

Species Richness of Papilionidae Butterflies (Lepidoptera: Papilionoidea) in the Hengduan Mountains and Its Future Shifts under Climate Change
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Research Area
2.2. Species Records
2.3. Environmental Variables
2.4. Maxent Simulation and Spatial Richness Analysis
3. Results
3.1. Model Performance and Key BioClim Variables
3.2. Current Distribution Pattern
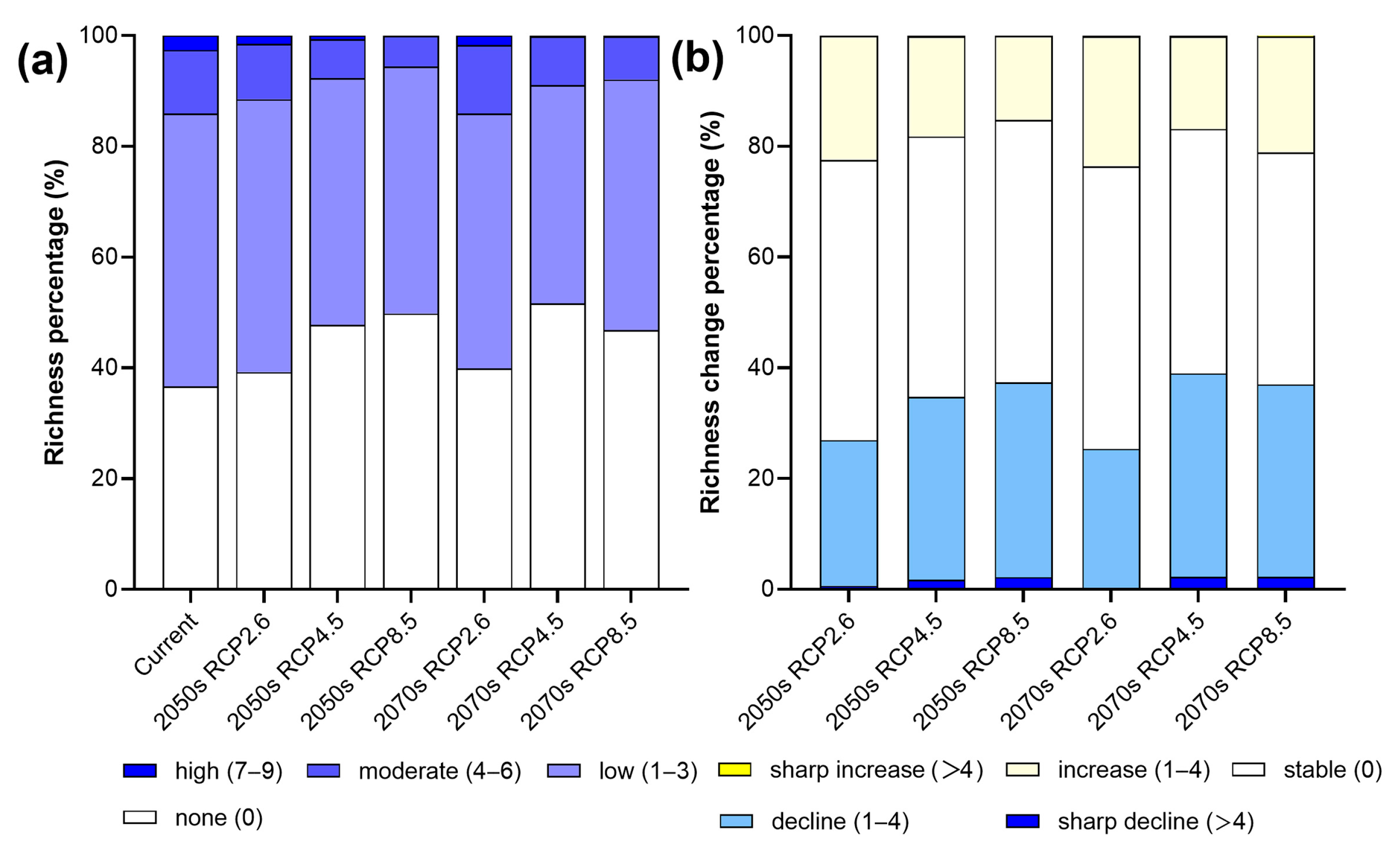
3.3. Future Changes
3.3.1. Parnassiinae
3.3.2. Papilioninae
4. Discussion
4.1. Causes Underlying Species Richness Pattern
4.2. Climate Vulnerability
4.3. Conservation Implications
5. Conclusions and Limitation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, W.L.; Suman, D.O.; Zhang, H.H.; Xu, Z.B.; Ma, F.Z.; Hu, S.J. Butterfly conservation in China: From science to action. Insects 2020, 11, 661. [Google Scholar] [CrossRef] [PubMed]
- CITES. Convention on International Trade in Endangered Species of Wild Fauna and Flora. Appendices I, II, and III. Available online: https://cites.org/eng/app/appendices.php (accessed on 10 April 2022).
- IUCN. IUCN Red List of Threatened Species. Available online: https://www.iucn.org/resources/conservation-tools/iucn-red-list-threatened-species (accessed on 10 April 2022).
- NFGA; MARA. List of Wildlife under Special State Protection. Available online: http://www.forestry.gov.cn/main/5461/20210205/122418860831352.html (accessed on 10 April 2022).
- Nakae, M. Papilionidae of the World; Roppon-Ashi: Tokyo, Japan, 2021; p. 336. [Google Scholar]
- McNeely, J.A.; Miller, K.R.; Reid, W.V.; Mettermeier, R.A.; Werner, T.B. Conserving the World’s Biological Diversity; International Union for the Conservation of Nature and Natural Resources: Gland, Switzerland, 1990; p. 193. [Google Scholar]
- Mittermeier, R.A.; Myers, N.; Mittermeier, C.G.; Robles, G.P. Hotspots: Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions; Toppan Printing Co: Tokyo, Japan, 1999; p. 430. [Google Scholar]
- Wu, C.S.; Hsu, Y.F. Butterflies of China; The Straits Publishing & Distributing Group: Fuzhou, China, 2017; Volume 4, pp. 1442–2036. [Google Scholar]
- Li, X.; Yuan, X.Z.; Deng, H.L. Vertical idistribution and diversity of butterflies in Hengduan Mountains southwest China. Chin. J. Ecol. 2009, 28, 1847–1852. [Google Scholar] [CrossRef]
- Liu, W.P. Butterfly resources and protection of rare species in Hengduan Mountains. Sichuan J. Zool. 2005, 24, 529–531. [Google Scholar]
- Lo, Y.F.P.; Bi, Z. A preliminary report on butterfly fauna (Insecta: Lepidoptera) of Tengchong section of Gaoligongshan National Nature Reserve, China. J. Threat. Taxa 2019, 11, 14452–14470. [Google Scholar] [CrossRef]
- Checa, M.F.; Levy, E.; Rodriguez, J.; Willmott, K. Rainfall as a significant contributing factor to butterfly seasonality along a climatic gradient in the Neotropics. BioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Hill, G.M.; Kawahara, A.Y.; Daniels, J.C.; Bateman, C.C.; Scheffers, B.R. Climate change effects on animal ecology butterflies and moths as a case study. Biol. Rev. 2021, 96, 2113–2126. [Google Scholar] [CrossRef]
- Comay, O.; Yehuda, O.B.; Schwartz-Tzachor, R.; Benyamini, D.; Pe’er, I.; Ktalav, I.; Pe’er, G. Environmental controls on butterfly occurrence and species richness in Israel: The importance of temperature over rainfall. Ecol. Evol. 2021, 11, 12035–12050. [Google Scholar] [CrossRef]
- Gibbs, M.; Wiklund, C.; Van Dyck, H. Temperature, rainfall and butterfly morphology: Does life history theory match the observed pattern? Ecography 2011, 34, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.J.; Fox, R. Insect responses to global change offer signposts for biodiversity and conservation. Ecol. Entomol. 2020, 46, 699–717. [Google Scholar] [CrossRef]
- Chowdhury, S.; Braby, M.F.; Fuller, R.A.; Zalucki, M.P. Coasting along to a wider range: Niche conservatism in the recent range expansion of the Tawny Coster, Acraea terpsicore (Lepidoptera: Nymphalidae). Divers. Distrib. 2021, 27, 402–415. [Google Scholar] [CrossRef]
- Zhao, D.S.; Wu, S.H.; Yin, Y.H.; Yin, Z.Y. Vegetation distribution on Tibetan Plateau under climate change scenario. Reg. Environ. Chang. 2011, 11, 905–915. [Google Scholar] [CrossRef]
- Hu, S.J.; Xing, D.H.; Gong, Z.X.; Hu, J.M. Projecting suitability and climate vulnerability of Bhutanitis thaidina (Blanchard) (Lepidoptera: Papilionidae) with conservation implications. Sci. Rep. 2019, 9, 15384. [Google Scholar] [CrossRef] [Green Version]
- Yi, C.H.; He, F.X.; He, Q.J.; Wang, L. The biological characteristics of Bhutanitis yulongensis Chou. Chin. J. Appl. Entomol. 2011, 48, 1505–1508. [Google Scholar]
- Yi, C.H.; He, Q.J.; Wang, L.; Chen, Y.; Yi, W. Distribution, endangered reasons and protective research of Bhutanitis thaidina. Hubei Agric. Sci. 2011, 50, 2851–2854. [Google Scholar] [CrossRef]
- Minter, M.; Dasmahapatra, K.K.; Thomas, C.D.; Morecroft, M.D.; Tonhasca, A.; Schmitt, T.; Siozios, S.; Hill, J.K. Past, current, and potential future distributions of unique genetic diversity in a cold-adapted mountain butterfly. Ecol. Evol. 2020, 10, 11155–11168. [Google Scholar] [CrossRef] [PubMed]
- Oberhauser, K.; Peterson, A.T. Modeling current and future potential wintering distributions of eastern North American monarch butterflies. PNAS 2003, 100, 14063–14068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Svancara, L.K.; Abatzoglou, J.T.; Waterbury, B. Modeling current and future potential distributions of milkweeds and the Monarch Butterfly in Idaho. Front. Ecol. Environ. 2019, 7, 168. [Google Scholar] [CrossRef] [Green Version]
- Taheri, S.; Naimi, B.; Rahbek, C.; Araújo, M.B. Improvements in reports of species redistribution under climate change are required. Sci. Adv. 2021, 7, eabe1110. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014. In Synthesis Report; World Meteorological Organization: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Li, B.Y. On the boundaries of the Hengduan Mountains. Mt. Res. 1987, 5, 74–82. [Google Scholar]
- Li, B.Y. Geomorphologic regionalization of the Hengduan Mountainous region. Mt. Res. 1989, 7, 13–20. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Zhang, J.Z.; Li, X.Y.; Cao, L.Y. Hydrologic regionalization of Hengduan Mountains. Mt. Res. 1983, 13, 41–54. [Google Scholar] [CrossRef]
- Chou, I. A classification of the genus Bhutanitis a rare butterfly in the world (Lepidoptera: Papilionidae). Entomotaxonoia 1992, 14, 48–54. [Google Scholar]
- Dong, D.Z.; Kavanauph, D.; Li, H. Butterfly resources of Nujiang canyon in Yunnan. J. Southwest Agric. Univ. 2002, 24, 289–292. [Google Scholar] [CrossRef]
- Huang, H. A list of butterflies collected from Nujiang and Dulongjiang, China with descriptions of new species, and revision notes. Neue Entomol. Nachr. 2003, 55, 3–114. [Google Scholar]
- Kocman, S. Parnassius of Tibet and the Adjacent Areas; Vadim Tshhikolovets: Pardubice, Czech, 2009; p. 224. [Google Scholar]
- Li, C.L. Butterflies and their distribution in Cangshan, western Yunnan. Southwest Agric. Univ. 1991, 13, 351–368. [Google Scholar]
- Li, C.L. A new species and four new subspecies of the family Parnassiidae from Yunnan, China. J. Southwest Agric. Univ. 1994, 16, 101–105. [Google Scholar]
- Li, C.L. Yunnan Butterfly; China Forestry: Beijing, China, 1995; p. 152. [Google Scholar]
- Mao, B.Y.; Yang, Z.Z.; Bai, X.M.; Xu, J.S.; Feng, J.M. The fauna and distribution pattern of butterflies in Cangshan Erhai National Nature Reserve. Dali Univ. 2009, 8, 49–58. [Google Scholar]
- Wu, C.S.; Bai, J.W. A review of the genus Byasa Moore in China (Lepidoptera: Papilionidae). Orient. Insects 2001, 35, 67–82. [Google Scholar] [CrossRef]
- Xie, S.G.; Li, S.H. The launa vertical distribution and diversity of butterflies in Labahe nature reserve of China. J. Southwest Univ. 2007, 29, 111–117. [Google Scholar] [CrossRef]
- Peng, X.; Lei, D. Investigation on butterfly resources in Shimian county. Sichuan J. Zool. 2007, 26, 903–905. [Google Scholar]
- Zhou, G.B.; Luo, Z.J.; He, X.l. Investigation of butterflies in Sichuan province. Sichuan J. Zool. 2005, 24, 531–532. [Google Scholar]
- Racheli, T.; Cotton, A.M. Guide to the Butterflies of the Palearctic Region; Papilionidae Part I; Omnes Artes: Milano, Italy, 2009; p. 70. [Google Scholar]
- Racheli, T.; Cotton, A.M. Guide to the Butterflies of the Palearctic Region; Papilionidae. Part II; Omnes Artes: Milano, Italy, 2010; p. 86. [Google Scholar]
- Hijmans, R.J. Cross-validation of species distribution models: Removing spatialsorting bias and calibration with a null model. Ecology 2011, 93, 679–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, X.G.; Zhang, L.; Zhang, J.; Wu, T.W.; Fang, Y.J. Climate change projections over East Asia with BCC_CSM1.1 climate model under RCP scenarios. J. Meteorol. Soc. Jpn. 2013, 91, 413–429. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Wei, T.Y.; Simko, V. R Package ‘Corrplot’: Visualization of A Correlation Matrix; Version 0.92. 2021. Available online: https://github.com/taiyun/corrplot (accessed on 10 April 2022).
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Mastrandrea, M.D.; Field, C.B.; Stocker, T.F.; Edenhofer, O.; Ebi, K.L.; Frame, D.J.; Held, H.; Kriegler, E.; Mach, K.J.; Matschoss, P.R.; et al. Guidance note for lead authors of the IPCC Fifth Assessment Report on consistent treatment of uncertainties. In Proceedings of the IPCC Cross-Working Group Meeting on Consistent Treatment of Uncertainties, Jasper Ridge, CA, USA, 6 July 2010. [Google Scholar]
- Aven, T.; Renn, O. An evaluation of the treatment of risk and uncertainties in the IPCC Reports on climate change. Risk Anal. 2015, 35, 701–712. [Google Scholar] [CrossRef]
- Wang, L. The topo-climatic resource of the Hengduan Mountains and the northward shifting of rubber tree planting. Geogr. Res. 1985, 4, 71–78. [Google Scholar] [CrossRef]
- Lemoine, N.P. Climate change may alter breeding ground distributions of eastern migratory Monarchs (Danaus plexippus) via range expansion of Asclepias host plants. PLoS ONE 2015, 10, e0118614. [Google Scholar] [CrossRef] [Green Version]
- von Stetten, J.M.; Bozano, G.C. Guide to the Butterflies of the Palearctic Region; Papilionidae Part III; Omnes Artes: Milano, Italy, 2021; p. 96. [Google Scholar]
- Qiu, H.X.; Lin, Y.R. Flora of China; Science Press: Beijing, China, 1988; Volume 24. [Google Scholar]
- Lidén, M.; Fukuhara, T.; Axberg, T. Phylogeny of Corydalis, ITS and morphology. Plant Syst. Evol. 1995, 9, 183–188. [Google Scholar] [CrossRef]
- Wang, Y.W. Study on the Phylogenetic of Corydalis; Institute of Botany, The Chinese Academy of Sciences: Beijing, China, 2006. [Google Scholar]
- Wu, Z.Y.; Zhuang, X.; Su, Z.Y. The systematic evolution of Corydalis in relation to florogenesis and floristic regionalization in the world. Acta Bot. Yunnanica 1996, 18, 241–267. [Google Scholar]
- Zhuang, X. The taxonmic and evolution and distribution of Papaveraceae. Acta Bot. Yunnanica 1993, 15, 137–148. [Google Scholar]
- Zhu, J.Q.; Gu, Y.; Chen, Z.B.; Chen, J.L. The Life History of Chinese Butterflies; Chongqing University Press: Chongqing, China, 2019; p. 598. [Google Scholar]
- Li, X.W. Flora of China; Science Press: Beijng, China, 1982; Volume 31, p. 509. [Google Scholar]
- Wu, Z.Y. Flora of China; Science Press: Beijing, China, 1999; Volume 32, p. 628. [Google Scholar]
- Yu, D.J. Flora of China; Science Press: Beijing, China, 1974; Volume 36, p. 443. [Google Scholar]
- Cun, Y.Z.; Wang, X.Q. Plant recolonization in the Himalaya from the southeastern Qinghai-Tibetan Plateau: Geographical isolation contributed to high population differentiation. Mol. Phylogenetics Evol. 2010, 56, 972–982. [Google Scholar] [CrossRef]
- Hu, S.J.; Condamine, F.L.; Monastyrskii, A.L.; Cotton, A.M. A new species of the Graphium (Pazala) mandarinus group from central Vietnam (Lepidoptera: Papilionidae). Zootaxa 2019, 4554, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.J.; Cotton, A.M.; Condamine, F.L.; Duan, K.; Wang, R.J.; Hsu, Y.F.; Zhang, X.; Cao, J. Revision of Pazala Moore, 1888: The Graphium (Pazala) mandarinus (Oberthür, 1879) group, with treatments of known taxa and descriptions of new species and new subspecies (Lepidoptera: Papilionidae). Zootaxa 2018, 4441, 401–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsumura, Y.; Kimura, M.; Nakao, K.; Uchiyama, K.; Ujino-Ihara, T.; Wen, Y.F.; Tong, Z.K.; Han, W.J. Effects of the last glacial period on genetic diversity and genetic differentiation in Cryptomeria japonica in East Asia. Tree Genet. Genomes 2020, 16, 19. [Google Scholar] [CrossRef]
- Zhang, H.H.; Cotton, A.M.; Condamine, F.L.; Wang, R.J.; Hsu, Y.F.; Duan, K.; Zhang, X.; Hu, S.J. Revision of Pazala Moore, 1888: The Graphium (Pazala) alebion and G.(P.) tamerlanus groups, with notes on taxonomic and distribution confusions (Lepidoptera: Papilionidae). Zootaxa 2020, 4759, 77–97. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.J.; Gutierrez, D.; Gutierrez, J.; Monserrat, V.J. An elevational shift in butterfly species richness and composition accompanying recent climate change. Glob. Chang. Biol. 2007, 13, 1873–1887. [Google Scholar] [CrossRef]
- Levanoni, O.; Levin, N.; Pe’er, G.; Turbé, A.; Kark, S. Can we predict butterfly diversity along an elevation gradient from space? Ecography 2011, 34, 372–383. [Google Scholar] [CrossRef]
- Rahbek, C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecol. Lett. 2005, 8, 224–239. [Google Scholar] [CrossRef]
- Nogues-Bravo, D.; Araujo, M.B.; Romdal, T.; Rahbek, C. Scale effects and human impact on the elevational species richness gradients. Nature 2008, 453, 216–219. [Google Scholar] [CrossRef] [Green Version]
- Trigas, P.; Panitsa, M.; Tsiftsis, S. Elevational gradient of vascular plant species richness and endemism in Crete-the effect of post-isolation mountain uplift on a continental island system. PLoS ONE 2013, 8, e59425. [Google Scholar] [CrossRef]
- Marini, L.; Bona, E.; Kunin, W.E.; Gaston, K.J. Exploring anthropogenic and natural processes shaping fern species richness along elevational gradients. J. Biogeogr. 2011, 38, 78–88. [Google Scholar] [CrossRef]
- Sun, L.; Luo, J.; Qian, L.S.; Deng, T.; Sun, H. The relationship between elevation and seed-plant species richness in the Mt. Namjagbarwa region (Eastern Himalayas) and its underlying determinants. Glob. Ecol. Conserv. 2020, 23, e01053. [Google Scholar] [CrossRef]
- Condamine, F.L.; Sperling, F.A.; Wahlberg, N.; Rasplus, J.Y.; Kergoat, G.J. What causes latitudinal gradients in species diversity? evolutionary processes and ecological constraints on swallowtail biodiversity. Ecol. Lett. 2012, 15, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Dieker, P.; Drees, C.; Assmann, T. Two high-mountain burnet moth species (Lepidoptera, Zygaenidae) react differently to the global change drivers climate and land-use. Biol. Conserv. 2011, 144, 2810–2818. [Google Scholar] [CrossRef]
- Wu, R.D.; Zhang, S.; Yu, D.W.; Zhao, P.; Li, X.H.; Wang, L.Z.; Yu, Q.; Ma, J.; Chen, A.; Long, Y.C. Effectiveness of China’s nature reserves in representing ecological diversity. Front. Ecol. Environ. 2011, 9, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Fuller, R.A.; Dingle, H.; Chapman, J.W.; Zalucki, M.P. Migration in butterflies: A global overview. Biol. Rev. 2021, 96, 1462–1483. [Google Scholar] [CrossRef]
- Foster, P. The potential negative impacts of global climate change on tropical montane cloud forests. Earth-Sci. Rev. 2001, 55, 73–106. [Google Scholar] [CrossRef]
- Cannone, N.; Sgorbati, S.; Guglielmin, M. Unexpected impacts of climate change on alpine vegetation. Front. Ecol. Environ. 2007, 5, 360–364. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.J.; Gutierrez, D.; Gutierrez, J.; Martinez, D.; Agudo, R.; Monserrat, V.J. Changes to the elevational limits and extent of species ranges associated with climate change. Ecol. Lett. 2005, 8, 1138–1146. [Google Scholar] [CrossRef]
- Maclean, I.M.; Wilson, R.J. Recent ecological responses to climate change support predictions of high extinction risk. Proc. Natl. Acad. Sci. USA 2011, 108, 12337–12342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, J.H.; Luo, Y.Q.; Niu, B.; Tu, A.Q.; Zhao, L. Biological characteristics and habitat requirements of Parnassius imperator (Lepidoptera: Parnassidae). Acta Ecol. Sin. 2012, 32, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Liang, Q.L.; Xu, X.T.; Mao, K.S.; Wang, M.C.; Wang, K.; Xi, Z.X.; Liu, J.Q. Shifts in plant distributions in response to climate warming in a biodiversity hotspot, the Hengduan Mountains. J. Biogeogr. 2018, 45, 1334–1344. [Google Scholar] [CrossRef]
- Lu, S.F.; Zhou, S.Y.; Yin, X.J.; Zhang, C.; Li, R.L.; Chen, J.H.; Ma, D.X.; Wang, Y.; Yu, Z.X.; Chen, Y.H. Patterns of tree species richness in Southwest China. Environ. Monit. Assess. 2021, 193, 97. [Google Scholar] [CrossRef]
- Hu, R.C.; Gu, Y.Y.; Luo, M.; Lu, Z.; Wei, M.; Zhong, J. Shifts in bird ranges and conservation priorities in China under climate change. PLoS ONE 2020, 15, e0240225. [Google Scholar] [CrossRef] [PubMed]
- Lamsal, P.; Kumar, L.; Aryal, A.; Atreya, K. Future climate and habitat distribution of Himalayan Musk Deer (Moschus chrysogaster). Ecol. Inform. 2018, 44, 101–108. [Google Scholar] [CrossRef]
- Taylor, G.S.; Braby, M.F.; Moir, M.L.; Harvey, M.S.; Sands, D.P.A.; New, T.R.; Kitching, R.L.; McQuillan, P.B.; Hogendoorn, K.; Glatz, R.V.; et al. Strategic national approach for improving the conservation management of insects and allied invertebrates in Australia. Austral Entomol. 2018, 57, 124–149. [Google Scholar] [CrossRef]
- Chowdhury, S.; Alam, S.; Chowdhury, S.U.; Rokonuzzaman, M.D.; Shahriar, S.A.; Shome, A.R.; Fuller, R.A. Butterflies are weakly protected in a mega-populated country, Bangladesh. Glob. Ecol. Conserv. 2021, 26, e01484. [Google Scholar] [CrossRef]
- CBD. Aichi Biodiversity Targets. Available online: https://www.cbd.int/sp/targets/ (accessed on 13 February 2023).
- NDRC; MNR. General Plans for the Conservation and Restoration of Important Ecosystems in China (Years 2021 to 2035); NDRC: Beijing, China, 2020.
- Chowdhury, S.; Jennions, M.D.; Zalucki, M.P.; Maron, M.; Watson, J.E.M.; Fuller, R.A. Protected areas and the future of insect conservation. Trends Ecol. Evol. 2023, 38, 85–95. [Google Scholar] [CrossRef]
- Diao, Y.X.; Wang, J.J.; Yang, F.L.; Wu, W.; Zhou, J.; Wu, R.D. Identifying optimized on-the-ground priority areas for species conservation in a global biodiversity hotspot. J. Environ. Manag. 2021, 290, 112630. [Google Scholar] [CrossRef]
- Wang, J.J.; Wu, R.D.; He, D.M.; Yang, F.L.; Hu, P.J.; Lin, S.W.; Wu, W.; Diao, Y.X.; Guo, Y. Spatial relationship between climatic diversity and biodiversity conservation value. Conserv. Biol. 2018, 32, 1266–1277. [Google Scholar] [CrossRef]
- Yang, F.L.; Hu, J.M.; Wu, R.D. Combining endangered plants and animals as surrogates to identify priority conservation areas in Yunnan, China. Sci. Rep. 2016, 6, 30753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özdemirel, B.K. Investigating cross congruence between butterfly taxa and ecological community within the framework of systematic conservation planning: A case study from Lesser Caucasus Ecoregion of Turkey. Eur. J. Ecol. 2021, 7, 54–62. [Google Scholar] [CrossRef]
- Fajardo, J.; Lessmann, J.; Bonaccorso, E.; Devenish, C.; Muñoz, J. Combined use of systematic conservation planning, species distribution modelling, and connectivity analysis reveals severe conservation gaps in a megadiverse country (Peru). PLoS ONE 2014, 9, e114367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddad, N.M.; Hudgens, B.; Damiani, C.; Gross, K.; Kuefler, D.; Pollock, K. Determining optimal population monitoring for rare butterflies. Conserv. Biol. 2008, 22, 929–940. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.Z.; Xu, H.G.; Chen, M.M.; Tong, W.J.; Wang, C.B.; Cai, L. Progress in construction of China butterfly diversity observation network (China BON-Butterflies). J. Ecol. Rural. Environ. 2018, 34, 27–36. [Google Scholar] [CrossRef]
- Taron, D.; Ries, L.; Daniels, J.C. Butterfly Conservation in North America; Springer: New York City, NY, USA, 2015; p. 192. [Google Scholar]
- Chowdhury, S. Threatened species could be more vulnerable to climate change in tropical countries. Sci. Total Environ. 2023, 858, 159989. [Google Scholar] [CrossRef]
- Chowdhury, S.; Zalucki, M.P.; Hanson, J.O.; Tiatragul, S.; Green, D.; Watson, J.E.M.; Fuller, R.A. Three-quarters of insect species are insufficiently represented by protected areas. One Earth 2023, 6, 139–146. [Google Scholar] [CrossRef]
- IUCN. IUCN WCPA Other Effective Area-Based Conservation Measures Specialist Group. Available online: https://www.iucn.org/our-union/commissions/group/iucn-wcpa-other-effective-area-based-conservation-measures-specialist (accessed on 28 February 2023).
- Yang, R.M.; Hu, S.J.; Blanchard, B.D.; Nakamura, A. The local ecological knowledge of butterfly diversity is derived from utilitarian purposes in Southwest China’s biodiversity hotspot. Biodivers. Conserv. 2022, 31, 1927–1943. [Google Scholar] [CrossRef]
- Haddad, N.M. The Last Butterflies: A Scientist’s Quest to Save a Rare and Vanishing Creature; Princeton University Press: Princeton, NJ, USA, 2019; p. 250. [Google Scholar]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef] [Green Version]
- Filazzola, A.; Matter, S.F.; Roland, J. Inclusion of trophic interactions increases the vulnerability of an alpine butterfly species to climate change. Glob. Chang. Biol. 2020, 26, 2867–2877. [Google Scholar] [CrossRef] [PubMed]
- Batalden, R.V.; Oberhauser, K.; Peterson, A.T. Ecological niches in sequential generations of eastern North American Monarch butterflies (Lepidoptera: Danaidae): The ecology of migration and likely climate change implications. Environ. Entomol. 2007, 36, 1365–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparks, T.H.; Yates, T.J. The effect of spring temperature on the appearance dates of British butterflies 1883–1993. Ecography 1997, 20, 368–374. [Google Scholar] [CrossRef]
- Crozier, L. Warmer winters drive butterfly range expansion by increasing survivorship. Ecology 2004, 85, 231–241. [Google Scholar] [CrossRef]
- Pateman, R.M.; Hill, J.K.; Roy, D.B.; Fox, R.; Thomas, C.D. Temperature-dependent alterations in host use drive rapid range expansion in a butterfly. Science 2012, 336, 1028–1030. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.-T.; Yang, F.-L.; Da, W.; Li, Y.-C.; Xi, H.-M.; Cotton, A.M.; Zhang, H.-H.; Duan, K.; Xu, Z.-B.; Gong, Z.-X.; et al. Species Richness of Papilionidae Butterflies (Lepidoptera: Papilionoidea) in the Hengduan Mountains and Its Future Shifts under Climate Change. Insects 2023, 14, 259. https://doi.org/10.3390/insects14030259
Yu X-T, Yang F-L, Da W, Li Y-C, Xi H-M, Cotton AM, Zhang H-H, Duan K, Xu Z-B, Gong Z-X, et al. Species Richness of Papilionidae Butterflies (Lepidoptera: Papilionoidea) in the Hengduan Mountains and Its Future Shifts under Climate Change. Insects. 2023; 14(3):259. https://doi.org/10.3390/insects14030259
Chicago/Turabian StyleYu, Xin-Tong, Fei-Ling Yang, Wa Da, Yu-Chun Li, Hong-Mei Xi, Adam M. Cotton, Hui-Hong Zhang, Kuang Duan, Zhen-Bang Xu, Zhi-Xian Gong, and et al. 2023. "Species Richness of Papilionidae Butterflies (Lepidoptera: Papilionoidea) in the Hengduan Mountains and Its Future Shifts under Climate Change" Insects 14, no. 3: 259. https://doi.org/10.3390/insects14030259