth: A model of inhibition and development. J. Insect Physiol. 2001, 47, 173–184. [Google Scholar] [CrossRef]
- Weather and Climate. Available online: http://www.pogodaiklimat.ru (accessed on 10 January 2022). (In Russian).
- Weather Schedule. Available online: https://rp5.ru/ (accessed on 10 January 2022). (In Russian).
- Historical Climate Data. Available online: https://climate.weather.gc.ca (accessed on 10 January 2022).
- National Weather Service. Available online: https://www.weather.gov (accessed on 10 January 2022).
- Régnière, J.; Nealis, V.; Porter, K. Climate suitability and management of the gypsy moth invasion into Canada. Biol. Invasions 2009, 11, 135–148. [Google Scholar] [CrossRef]
- Keena, M.A.; Odell, T.M. Effects of Laboratory Rearing on Gypsy Moth (Lepidoptera: Lymantriidae); General Technical Report NE-181; Northeastern Forest Experiment Station, USDA Forest Service: Radnor, PA, USA, 1992; 23p.
- Thompson, L.M.; Faske, T.M.; Banahene, N.; Grim, D.; Agosta, S.J.; Parry, D.; Tobin, P.C.; Johnson, D.M.; Grayson, K.L. Variation in growth and developmental responses to supraoptimal temperatures near latitudinal range limits of gypsy moth Lymantria dispar (L.), an expanding invasive species. Physiol. Entomol. 2017, 42, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Trotter, R.T.; Limbu, S.; Hoover, K.; Nadel, H.; Keena, M.A. Comparing Asian gypsy moth [Lymantria dispar asiatica (Lepidoptera: Erebidae) and L. dispar japonica] trap data from East Asian ports with lab parameterized phenology models: New tools and questions. Ann. Entomol. Soc. Am. 2020, 113, 125–138. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, K.A. User’s Guide for GMPHEN: Gypsy Moth Phenology Model; Technical Report NE–158; U.S. Department of Agriculture, Forest Service: Radnor, PA, USA, 1992; 29p.
- Thompson, L.M.; Powers, S.D.; Appolon, A.; Hafker, P.; Milner, L.; Parry, D.; Agosta, S.J.; Grayson, K.L. Climate-related geographical variation in performance traits across the invasion front of a widespread non-native insect. J. Biogeogr. 2021, 48, 405–414. [Google Scholar] [CrossRef]
- Srivastava, V.; Griess, V.C.; Keena, M.A. Assessing the potential distribution of Asian gypsy moth in Canada: A comparison of two methodological approaches. Sci. Rep. 2020, 10, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, V.; Keena, M.A.; Maennicke, G.E.; Hamelin, R.C.; Griess, V.C. Potential differences and methods of determining gypsy moth female flight capabilities: Implications for the establishment and spread in novel habitats. Forests 2021, 12, 103. [Google Scholar] [CrossRef]
- Soukhovolsky, V.; Tarasova, O.; Pavlushin, S.; Osokina, E.; Akhanaev, Y.; Kovalev, A.; Martemyanov, V. Economics of a feeding budget: A case of diversity of host plants for Lymantria dispar L. (Lepidoptera) feeding on leaves and needles. Diversity 2023, 15, 102. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Flight Dates | Flight Duration, Days | Number of Males, Per Trap | ||
|---|---|---|---|---|---|
| Start | End | Median | |||
| 58°11′ N–68°15′ E—Tyumen region, Tobolsk, Siberia | |||||
| 2020 | 12 July | 22 July | 16 July | 11 | 65 |
| 2021 | 12 July | 12 August | 20 July | 32 | 49 |
| 56°85′ N–53°92′ E—Republic of Udmurtia, Votkinsky district, biostation “Siva”, Ural 1 | |||||
| 2014 | 15 July | 9 August | 2 August | 26 | 20 |
| 2015 | 5 July | 5 August | 22 July | 32 | 18 |
| 2016 | 7 July | 1 August | 12 July | 26 | 12 |
| 56°50′ N–60°35′ E—Sverdlovsk region, Ekaterinburg, Ural 2 | |||||
| 2009 | 23 July | 10 August | 25 July | 18 | 176 |
| 2010 | 3 July | 7 August | 22 July | 36 | 182 |
| 2011 | 19 July | 12 August | 28 July | 25 | 30 |
| 2012 | 3 July | 18 July | 11 July | 16 | 17 |
| 2013 | 25 July | 14 August | 29 July | 21 | 799 |
| 2014 | 31 July | 19 August | 9 August | 20 | 7 |
| 2015 | 16 July | 24 July | 23 July | 9 | 3 |
| 2016 | 8 July | 31 July | 20 July | 24 | 575 |
| 2017 | 25 July | 28 August | 8 August | 35 | 66 |
| 2018 | 1 August | 6 August | 2 August | 6 | 4 |
| 2019 | 15 July | 19 July | 17 July | 5 | 2 |
| 56°33′ N–76°37′ E—Novosibirsk region, Kyshtovka village, Siberia | |||||
| 2019 | - 3 | 12 August | 30 July | ~20 | 3399 |
| 2020 | 6 July | 26 July | 17 July | 20 | 445 |
| 2021 | 5 July | 8 August | 15 July | 35 | 1166 |
| 2022 | 11 July | 14 August | 31 July | 33 | 4223 |
| 53°74′ N–78°00′ E—Novosibirsk region, Karasuk, Siberia | |||||
| 2013 | 10 July | 16 August | 29 July | 37 | 585 |
| 2022 | 7 July | 4 August | 12 July | 28 | 1454 |
| 48°42′ N–44°30′ E—Volgograd region, Volgograd, Eastern Europe | |||||
| 2010 | 2 July | 31 July | 6 July | 30 | 14 |
| 2011 | 8 July | 28 July | 15 July | 21 | 46 |
| 2012 | 7 July | 27 July | 18 July | 21 | 152 |
| 2014 | 19 June | 23 July | 4 July | 35 | 654 |
| 46°20′ N–48°02′ E—Astrakhan region, Astrakhan, Eastern Europe | |||||
| 2011 | 28 June | 5 August | 9 July | 39 | 814 |
| 2013 | 14 June | 29 July | 6 July | 45 | 424 |
| 2014 | 18 June | 20 July | 1 July | 33 | 478 |
| 43°15′ N–76°54′ E—Kazakhstan, Alma-Ata, Central Asia | |||||
| 2007 | 3 July | 3 August | 15 July | 31 | 170 |
| 42°52′ N–76°34′ E—Republic of Kyrgyzstan, Bishkek, Central Asia | |||||
| 2009 | 3 July | 8 August | 23 July | 37 | 409 |
| 2012 | 25 June | 30 July | 24 July | 36 | 2956 |
| 2013 | 24 June | 5 August | 18 July | 43 | 367 |
| 40°56′ N–E73°00′ E—Republic of Kyrgyzstan, Jalal-Abad, Central Asia | |||||
| 2004 | 27 June | 18 July | 6 July | 21 | 57 |
| 2005 | 21 June | 24 July | 6 July | 33 | 164 |
| 2017 | 11 June | 26 July | 23 June | 45 | 4498 |
| Year | SET for Development to Imago Stage | SET | ||||
|---|---|---|---|---|---|---|
| At Beginning of Flight | At End of Flight | At Flight Median | Summer–Autumn Embryo Development | Heat Availability | ||
| At Flight Median | At End of Flight | |||||
| 58°11′ N–68°15′ E—Tyumen region, Tobolsk (WMO id 28275 1) | ||||||
| 2020 | 705 | 865 | 771 | 596 | 502 | 1367 |
| 2021 | 715 | 1017 | 776 | 517 | 276 | 1293 |
| 56°85′ N–53°92′ E—Republic of Udmurtia, biostation “Siva” (WMO id 28413) | ||||||
| 2014 | 756 | 1014 | 910 | 461 | 357 | 1371 |
| 2015 | 698 | 1004 | 850 | 551 | 397 | 1401 |
| 2016 | 686 | 1066 | 782 | 904 | 620 | 1686 |
| 56°50′ N–60°35′ E—Sverdlovsk region, Ekaterinburg (WMO id 28440) | ||||||
| 2009 | 738 | 892 | 757 | 549 | 414 | 1306 |
| 2010 | 659 | 1176 | 897 | 564 | 285 | 1461 |
| 2011 | 700 | 989 | 835 | 449 | 295 | 1284 |
| 2012 | 781 | 929 | 900 | 744 | 715 | 1644 |
| 2013 | 826 | 1067 | 880 | 491 | 304 | 1371 |
| 2014 | 763 | 976 | 868 | 273 | 165 | 1141 |
| 2015 | 713 | 786 | 776 | 411 | 401 | 1187 |
| 2016 | 687 | 989 | 848 | 769 | 628 | 1617 |
| 2017 | 655 | 1041 | 824 | 347 | 130 | 1171 |
| 2018 | 766 | 816 | 777 | 426 | 387 | 1203 |
| 2019 | 667 | 723 | 691 | 537 | 505 | 1228 |
| 56°33′ N–76°37′ E—Novosibirsk region, Kyshtovka village (WMO id 29405) | ||||||
| 2019 | - | 846 | 709 | 403 | 265 | 1111 |
| 2020 | 646 | 923 | 801 | 547 | 416 | 1348 |
| 2021 | 563 | 921 | 669 | 538 | 286 | 1207 |
| 2022 | 603 | 965 | 817 | 356 | 208 | 1173 |
| 53°74′ N–78°02′ E—Novosibirsk region, Karasuk (WMO id 29814) | ||||||
| 2013 | 554 | 1043 | 816 | 471 | 244 | 1287 |
| 2022 | 736 | 1211 | 909 | 710 | 408 | 1619 |
| 48°42′ N–44°30′ E—Volgograd region, Volgograd (WMO id 34560) | ||||||
| 2010 | 974 | 1374 | 1102 | 1296 | 931 | 2305 |
| 2011 | 936 | 1364 | 1112 | 1020 | 768 | 2132 |
| 2012 | 738 | 1029 | 887 | 1564 | 1422 | 2451 |
| 2014 | 787 | 1264 | 957 | 1247 | 940 | 2204 |
| 46°20′ N–48°02′ E—Astrakhan region, Astrakhan (WMO id 34880) | ||||||
| 2011 | 788 | 1533 | 1085 | 1243 | 795 | 2328 |
| 2013 | 792 | 1503 | 1191 | 1247 | 935 | 2438 |
| 2014 | 834 | 1429 | 1194 | 1318 | 1083 | 2512 |
| 43°15′ N–76°54′ E—Kazakhstan, Alma-Ata (WMO id 36870) | ||||||
| 2007 | 1074 | 1534 | 1203 | 1194 | 863 | 2397 |
| 42°52′ N–76°34′ E—Republic of Kyrgyzstan, Bishkek (WMO id 38353) | ||||||
| 2009 | 911 | 1533 | 1237 | 988 | 692 | 2225 |
| 2012 | 1097 | 1721 | 1342 | 1288 | 909 | 2630 |
| 2013 | 881 | 1604 | 1195 | 1222 | 813 | 2417 |
| 40°56′ N–E73°00′ E—Republic of Kyrgyzstan, Jalal-Abad (WMO id 38613) | ||||||
| 2004 | 1175 | 1544 | 1337 | 1672 | 1465 | 3009 |
| 2005 | 1080 | 1710 | 1350 | 1739 | 1379 | 3089 |
| 2017 | 898 | 1694 | 1114 | 1357 | 1937 | 3051 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponomarev, V.I.; Klobukov, G.I.; Napalkova, V.V.; Akhanaev, Y.B.; Pavlushin, S.V.; Yakimova, M.E.; Subbotina, A.O.; Picq, S.; Cusson, M.; Martemyanov, V.V. Phenological Features of the Spongy Moth, Lymantria dispar (L.) (Lepidoptera: Erebidae), in the Northernmost Portions of Its Eurasian Range. Insects 2023, 14, 276. https://doi.org/10.3390/insects14030276
Ponomarev VI, Klobukov GI, Napalkova VV, Akhanaev YB, Pavlushin SV, Yakimova ME, Subbotina AO, Picq S, Cusson M, Martemyanov VV. Phenological Features of the Spongy Moth, Lymantria dispar (L.) (Lepidoptera: Erebidae), in the Northernmost Portions of Its Eurasian Range. Insects. 2023; 14(3):276. https://doi.org/10.3390/insects14030276
Chicago/Turabian StylePonomarev, Vasiliy I., Georgiy I. Klobukov, Viktoria V. Napalkova, Yuriy B. Akhanaev, Sergey V. Pavlushin, Maria E. Yakimova, Anna O. Subbotina, Sandrine Picq, Michel Cusson, and Vyacheslav V. Martemyanov. 2023. "Phenological Features of the Spongy Moth, Lymantria dispar (L.) (Lepidoptera: Erebidae), in the Northernmost Portions of Its Eurasian Range" Insects 14, no. 3: 276. https://doi.org/10.3390/insects14030276